Highly Conserved Influenza A Nucleoprotein as a Target for Broad-Spectrum Intervention: Characterization of a Monoclonal Antibody with Pan-Influenza Reactivity
Jingrui Liu, Wenming Gao, Kunkun Zhao, Zongmei Huang, Lin Liu, Jingjing Chang, Xiaoyang Cao, Wenwen Zhou, Xiaojie Zhou, Yuman Liu, Xinsheng Li, Yapeng Song

TL;DR
A new monoclonal antibody called 2D8 can detect multiple influenza A subtypes, offering a tool for universal diagnostics and vaccines.
Contribution
The antibody 2D8 targets a highly conserved NP protein epitope, enabling broad detection of influenza A subtypes.
Findings
2D8 mAb cross-reacts with H1, H3, H5, H7, and H9 influenza subtypes.
The antibody binds to a conserved NP motif (38RFYIQMCTEL47) present in 35 global virus samples.
2D8 shows no cross-reactivity with unrelated viruses and has an IgG2b/κ subclass with high titer.
Abstract
The influenza A virus poses a persistent global threat to human health and the poultry industry, as it evolves rapidly and can cross species to spread. To enhance our ability to track these viruses, scientists are seeking “universal” tools that can identify multiple different influenza virus strains at once. In this study, we developed a special monoclonal antibody named 2D8, which targets the NP protein of the influenza virus. This protein is a part of the virus that remains almost unchanged across different influenza subtypes. Our tests showed that this antibody can identify multiple influenza subtypes, including those found in birds, pigs, and humans, such as H1, H3, H5, H7, and H9. We also identified the precise antigenic epitope where this antibody binds to the virus and confirmed that this site is almost identical in 35 different virus samples from around the world. These findings…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
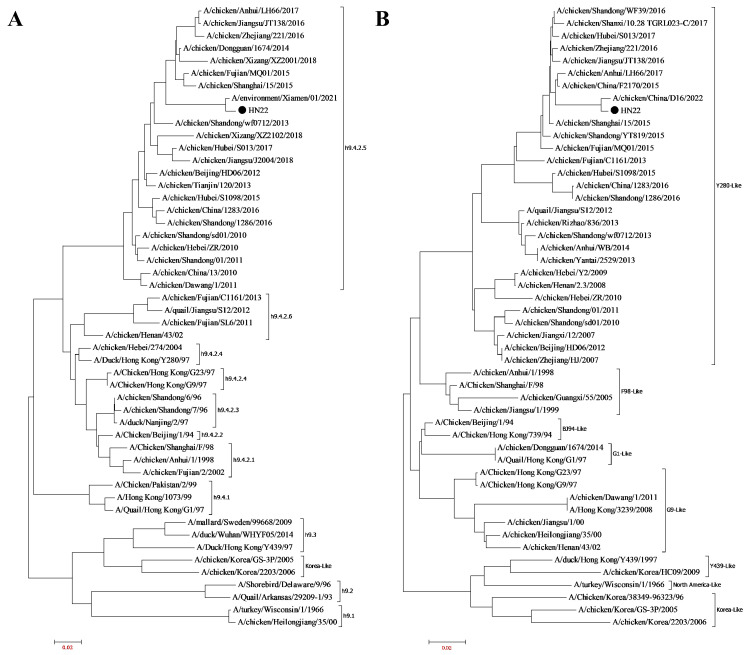 Figure 1
Figure 1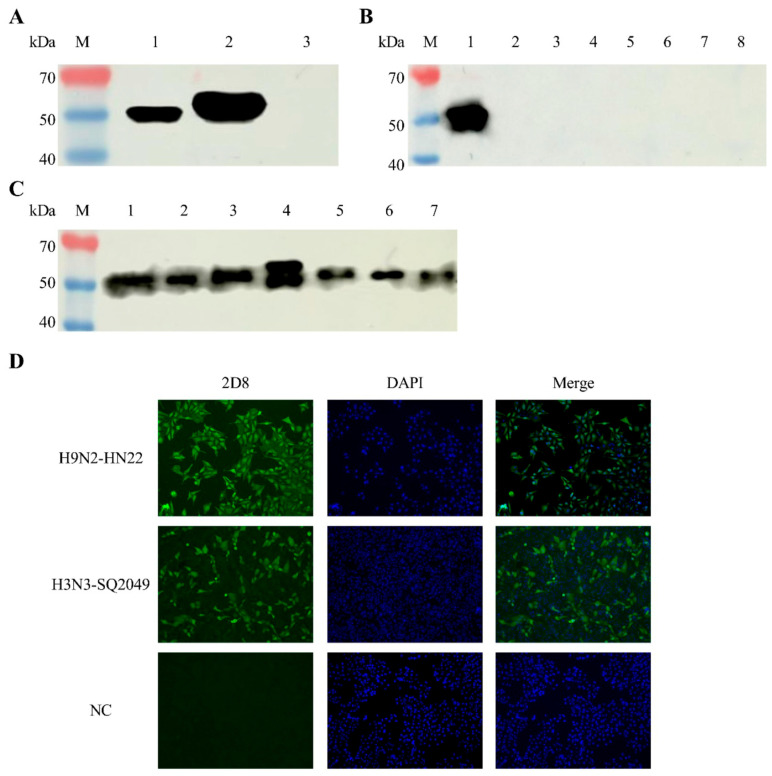 Figure 2
Figure 2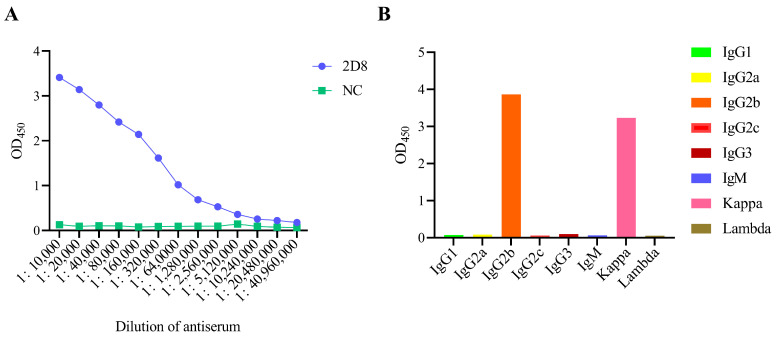 Figure 3
Figure 3 Figure 4
Figure 4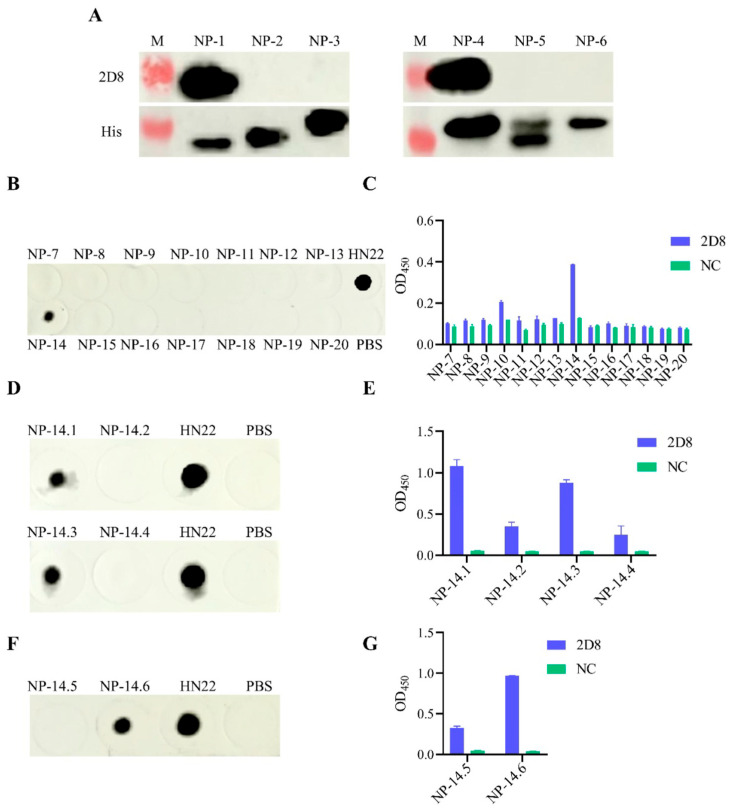 Figure 5
Figure 5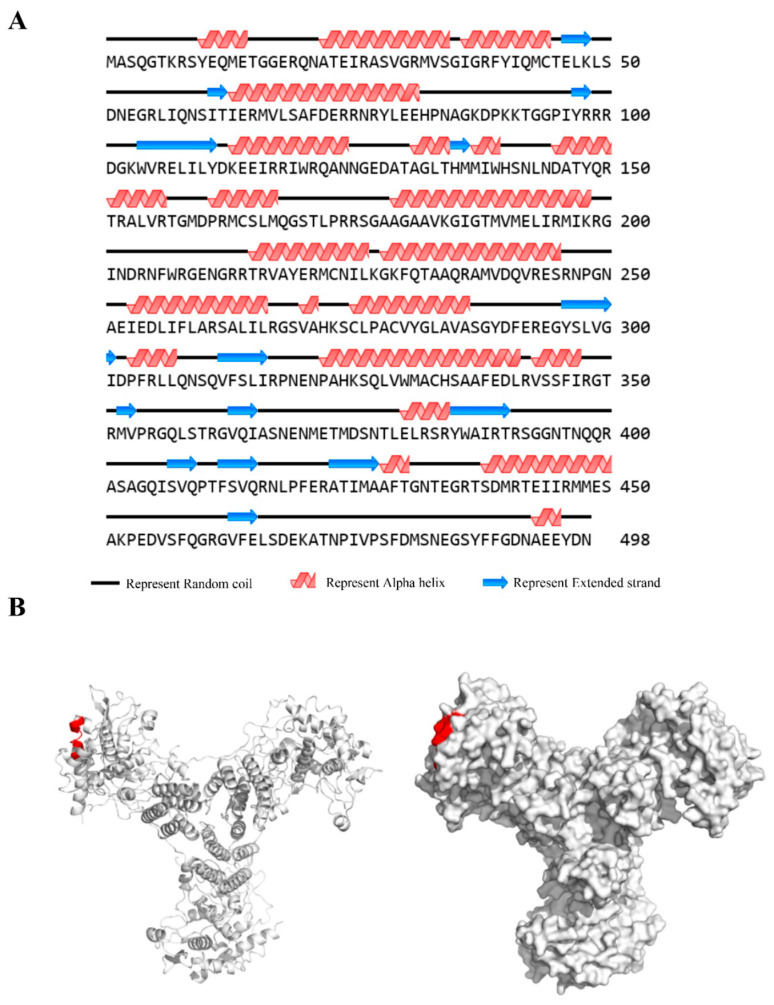 Figure 6
Figure 6 Figure 7
Figure 7- —National Natural Science Foundation of China
- —Henan Open Competition Mechanism to Select the Best Candidates Program
- —Key Scientific Research Projects of Colleges and Universities in Henan Province
- —Scientific and Technological Project of Henan Province, China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInfluenza Virus Research Studies · Respiratory viral infections research · interferon and immune responses
1. Introduction
Influenza A viruses, a major global health threat, are single-stranded, negative-sense RNA viruses belonging to the Orthomyxoviridae family [1]. These viruses are classified into subtypes based on the antigenic properties of their surface glycoproteins, hemagglutinin (HA), and neuraminidase (NA), with 16 HA and 9 NA subtypes identified across a range of hosts, including humans, birds, and mammals [2]. Human influenza A viruses, notably subtypes H1N1, H3N2, and seasonal variants, contribute to significant morbidity and mortality worldwide, exacerbating the global burden of respiratory diseases. In particular, the ongoing evolution of influenza A viruses, coupled with their ability to cross species barriers and reassort with avian strains, poses continuous threats to public health, as seen in recent years with novel influenza variants [3,4].
One of the most concerning subtypes of avian influenza virus (AIV), H9N2, has demonstrated zoonotic potential and has been associated with human infections in various regions [5,6,7,8,9,10,11]. Over time, H9N2 has undergone significant genetic diversification and recombination with other influenza subtypes, enhancing its ability to infect both poultry and humans [12,13,14,15,16,17,18,19,20,21]. Although vaccines targeting H9N2 have been used in the poultry industry, their effectiveness has waned over time due to viral mutations and antigenic drift, underscoring the urgent need for broad-spectrum diagnostic and therapeutic solutions that can provide protection against a wide range of influenza A subtypes.
The nucleoprotein (NP) of influenza A viruses is a highly conserved, internal protein essential for viral replication. It is one of the most abundant viral proteins and plays a critical role in the assembly and functioning of the viral ribonucleoprotein (vRNP) complex, which is required for the replication, transcription, and packaging of the viral genome [22,23,24,25,26]. Due to its fundamental role in the virus life cycle and its relative conservation across influenza A subtypes, the NP protein has emerged as a promising target for the development of universal diagnostics and broad-spectrum vaccines. In fact, diagnostic assays based on monoclonal antibodies (mAbs) targeting the NP protein have demonstrated the ability to recognize diverse influenza A subtypes, facilitating early detection and surveillance of both seasonal and emerging strains [27].
Studies have shown that anti-NP antibodies cross-react with a wide range of influenza A strains, including those from avian, swine, and human origins [28]. The conserved nature of the NP protein across influenza A subtypes allows for the development of diagnostic tools that can detect multiple viral strains with a single reagent, regardless of their HA or NA antigenic variation. Furthermore, NP-based vaccines have demonstrated broad protective potential against various influenza A strains, making them an attractive target for universal influenza vaccines [29,30,31,32,33].
In this study, we focused on the development of a mAb targeting the NP protein of H9N2 AIV, with the aim of expanding its applicability to human influenza viruses. By immunizing mice with the H9N2 strain, monoclonal antibodies were prepared. The broad spectrum and specificity of the monoclonal antibody were verified through various methods. The conservation of the recognized B-cell epitopes was determined through epitope mapping and homology analysis, offering insights into the potential for universal diagnostics. This research provides foundational knowledge for the development of mAb-based diagnostic assays that can accurately detect influenza A infections across species, aiding in both surveillance and early detection of emerging strains.
2. Materials and Methods
2.1. Cell Lines, Viruses, and Animals
Madin-Darby canine kidney (MDCK) cells were cultivated in Dulbecco’s modified Eagle’s medium (BioChannel, Nanjing, China), supplemented with 10% fetal bovine serum (KEL Biotech, Shanghai, China). Mouse myeloma cells (SP2/0) were cultured in hybridoma cell serum-free medium (BasalMedia, Shanghai, China) at 37 °C with 5% CO_2_. The MDCK cell line (ATCC Cat # CCL-34) and SP2/0 cell line (ATCC Cat # CRL-1581) were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). The following strains of influenza virus have been identified: avian influenza virus H9N2 subtype HN22 strain (A/chicken/China/HN22/2022) (GenBank accession number: PP030808-PP030815); avian influenza virus H9N2 subtype SQ2023 strain (A/chicken/Shangqiu/SQ2023/2023) (GenBank accession number: PP940140-PP940147); avian influenza virus H3N3 subtype SQ2049 strain (A/chicken/Henan Shangqiu/SQ2049/2023) (GenBank accession number: PP758466-PP758473); swine influenza virus H1N1 subtype ZC90 strain (A/swine/Zhucheng/90/2014) (GenBank accession number: KX264367-KX264374); the infectious bronchitis virus C2023-03 strain (CK/CH/HN/YZ202303) (GenBank accession number: PP504785); and the fowl aviadenovirus WZ strain (FAdV-WZ) (GenBank accession number: MZ508442.1). In the present study, three strains of viruses were preserved in the laboratory, namely, the fowl aviadenovirus HN1472 strain (FAdV-HN1472) (GenBank accession number: OR975470.1), the duck adenovirus GD2588 strain (DAdV-GD2588) (GenBank accession number: PV988401), and the chicken anemia virus HN2021-1412 strain (CIAV-1412) (GenBank accession number: MZ369153). The hemagglutination inhibition test antigens of the following influenza virus subtypes were procured from Harbin Guosheng Biotechnology Co., Ltd. (Harbin, China): H5 (Re-13 strain and Re-14 strain) and H7 (Re-4 strain). The hemagglutination inhibition test antigens of the following virus were procured from Beijing Zhonghai Biotech Co., Ltd. (Beijing, China): the egg drop syndrome virus 911 strain (EDSV-911) and the Newcastle disease virus La Sota strain (NDV-La Sota). Six 6–8-week-old BALB/c mice were obtained from Changchun Changsheng Biotechnology Co., Ltd. (Changchun, China). Among them, three were used as the immunization group and the other three as the control group. Specific pathogen-free (SPF) chicken embryos aged 9–11 days were purchased from Beijing Boehringer Ingelheim Vital Biotechnology Co., Ltd. (Beijing, China). All immunization procedures adhered to the institutional animal care and use guidelines (Animal Welfare and Ethics Approval No. HNND2024052225).
2.2. The Isolation of the Virus Strain
The H9N2-HN22 strain was isolated from diseased tissues of a chicken farm in Henan Province and purified by SPF chicken embryos. The purified virus was then serially diluted 10-fold with phosphate-buffered saline (PBS) and inoculated into 9–11-day-old SPF chicken embryos at a dose of 0.2 mL per embryo. The embryonic chicken eggs were cultured in an incubator set at 37 °C, and observations were conducted up to 120 h post-inoculation. The hemagglutination (HA) titers of the allantoic fluid were determined for each chicken embryo, and the 50% egg infectious dose (EID_50_) and 50% lethal dose (ELD_50_) of the virus were calculated using the Reed–Muench method. The virus was serially diluted 10-fold and inoculated into MDCK cells. Following a 72 h incubation period at 37 °C within an incubator, the cell culture medium was harvested for the purpose of determining the HA titers. The 50% tissue culture infectious dose (TCID_50_) was subsequently calculated in accordance with the Reed–Muench method.
2.3. Genome Sequencing and Analysis
The viral RNA was extracted according to the instructions for the FastPure Viral DNA/RNA Mini Kit (Vazyme, Nanjing, China) reagent kit and then reverse-transcribed into cDNA using the HiScript II Q Select RT SuperMix for qPCR (+gDNA wiper) (Vazyme, Nanjing, China) reagent kit. Whole-genome amplification was performed using the primers designed by Hoffmann [34], and the amplification results were sent to Sangon Biotech (Shanghai) Co., Ltd. (Shanghai, China) for Sanger sequencing. Phylogenetic analysis was then carried out using the MEGA 7.0 software.
2.4. Mouse Immunization
The viral precipitates were resuspended in PBS and then incubated with 0.1% formaldehyde at 37 °C and oscillated at 180 rpm for 72 h to inactivate the virus. The inactivated virus was then filtered and sterilized using a 0.22 µm filter before being stored at −80 °C.
Female BALB/c mice were injected subcutaneously with the inactivated H9N2-HN22 strain mixed with Freund’s complete adjuvant (Sigma-Aldrich, St. Louis, MO, USA) at a volume ratio of 1:1. The mice were injected with 300 μL per mouse at the first immunization and then boosted with the same dose at two-week intervals. Serum was collected, and the serum titer was assessed using an ELISA. The mouse with the highest antibody level was further injected with a total of 300 μL of antigen intraperitoneally, three days prior to cell fusion, as part of the final immunization. Subsequently, splenocytes were aseptically extracted and fused with SP2/0 cells using 50% (w/v) PEG 1500 (Roche, Basel, Switzerland). The fused cells were cultured in 96-well plates in hybridoma cell serum-free medium with HAT for 10 days. Positive hybridoma cell lines identified by ELISA were subcloned three times by limited dilution. The mAb subtypes were identified using a mouse monoclonal antibody-isotyping ELISA kit (Proteintech, Wuhan, China), according to the manufacturer’s instructions. The specificity and reactivity of the antibodies were detected using Western blot and immunofluorescence assay.
2.5. Indirect Enzyme-Linked Immunosorbent Assay (iELISA)
An indirect ELISA was used to measure antibody levels in the supernatant of hybridoma cells. Briefly, ELISA plates were coated with inactivated H9N2-HN22 virus and incubated at 37 °C for 2 h and at 4 °C overnight. After three washes with PBST buffer, the plates were blocked with 5% skimmed milk at 37 °C for 2 h.
Following three more washes with PBST, the hybridoma cell culture supernatants were added to the plates and incubated at 37 °C for 1 h. After three more washes with PBST, the plates were incubated with HRP-conjugated goat anti-mouse IgG (H + L) (Proteintech, Wuhan, China) at 37 °C for 1 h. Following another three washes, TMB substrate (Solarbio, China) was added to the wells, and the reaction was carried out for 15 min before being terminated with 2 M H_2_SO_4_. The absorbance value was then detected at OD_450_ nm using an enzyme meter. In the course of the experiment, the reliability of the experiment was improved by setting up three replicate groups.
2.6. Subtype Identification
Mouse monoclonal antibody isotyping was performed using the mouse monoclonal antibody-isotyping ELISA kit (Proteintech, Wuhan, China), according to the manufacturer’s protocol, to determine the subtypes of selected mAb.
2.7. Western Blot
The collected samples were mixed with 5× protein loading buffer and boiled for 10 min. The proteins in the samples were separated by SDS-PAGE electrophoresis at 10% and transferred to nitrocellulose membranes (NC membranes) (Cytiva, Dassel, Germany), which were blocked with 5% skimmed milk at room temperature (RT) for 2 h. The NC membranes were then incubated with monoclonal or tagged antibodies screened with positive sera for 2 h at RT or overnight at 4 °C. After three washes with TBST, the membranes were incubated with HRP-conjugated goat anti-mouse IgG (H + L) as the secondary antibody at RT for 2 h at a dilution of 1:5000. Following three additional washes with TBST, the target proteins were detected using an Ultrasensitive ECL Detection Kit (Proteintech, Wuhan, China) and a Western blot imaging system (Cytiva, Tokyo, Japan).
2.8. Indirect Immunofluorescence Assay (IFA)
To identify the specificity of the anti-H9N2 AIV mAb, an immunofluorescence assay (IFA) was performed on MDCK cells infected with H9N2 AIV and H3N3 AIV. After 48 h, the cells were fixed with 4% paraformaldehyde for 30 min and permeabilized with a solution of 0.5% Triton X-100 for 15 min. Following an overnight incubation with 5% skim milk at 4 °C, the antibodies were added and incubated at RT for 1 h. After three washes with PBS, the cells were incubated with Dy-Light 488-conjugated goat anti-mouse IgG (H + L) (Abbikne, Wuhan, China) at a dilution of 1:5000 for 1 h. After staining with a 4′,6-diamidino-2-phenylindole (DAPI) solution (Solarbio, Beijing, China) for 15 min, the cells were imaged using a fluorescence microscope (Thermo Fisher Scientific, Waltham, MA, USA).
2.9. Dot Blot
The NP protein peptides were applied to the NC membrane, with PBS acting as the negative control and the isolate acting as the positive control. The samples were covered with 5% skimmed milk at RT for 1 h. After three washes with TBST, the NC membranes were then incubated with the antibodies screened with positive sera for 1 h at RT. After three further TBST washes, the membranes were treated with a 1:5000 dilution of HRP-conjugated goat anti-mouse IgG (H + L) as the secondary antibody for 1 h. The target proteins were detected using an Ultrasensitive ECL Detection Kit and a Western blot imaging system.
2.10. Truncation and Expression of NP Genes
The H9N2-HN22 NP protein was cloned into the pET-28a (+) vector and then transformed into BL21 (DE3) for expression. To find the B-cell epitope of this NP mAb, we used truncated primers based on the NP gene sequence of the isolate. To achieve a consistent size of the truncated fragments, two distinct expression vectors, pET-28a (+) and pET-32a (+), were employed. The truncated NP protein was divided into three pieces: NP-1, NP-2, and NP-3. These fragments were copied into the pET-28a (+) vector and then transformed into BL21 (DE3) for expression. The Western blot analysis was performed using the mAb and the His-tagged antibody, as explained earlier. To further locate the antigenic epitope, NP-4, NP-5, and NP-6 were cloned into the pET-32a (+) for expression, followed by Western blot analysis. In order to work out the antigenic epitopes, different peptides were made bit and bit, and the antigenic sites of the mAb were found using Dot Blot and ELISA. The primers used for the above-mentioned NP-truncating genes are shown in Table 1.
2.11. Further Localization of the Amino Acid Sequence of the Linear B-Cell Epitope
Some positive fragments were found after the first truncation. We then made more of these fragments through further truncation. The peptides made using this method were then identified by ELISA and Dot Blot. The peptides were synthesized by GL Biochem (Shanghai) Ltd. (Shanghai, China).
2.12. H9N2-HN22 NP Protein Structure Prediction and Conservation Analysis of Antigenic Epitopes
Online websites (https://novopro.cn/tools/secondary-structure-prediction.html (accessed on 18 May 2025)) were used to predict the secondary structure of the H9N2-HN22 NP protein and analyze the characteristics of the selected epitopes. The three-dimensional structure of the H9N2-HN22 NP protein was predicted using the SWISS-MODLE websites (https://swissmodel.expasy.org/ (accessed on 18 July 2025)), and the selected antigenic epitopes were displayed on the 3D structure of the NP protein using the PyMol software (version 1.5.0.3). The full-length genes of the influenza virus NP protein from 35 different types of the virus were obtained from the GISAID database. Using the MEGA 7.0 software, the sequences of the NP proteins of the selected virus strains were compared to see how much the antigenic epitopes were conserved.
2.13. Statistical Analysis
This study used the GraphPad Prism software (version 9.0.2.161) to conduct the statistical analysis. T-tests were employed to analyze statistical significance, with * p < 0.05 indicating a significant difference, ** p < 0.01 indicating a highly significant difference, and *** p < 0.001 indicating an extremely significant difference.
3. Results
3.1. Virus Isolation and Identification
The diseased material was collected and inoculated via allantoic cyst. The allantoic fluid HA titer was 9 log2. The results of the whole-genome sequencing showed that this virus was an H9N2 subtype of avian influenza virus. This strain was named H9N2-HN22 strain (A/chicken/China/HN22/2022). The Reed–Muench method showed that the EID_50_ value was 10^−8.0^/0.1 mL, the ELD_50_ value was 10^−8.0^/0.1 mL, and the TCID_50_ value was 10^−5.1^/0.1 mL. A phylogenetic evolutionary tree for the HA gene and the NA gene was made by using the MEGA 7.0 software. Phylogenetic analysis indicated that the HA gene belonged to the h9.4.2.5 lineage, and the NA gene belonged to the Y280 lineage (Figure 1).
3.2. Generation of mAbs Against H9N2-HN22 Strain
To make mAbs against H9N2 AIVs, 6-week-old BALB/c mice were subcutaneously immunized with the inactivated H9N2-HN22 strain. Then, the SP2/0 myeloma cells were mixed with splenocytes from immunized mice using a substance called PEG 1500. Antibody-positive hybridoma cell lines were screened using an ELISA with inactivated viruses as coating antigens. By using a process called limited dilution subcloning, one stable and continuously antibody-secreting cell lines were identified, namely, 2D8. The OD_450_ value of the hybridoma supernatant measured by the established ELISA method was 2.634, and the titer was 1:80,000.
3.3. Identification of 2D8 mAb Characteristics
To understand the characteristics of 2D8 mAb, a series of tests was performed, including Western blot, IFA, subtype identification, and ascites titer determination. The results from the Western blot showed that the 2D8 mAb reacted specifically with the H9N2-HN22 NP protein, demonstrating strong specificity. Importantly, it only reacted with the influenza strain and did not cross-react with other viral pathogens. Moreover, it displayed that it can fight off many different types of flu (Figure 2A–C and Figures S1–S3).
The IFA results confirmed that the 2D8 mAb reacted strongly with cells infected with H9N2-HN22 and H3N3-SQ2049 (Figure 2D and Figure S4). The ascites titer of 2D8 mAb detected by indirect ELISA reached an impressive level of 1:20,480,000 (Figure 3A). Also, 2D8 mAb’s heavy chain is IgG2b, and its light chain type is a kappa chain (Figure 3B).
3.4. Peptide Truncation Strategies
To find out which parts of the H9N2-HN22 NP protein are recognized by the mAb, the NP protein was cut into different lengths and used as the antigen (Figure 4). At first, the H9N2-HN22 NP protein was split into three parts, each with 200 amino acids (aa), and expressed in the prokaryotic expression system. According to the results of the Western blot, it was further divided into three segments of 80 aa each (Figure 4A). After the second truncation by Western blot, the NP protein was further truncated into 14 overlapping peptides, with each peptide made up of 15 aa (Figure 4B). Based on the results of the third truncation, we were able to accurately identify the region targeted by the mAb by truncating both ends of the NP protein (Figure 4C).
3.5. Identification of Antigenic Epitopes in 2D8 mAb
The shortened proteins made from H9N2-HN22 NP protein, called NP-1, NP-2, and NP-3, were prepared, respectively, and identified by using a special antibody, 2D8 mAb. The results showed that only part of NP-1 specifically bound to 2D8 mAb, suggesting that the antigenic epitopes are between amino acids 1 and 200 (Figure 5A and Figures S5–S8). This region was later split into three parts called NP-4, NP-5, and NP-6. The results showed that the 2D8 mAb only specifically interacted with the NP-4 fragment, suggesting that the antigenic epitope of the 2D8 mAb was located at 1–80 aa (Figure 5A). Then, fragment NP-4 was split into 14 overlapping peptide segments to see how well it could bind to the mAb. The results demonstrated that one mAb specifically attached to a part of the NP-14 (Figure 5B,C and Figure S9). After this, we cut the peptide segment NP-14 at both the N-terminal and C-terminal ends by either two or four amino acids, respectively. The results showed that the mAb only reacted with the NP-14.1 peptide and the NP-14.3 peptide. These were truncated by 2 aa at the N-terminal and 2 aa at the C-terminal, respectively (Figure 5D,E, Figures S10 and S11). Finally, 1 aa was truncated from the N-terminus of the NP-14.1 peptide and from the C-terminus of the NP-14.3 peptide. The results showed that this monoclonal antibody only reacted with the 1-aa truncated C-terminal region of the NP-14.3 peptide, namely, the NP-14.6 peptide (Figure 5F,G and Figure S12). Finally, it was concluded that the epitope targeted by the 2D8 mAb is ^38^RFYIQMCTEL^47^.
3.6. Spatial Structure of Epitope
The website https://novopro.cn/tools/secondary-structure-prediction.html (accessed on 18 May 2025) was used to predict the secondary structure of the NP protein. The results revealed that the epitope ^38^RFYIQMCTEL^47^ formed an α-helix, a random coil structure, and a structurally stable β-folding (Figure 6A). The linear B-cell epitope that was found was shown on the 3D structure of the NP protein that had been made using the PyMol software (version 1.5.0.3). It was found that a part of the NP protein structure was exposed on the surface of the NP protein (Figure 6B). This part of the NP protein is called the antigenic epitope ^38^RFYIQMCTEL^47^ (red portion).
3.7. Conservative Analysis of Identified Epitope
A comparison of the NP protein sequences of 35 different types of the influenza A virus, from both inside and outside of China, using the MEGA 7.0 software, showed that the NP-^38^RFYIQMCTEL^47^ sequence was highly conserved among these 35 strains. It is interesting to note that only three strains showed changes to the amino acids. This conserved clade encompasses the overwhelming majority of subtypes within Group 1 and Group 2. These influenza viral strains, encompassing H1 to H16, exhibit stability, irrespective of their host origins and isolation time points (Figure 7).
4. Discussion
Avian influenza viruses (AIVs) are characterized by high variability, rapid transmission, and elevated mortality rates, seriously threatening the development of the poultry industry and human life safety. The surface proteins of AIV, that is, HA, NA, and M2, have long been the primary targets for the development of antiviral strategies [35,36]. Nevertheless, recent investigations have demonstrated that novel vaccines targeting viral internal proteins exhibit more favorable antiviral efficacy [37,38]. NP is the core component of the vRNP complex, indispensable for the entire AIV life cycle [23]. Accumulating evidence indicates that anti-NP monoclonal antibodies mediate antiviral activity by inhibiting vRNP nuclear import as well as viral transcription and replication within the nucleus [39,40]. The high conservation and abundant expression of NP during viral infection render it an optimal antigenic target for the diagnosis of multiple influenza virus subtypes [41,42]. In the present study, we screened a highly broad-spectrum anti-NP monoclonal antibody, 2D8, whose recognized epitope is highly conserved across most AIV subtypes.
In this study, we successfully identified a stable monoclonal antibody (2D8 mAb) targeting the NP protein of the H9N2 subtype AIV through the immunopurification of the HN22 strain. The reactivity of this 2D8 mAb with the H9N2-HN22 strain was confirmed using various techniques, including ELISA, Western blot, and IFA. The results revealed that 2D8 mAb specifically recognizes the H9N2-HN22 strain, and, importantly, it demonstrated strong reactivity against several other influenza A subtypes, including H1, H3, H5, H7, and H9, without cross-reacting with other common poultry viruses. This broad-spectrum reactivity is consistent with the known conservation of NP across different influenza subtypes, making this antibody a valuable tool for diagnostic and surveillance applications. This cross-reactivity is especially significant as it contrasts with other NP-targeting monoclonal antibodies (e.g., 3A4 and 3G2) that are limited to human influenza viruses or specific subtypes [27,43].
Further analysis identified the specific antigenic site of 2D8 mAb as the peptide ^38^RFYIQMCTEL^47^ within the NP protein. Conservation analysis of this epitope across 35 representative influenza A strains showed remarkable stability, surpassing that of many HA and NA epitopes. This high level of conservation makes it a promising candidate for inclusion in universal influenza vaccines, particularly those aiming to provide cross-protection against diverse influenza A subtypes.
NP is a critical target for cytotoxic T lymphocytes (CTLs), which play a crucial role in inducing cellular immunity and cross-protective responses across different viral lineages [25,44,45]. Several studies have demonstrated that NP-based vaccines, such as those using adenovirus vectors or the AIP-C5 vaccine, can offer protection against influenza viruses by stimulating robust T-cell responses [30]. The OVX836 vaccine, which uses NP, has been shown to induce NP-specific lung CD8+ tissue-resident memory (TRM) cells, providing long-term protection against influenza [46]. The epitope identified in this study (^38^RFYIQMCTEL^47^) can be presented by MHC-I molecules on antigen-presenting cells (APCs), triggering a T-cell receptor (TCR)-mediated response and subsequently activating CD8+ T lymphocytes [47,48,49,50,51]. While immunization with peptides containing this sequence has shown partial protection (25%), it highlights the potential for developing a minimal epitope-based vaccine, especially if combined with multiple epitopes or fused with carrier proteins and potent adjuvants to enhance immune responses. This epitope, although a B-cell epitope, also acts as a T-cell epitope, indicating its potential to induce both humoral and cellular immunity.
Nevertheless, this study still has certain limitations. The broad-spectrum reactivity validation of the 2D8 monoclonal antibody (mAb) only encompassed a subset of host-derived strains of major influenza A virus subtypes (H1, H3, H5, H7, and H9), with insufficient coverage of influenza A viruses from diverse hosts (e.g., humans, wild birds, and non-human primates) and emerging recombinant strains. This deficiency may restrict the translational applicability of the 2D8 mAb in clinical diagnosis and therapeutic interventions. However, our current findings have confirmed that the 2D8 monoclonal antibody possesses notable broad-spectrum activity, and the epitope it recognizes is highly conserved among most AIV subtypes. Therefore, based on the current research progress, subsequent experiments will focus on establishing 2D8-based diagnostic assays to systematically evaluate its diagnostic performance against commercially available diagnostic platforms.
5. Conclusions
This study successfully screened a monoclonal antibody (2D8 mAb) targeting the NP protein of the H9N2 subtype AIV and identified its conserved B-cell epitope. The mAb exhibits broad-spectrum reactivity with multiple influenza A subtypes, underscoring its potential as a universal diagnostic tool for influenza A virus detection. More importantly, the identified epitope has been shown to be both a B-cell and a T-cell epitope, suggesting its potential for inclusion in universal influenza vaccines. These findings represent a significant step toward overcoming the limitations of current strain-specific vaccines and developing broadly protective, next-generation influenza vaccines, which could play a pivotal role in pandemic preparedness and global influenza surveillance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Paules C. Subbarao K. Influenza Lancet 201739069770810.1016/S 0140-6736(17)30129-028302313 · doi ↗ · pubmed ↗
- 2Zhuang Q. Wang S. Liu S. Hou G. Li J. Jiang W. Wang K. Peng C. Liu D. Guo A. Diversity and distribution of type A influenza viruses: An updated panorama analysis based on protein sequences Virol. J.2019168510.1186/s 12985-019-1188-731242907 PMC 6595669 · doi ↗ · pubmed ↗
- 3Garg S. Reinhart K. Couture A. Kniss K. Davis C.T. Kirby M.K. Murray E.L. Zhu S. Kraushaar V. Wadford D.A. Highly Pathogenic Avian Influenza A(H 5N 1) Virus Infections in Humans N. Engl. J. Med.202539284385410.1056/NEJ Moa 241461039740051 · doi ↗ · pubmed ↗
- 4Ryt-Hansen P. George S. Hjulsager C.K. Trebbien R. Krog J.S. Ciucani M.M. Langerhuus S.N. De Beauchamp J. Crumpton J.C. Hibler T. Rapid surge of reassortant A(H 1N 1) influenza viruses in Danish swine and their zoonotic potential Emerg. Microbes Infect.202514246668610.1080/22221751.2025.246668639945729 PMC 11849018 · doi ↗ · pubmed ↗
- 5Song W. Qin K. Human-infecting influenza A (H 9N 2) virus: A forgotten potential pandemic strain?Zoonoses Public Health 20206720321210.1111/zph.1268531930694 · doi ↗ · pubmed ↗
- 6Tan M. Zeng X. Xie Y. Li X. Liu J. Yang J. Yang L. Wang D. Reported human infections of H 9N 2 avian influenza virus in China in 2021 Front. Public Health 202311125596910.3389/fpubh.2023.125596938155898 PMC 10753182 · doi ↗ · pubmed ↗
- 7Duong M.H. Phan T.N.U. Nguyen T.H. Ho N.H.N. Nguyen T.N. Nguyen V.T. Cao M.T. Luong C.Q. Nguyen V.T. Nguyen V.T. Human Infection with Avian Influenza A(H 9N 2) Virus, Vietnam, April 2024 Emerg. Infect. Dis.20253138839210.3201/eid 3102.24114639983677 PMC 11845148 · doi ↗ · pubmed ↗
- 8Potdar V. Hinge D. Satav A. Simões E.A.F. Yadav P.D. Chadha M.S. Laboratory-Confirmed Avian Influenza A(H 9N 2) Virus Infection, India, 2019 Emerg. Infect. Dis.2019252328233010.3201/eid 2512.19063631742537 PMC 6874269 · doi ↗ · pubmed ↗