Goose Astrovirus Type 2 Causes Intestinal Injury and Disrupts Homeostasis in Goslings
Xinyang Li, Wenhao Yang, Ming Zhu, Haoran Xu, Junjie Yang, Zewen Yi, Yingjun Lv

TL;DR
This study shows how Goose Astrovirus Type 2 harms the intestines of young goslings, causing damage and inflammation, and reveals how the birds respond to the infection.
Contribution
The study reveals how GAstV-2 affects intestinal structure and homeostasis in goslings, identifying new mechanisms of pathogenesis.
Findings
GAstV-2 infects the duodenum, causing crypt necrosis and reduced villus height.
The virus disrupts stem cell populations and tight junctions, impairing intestinal barrier function.
Infection triggers an inflammatory response and compensatory increases in Paneth cells and Bmi1+ stem cells.
Abstract
Goose astrovirus type 2 (GAstV-2) was first identified in Eastern China in 2018. This virus causes mortality rates of up to 50% in goslings aged 3 to 20 days, resulting in significant economic losses to the goose industry. Infected goslings primarily exhibit urate deposition on the surfaces of visceral organs and in joint cavities, pale and swollen kidneys, and lymphocyte depletion in the spleen. However, as a fecal-oral transmitted pathogen, the interactions between GAstV-2 and digestive system organs, as well as the associated pathological damage, remain poorly understood. In this study, we found that following GAstV-2 infection in goslings, the virus was detectable in all segments of the intestine but not in the esophagus, glandular stomach, or muscular stomach. The highest viral load was observed in the duodenum, where it induced crypt necrosis, shortened villus height, a reduced…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
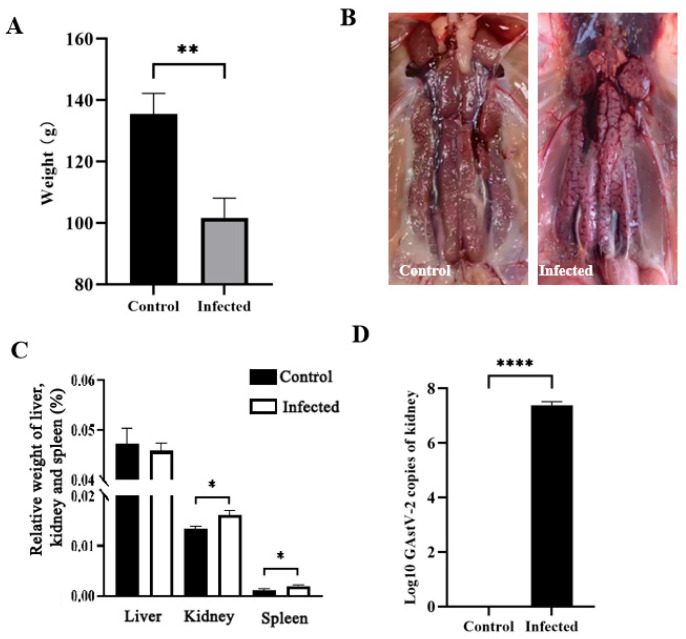 Figure 1
Figure 1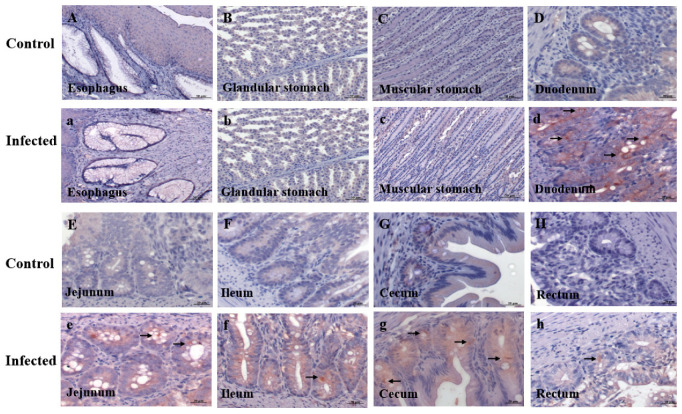 Figure 2
Figure 2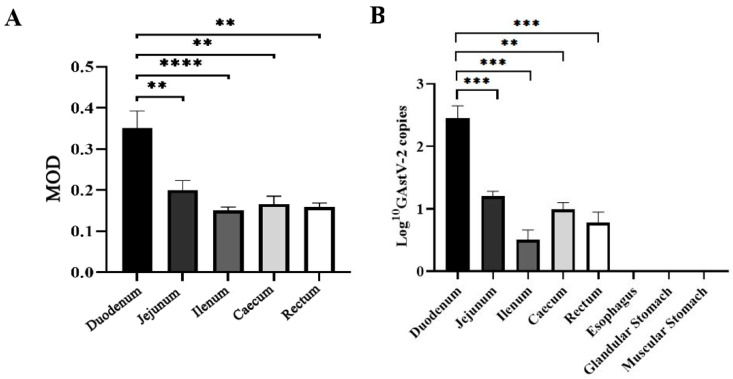 Figure 3
Figure 3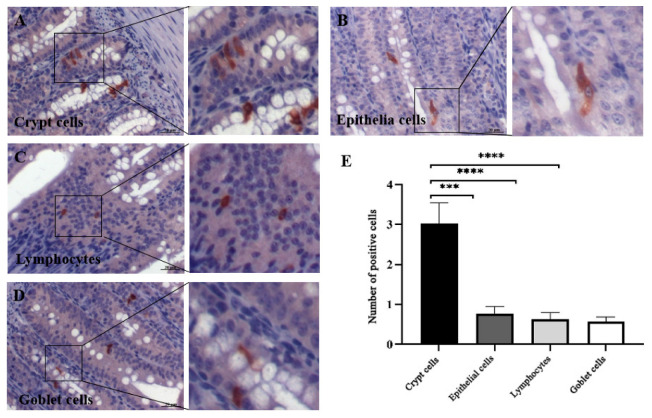 Figure 4
Figure 4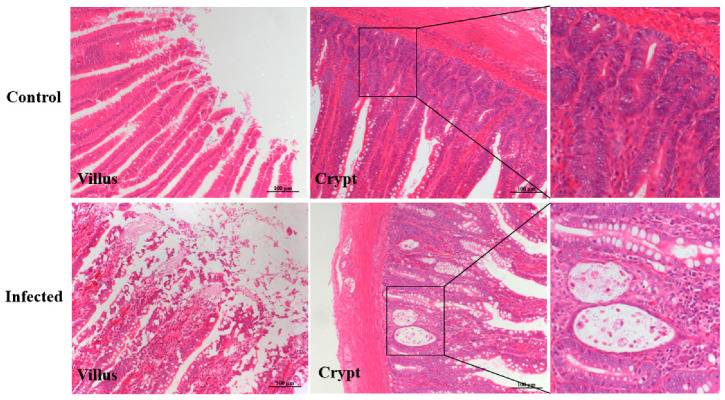 Figure 5
Figure 5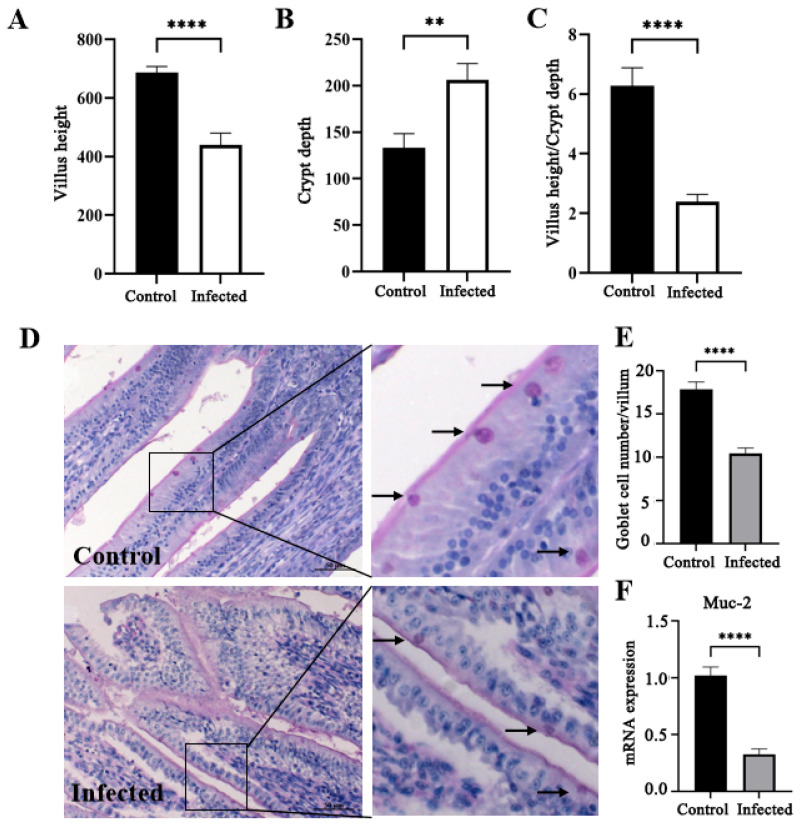 Figure 6
Figure 6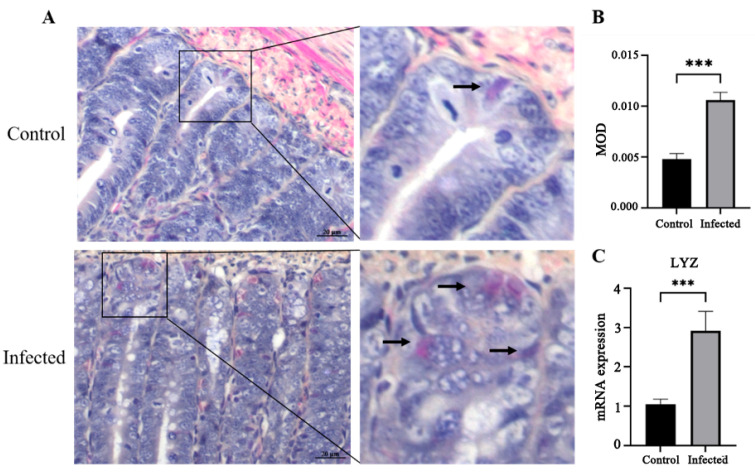 Figure 7
Figure 7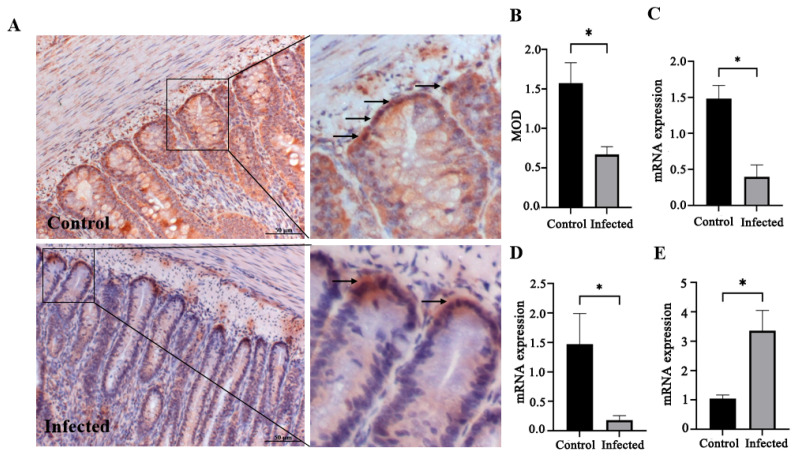 Figure 8
Figure 8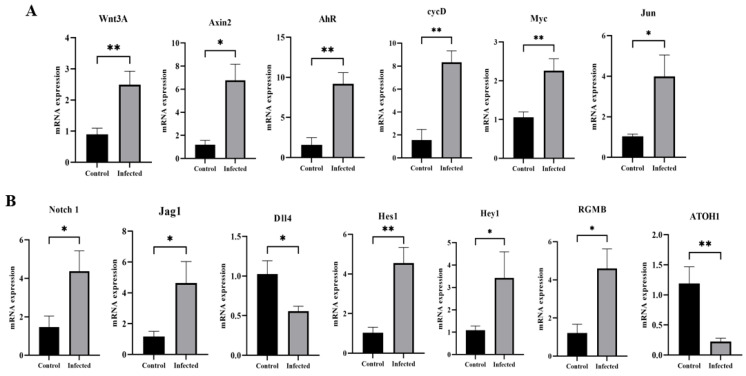 Figure 9
Figure 9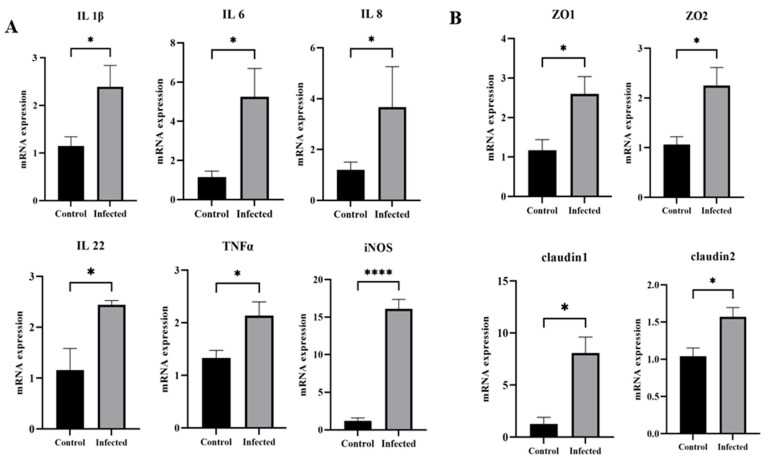 Figure 10
Figure 10- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsViral gastroenteritis research and epidemiology · Virus-based gene therapy research · Animal Virus Infections Studies
1. Introduction
Goose astrovirus (GAstV) is a non-enveloped, positive-sense, single-stranded RNA virus with a diameter of about 28–30 nm. Its genome, approximately 7.2 kb in length, consists of a 5′ untranslated region (UTR), three open reading frames (ORF1a, ORF1b, and ORF2), a 3′ UTR, and a poly(A) tail. ORF1a and ORF2b encode nonstructural proteins involved in RNA transcription and replication, such as RNA-dependent RNA polymerase (RdRP) and nuclear localization signal (NLS), while ORF2 encodes capsid proteins associated with host immune responses [1,2]. Based on amino acid sequence differences in ORF2, GAstV is classified into two genotypes, GAstV-1 and GAstV-2, which share only 55.6–57.9% genome-wide similarity and 40.4–41.6% similarity in ORF2 [3]. GAstV-2 has been identified as the primary causative agent of gout outbreaks in goslings, although GAstV-1 may also contribute, either alone or in co-infection with GAstV-2 [4,5,6]. First reported in 2018, GAstV-2 predominantly infects goslings aged 3–20 days, causing mortality rates of 5–50% in field cases and up to 93.1% in experimental infections [7,8,9,10]. Currently, there are no commercial vaccines or antiviral drugs available, leading to significant economic losses in goose production.
Gross lesions in affected goslings typically include swollen kidneys and urate deposition in the viscera and joints. Histopathological findings include renal tubular epithelial cell necrosis, hepatocyte degeneration, splenic lymphocyte depletion, and inflammatory cell infiltration [11,12]. Although GAstV-2 is transmitted via the fecal-oral route, most studies have focused on kidney pathology, with limited attention to its effects on the digestive system. The virus must traverse the intestinal barrier before reaching systemic circulation, yet the primary site of invasion, target cell types, and pathological changes in the gastrointestinal tract remain poorly characterized. Limited reports indicate GAstV-2 can be detected in the glandular stomach and duodenum and may alter the gut microbiota, increasing proinflammatory bacteria and reducing beneficial short-chain fatty acid producers [13,14]. These observations suggest that GAstV-2 may cause metabolic disturbances and intestinal injury, similar to astroviruses in other species such as human astrovirus, turkey astrovirus, and chicken astrovirus, which can damage intestinal epithelial cells, increase epithelial permeability, and impair growth [15,16,17].
Considering these knowledge gaps, the present study aimed to determine the distribution of GAstV-2 in the digestive tract of goslings, identify its target cell types, and characterize associated pathological and functional changes in the duodenum, with particular focus on intestinal stem cells, goblet cells, Paneth cells, inflammatory responses, tight junction integrity, and the involvement of Notch and Wnt signaling pathways.
2. Materials and Methods
2.1. Virus
The strain GAstV-2 (JSHA strain, GenBank: MK125058) used in this study was isolated and maintained in our laboratory. The virus titer, determined through titration on goose kidney epithelial cells using the Reed and Muench method, was 1 × 10^4.69^ TCID_50_/mL.
2.2. Animal Experiments
1-day-old Yangzhou white goose goslings, supplied by Nanjing Sanhua Goose Company Ltd. (Nanjing, China), were used in this study. Prior to the experiment, five goslings were euthanized via intravenous administration of pentobarbital sodium for baseline health assessment. Necropsy revealed no gross pathological lesions in all examined organs, and GAstV-2 RNA was not detected by RT-PCR in the kidney, spleen, or liver tissues. Subsequently, twenty 1-day-old goslings were randomly allocated into two groups: an infected group and a negative control group (n = 10 per group). All subjects were maintained in isolated negative-pressure housing under standardized conditions. They were supplied with sterilized water and a nutritionally identical, antibiotic-free diet ad libitum. The infected group received an oral challenge comprising 0.8 mL of GAstV-2 viral suspension (0.8 × 10^4.69^ TCID_50_/goose), whereas control animals were administered an equivalent volume of PBS via the identical route. Clinical signs were recorded daily after infection. Any mortalities were subjected to immediate necropsy, and gross lesions were documented. At 7 days post-infection (dpi), all surviving goslings were euthanized by intravenous injection of pentobarbital sodium following body weight measurement. The kidneys, livers, and spleens were weighed to determine relative organ weights (organ-to-body weight ratio). A portion of the kidneys, esophaguses, glandular stomachs, muscular stomachs, duodenums, jejunums, ileums, ceca, and rectums was fixed in 4% paraformaldehyde for histopathological evaluation, while the remainder was preserved at −80 °C for further analysis. Each gosling was considered to be a replicate in this study.
2.3. Histopathological Analysis
Histopathological examination was performed as previously described [11]. Briefly, fixed tissue samples were dehydrated through a graded alcohol series, cleared in xylene, and embedded in paraffin. Serial sections (4 μm thick) were then prepared and stained for examination under a light microscope (Carl Zeiss, Munich, Germany).
2.4. Periodic Acid-Schiff (PAS) and Phloxine B Paneth Cell Staining
Duodenal sections were stained with PAS and Phloxine B for Paneth cell staining to detect goblet and Paneth cells using commercial kits (Beyotime, Shanghai, China). Twenty-five random, non-overlapping high-powered fields of duodenum tissue at 400-fold magnification were chosen to count the number of PAS-positive cells. Paneth cells exhibited pink staining, and the positive signals were semi-quantitatively analyzed using ImageJ software (version 1.54g).
2.5. Virus Determination
Viral RNA was extracted from kidney and intestine samples using the RNeasy Isolation Kit (Vazyme, Nanjing, China). Subsequently, RNA was reverse-transcribed into cDNA with the HiScript Q RT SuperMix Kit (Vazyme, Nanjing, China) in accordance with the manufacturer’s instructions. Viral loads were then quantified via a SYBR Green I-based real-time PCR following a previously established protocol from our laboratory [18].
2.6. Quantitative Real-Time PCR (qRT-PCR) Analysis
Total RNA was extracted from duodenal tissue samples using RNA-easy Isolation Reagent. According to the manufacturer’s protocol, the extracted RNA was reverse-transcribed into cDNA using the HiScript Q RT SuperMix Kit. Quantitative real-time PCR (qPCR) was conducted on an AB7300 thermal cycler (Life Technologies, Carlsbad, CA, USA). Specific primers for the following target genes were designed with Primer 5.0 software: Lyz, Muc2, Lgr5, Bmi1, OLFM4, Wnt3A, Axin2, AhR, cycD, Myc, Jun, Notch1, Hes1, Dll4, Jag1, Hey1, NRARP, ATOH1, ZO-1, ZO-2, Claudin-1, Claudin-2, IL-1β, IL-6, IL-8, IL-22, TNF-α, and iNOS. Gene expression levels were quantified using the 2^−ΔΔCT^ method and normalized to GAPDH as the endogenous control. All reactions were performed in triplicate. Primer sequences are provided in Supplementary Table S1.
2.7. Immunohistochemical (IHC) Examination
IHC staining was performed following a previously described protocol with slight modifications [19]. Briefly, tissue sections were deparaffinized, rehydrated, and treated with 3% H_2_O_2_ for 10 min to inactivate endogenous peroxidase. Antigen retrieval was performed by heating the sections in citrate buffer at 100 °C for 8 min. Non-specific binding was blocked with 5% bovine serum albumin for 30 min. Sections were then incubated overnight at 4 °C with rabbit anti-GAstV-2 Capsid protein polyclonal antibody and Lgr5 Rabbit polyclonal antibody (Beyotime, Shanghai, China) as the primary antibody. After washing with PBS, sections were incubated with secondary antibodies for 1 h at 37 °C. Staining was visualized using 3,3′-diaminobenzidine (DAB) substrate, followed by counterstaining with hematoxylin for 8 min. Stained sections were examined under a light microscope, and positive signals were semi-quantitatively analyzed using ImageJ software (version 1.54g). Briefly, for each tissue section, 25 non-overlapping fields of view containing positive signals were randomly selected at 400× magnification. Within each field, the area of positive signals (brown deposition) was measured, and the percentage of positive area relative to the total field area was calculated. The values obtained from the 25 fields per sample were averaged to determine the mean optical density (MOD), which served as a quantitative indicator of target protein expression.
2.8. Statistical Analysis
The differences between the control group and experimental group were analyzed by Student’s t-test using SPSS software 16.0. Data are presented as mean ± standard deviation (SD). Statistical significance was defined as p < 0.05.
3. Results
3.1. Clinical Changes and Confirmation of GAstV-2 Infection
The goslings in the infected group exhibited depression, reduced feed intake, and mild diarrhea, with a mortality rate of 10%. No clinical signs were observed in the control group. At 7 days post-infection (dpi), body weight in the infected group was significantly lower than that in the control group (p < 0.05; Figure 1A). Gross examination revealed swollen kidneys and urate deposition in infected goslings, whereas no lesions were present in controls (Figure 1B). Relative kidney and spleen weights were significantly higher in infected birds than in controls (p < 0.05; Figure 1C). Viral loads in the kidneys of infected goslings reached 10^7^ copies/μL, whereas no viral RNA was detected in controls (Figure 1D), confirming successful GAstV-2 infection.
3.2. Distribution and Localization of GAstV-2 in Digestive Tissues
Immunohistochemistry (IHC) detected GAstV-2 antigens (brown deposition) in the duodenum, jejunum, ileum, cecum, and rectum of infected goslings, with no signals in the esophagus, glandular stomach, or muscular stomach. No positive signals were observed in any digestive tissue of controls (Figure 2). Quantitative analysis showed the strongest IHC signal intensity in the duodenum among all intestinal segments (p < 0.01) (Figure 3A). Consistent with these findings, qRT-PCR revealed that the viral load was highest in the duodenum compared to all other intestinal segments (p < 0.01), while no detectable virus was found in the esophagus, glandular stomach, or muscular stomach (Figure 3B). Within the duodenum, viral antigens were localized to crypt cells, intestinal epithelial cells, lamina propria lymphocytes, and goblet cells, with crypt cells harboring the greatest viral load (p < 0.01) (Figure 4).
3.3. Histopathological Changes in the Duodenum
Hematoxylin and eosin staining revealed crypt cell necrosis characterized by karyopyknosis and karyorrhexis, along with mild villus shedding in infected goslings, whereas no histological abnormalities were observed in controls (Figure 5).
3.4. Effect of GAstV-2 on Villus Height, Crypt Depth, Goblet Cells, and Mucus Production
The infected goslings exhibited significantly reduced villus height, increased crypt depth, and a decreased villus height-to-crypt depth ratio compared with the controls (p < 0.05; Figure 6A–C). Goblet cell counts and Muc-2 mRNA expression were markedly lower in the infected birds (p < 0.05; Figure 6D–F).
3.5. Effect of GAstV-2 on Paneth Cells and Intestinal Stem Cells
Phloxine B staining and quantitative analysis showed a significant increase in Paneth cell abundance in infected goslings compared with controls (p < 0.05; Figure 7A,B). Similarly, Lyz mRNA expression was elevated (p < 0.05; Figure 7C). IHC revealed that Lgr5+ ISCs were significantly reduced in number in infected goslings (p < 0.05; Figure 8A,B), accompanied by decreased mRNA expression of Lgr5 and OLFM4 (p < 0.05; Figure 8C,D). Conversely, Bmi1 mRNA expression was significantly increased in infected goslings (p < 0.05; Figure 8E).
3.6. Effect of GAstV-2 on Notch and Wnt Signaling Pathways
qRT-PCR analysis indicated that GAstV-2 infection significantly upregulated the mRNA expression of Wnt pathway genes Wnt3A, Axin2, cycD, AhR, Myc, and Jun (p < 0.05; Figure 9A). Notch pathway-related genes, including Notch1, Jag1, Hes1, Hey1, and NRARP, were also significantly upregulated in infected goslings, while Dll4 and ATOH1 were downregulated (p < 0.05; Figure 9B).
3.7. Effect of GAstV-2 on Inflammatory Cytokines and Tight Junction-Associated Genes
Infected goslings displayed significant increases in the mRNA expression of proinflammatory cytokines IL-1β, IL-6, IL-8, IL-22, TNF-α, and iNOS (p < 0.05; Figure 10A). Expression of tight junction-related genes ZO-1, ZO-2, Claudin-1, and Claudin-2 was also significantly elevated compared with controls (p < 0.05; Figure 10B), indicating that GAstV2 infection caused inflammation and enhanced tight junction expression in the duodenum.
4. Discussion
GAstV-2 is transmitted via the fecal–oral route, yet its localization and pathogenic effects within the digestive system are not well defined. In this study, oral inoculation of goslings with GAstV-2 resulted in reduced body weight, pale and swollen kidneys, and urate deposition, confirming the successful establishment of a gout model in goslings. Viral antigens were detected in the duodenum, jejunum, ileum, and cecum, but not in the esophagus, glandular stomach, or muscular stomach. These results indicate a clear intestinal tropism, suggesting the intestine may be the primary site of invasion following oral exposure. The duodenum harbored the highest viral load, which may be attributable to its C-shaped structure that favors viral retention or to its strong digestive and absorptive capacity. In contrast, Wei et al. reported GAstV-2 localization in the glandular stomach, a difference that may reflect variation in viral strain or experimental conditions [13]. It is also possible that after a single oral challenge, GAstV-2 transits rapidly through the glandular stomach to the intestine, limiting gastric epithelial contact. Additional studies are needed to determine whether the glandular stomach can serve as a site of GAstV-2 replication.
Our experiments further revealed that GAstV-2 localizes to intestinal crypt cells, goblet cells, intestinal epithelial cells, and lymphocytes, with the highest viral content detected in crypt cells. This distribution is consistent with infection patterns reported for astroviruses in other species: HAstV can infect intestinal epithelial cells [15], murine astrovirus localizes to goblet cells [20], and CAstV targets crypt epithelial cells [17]. Our previous work also detected GAstV-2 antigens in the cytoplasm of peripheral blood lymphocytes [21]. Furthermore, CAstV has been reported to replicate within crypts [17]. To date, there have been no reports describing GAstV-2 invasion of intestinal target cells, and it remains to be determined whether GAstV-2 can replicate within crypts.
Crypt cells play a vital role in maintaining intestinal barrier function. Located at the base of the crypt are ISCs and Paneth cells. ISCs are essential for sustaining intestinal epithelial homeostasis through continuous self-renewal and support of the rapid epithelial turnover. ISCs can be classified into two distinct populations: active ISCs, which are marked by the expression of Lgr5, and quiescent ISCs, identified by the expression of Bmi1. Lgr5+ ISCs divide regularly under steady-state conditions and are highly sensitive to injury or stress, whereas Bmi1+ ISCs contribute little to epithelial renewal during homeostasis but are activated to mediate regeneration following damage [22]. Paneth cells perform multiple functions, including antibacterial defense through the secretion of antimicrobial proteins such as lysozyme, regulation of intestinal homeostasis, and support of ISC maintenance. In this study, GAstV-2 infection resulted in a reduction in Lgr5+ ISCs, accompanied by increases in both Bmi1+ ISCs and Paneth cells. These findings suggest that GAstV-2 damages Lgr5+ ISCs, and that goslings respond by enhancing resistance to infection and promoting epithelial regeneration through increased numbers of Paneth cells and Bmi1+ ISCs.
ISCs generate transit-amplifying (TA) progenitors, which undergo terminal differentiation into specialized intestinal cell types such as enterocytes, goblet cells, and Paneth cells under the regulation of the Notch signaling pathway. Inhibition of Notch signaling has been shown to promote the development of secretory progenitor cells by upregulating the atonal basic helix-loop-helix (bHLH) transcription factor 1 (ATOH1) [23,24]. In this study, GAstV-2 infection activated the Notch signaling pathway while significantly reducing ATOH1 mRNA expression, indicating that GAstV-2 suppresses ISC differentiation into goblet cells. This finding is supported by the observed histopathological decrease in goblet cell numbers and the reduced expression of the Muc-2 gene. Wnt signaling is another key regulatory pathway for ISCs and plays a decisive role in determining their fate during epithelial repair following injury [25]. Here, GAstV-2 infection also activated the Wnt signaling pathway, which may contribute to the regeneration of damaged epithelial cells.
Tight junctions play a critical role in host defense as part of the innate immune barrier. In this study, GAstV-2 infection upregulated the expression of ZO and Claudin genes, suggesting that tight junctions may help limit viral invasion during infection. In contrast, HAstV-1 and TAstV-2 have been shown to disrupt tight junction complexes, increasing intestinal permeability and leading to diarrhea [26,27]. These findings suggest that different astrovirus species may exert distinct regulatory effects on intestinal tight junctions, potentially reflecting host-specific or virus-specific adaptations. Indeed, phylogenetic analysis of complete genomic sequences placed GAstV-2 within a distinct clade, separate from astroviruses infecting humans, turkeys, and chickens. Comparative sequence alignments further revealed low genetic similarity, with nucleotide and amino acid identity below 65% relative to these viruses [15,28,29]. Moreover, GAstV-2 exhibits notable differences in pathogenicity. While turkey and human astroviruses primarily cause intestinal damage and diarrhea, GAstV-2 infection typically results in kidney injury in geese. The intestine may serve primarily as the entry site for GAstV-2, and the upregulation of tight junction-related genes likely represents a host defense response against viral invasion.
Cytokine production is a key component of the host immune response to viral infection and plays an important role in immune regulation. However, excessive cytokine release can lead to tissue damage and aggravation of clinical symptoms. In this study, GAstV-2 infection triggered robust production of proinflammatory cytokines, which may contribute to intestinal injury. Similar cytokine induction by GAstV-2 has been reported in the blood, kidneys, and spleen [14,30,31], indicating that this response is part of a broader systemic inflammatory reaction during infection.
5. Conclusions
In conclusion, following oral infection of goslings with GAstV-2, the virus initially targets the duodenum, localizing predominantly in intestinal crypts. This leads to crypt cell necrosis, reduced villus height, and decreased numbers of Lgr5+ ISCs, along with inhibition of ISC differentiation into goblet cells through modulation of the Notch signaling pathway. These changes, coupled with an enhanced inflammatory response, contribute to intestinal injury. Conversely, infected goslings exhibit increased numbers of Paneth cells and Bmi1+ ISCs, upregulation of tight junction–associated genes, and activation of the Wnt signaling pathway, which may enhance resistance to infection and promote epithelial regeneration. Collectively, these findings provide new insight into GAstV-2–host interactions in the intestine and advance our understanding of the pathogenic mechanisms underlying GAstV-2 infection.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ren D. Zhang H. Ye X. Jia X. Chen R. Tang T. Ye J. Wu S. Current situation of goose astrovirus in China: A review Viruses 2025178410.3390/v 1701008439861873 PMC 11768540 · doi ↗ · pubmed ↗
- 2Xu L. Jiang B. Cheng Y. He Y. Wu Z. Wang M. Jia R. Zhu D. Liu M. Zhao X. Infection and innate immune mechanism of goose astrovirus Front. Microbiol.202314112176310.3389/fmicb.2023.112176336778860 PMC 9909288 · doi ↗ · pubmed ↗
- 3Chen Q. Xu X. Yu Z. Sui C. Zuo K. Zhi G. Ji J. Yao L. Kan Y. Bi Y. Characterization and genomic analysis of emerging astroviruses causing fatal gout in goslings Transbound. Emerg. Dis.20206786587610.1111/tbed.1341031680474 · doi ↗ · pubmed ↗
- 4Wang A. Wu Z. Zhou Q. Zhang X. Zhu Y. Xie J. Feng Q. Dong H. Cheng Y. Jia W. Isolation, identification, and pathogenicity of a goose astrovirus 1 strain from goslings in Jiangsu province, China Microb. Pathog.202520010732410.1016/j.micpath.2025.10732439864762 · doi ↗ · pubmed ↗
- 5Chen L. Cui H. Li J. Zhang Y. Wang H. Yang Y. Wang X. Zhang C. Liu J. Epidemiological investigation of goose astrovirus in Hebei province, China, 2019–2021 Microorganisms 20241299010.3390/microorganisms 1205099038792819 PMC 11123679 · doi ↗ · pubmed ↗
- 6Bi Z. Lv X. Zhang Z. Cai L. Zhang M. Li W. Ding Y. Liu H. Yang K. Zhu Y. Emerging fatal gout disease in Chinese goslings linked to acute kidney injury induced by novel goose astrovirus infection Front. Cell Infect. Microbiol.20241414708083935993610.3389/fcimb.2024.1470808 PMC 11445172 · doi ↗ · pubmed ↗
- 7Zhang Q. Cao Y. Wang J. Fu G. Sun M. Zhang L. Meng L. Cui G. Huang Y. Hu X. Isolation and characterization of an astrovirus causing fatal visceral gout in domestic goslings Emerg. Microbes. Infect.20187712967472610.1038/s 41426-018-0074-5PMC 5908792 · doi ↗ · pubmed ↗
- 8Zhang X. Ren D. Li T. Zhou H. Liu X. Wang X. Lu H. Gao W. Wang Y. Zou X. An emerging novel goose astrovirus associated with gosling gout disease, China Emerg. Microbes. Infect.201871523018578610.1038/s 41426-018-0153-7PMC 6125322 · doi ↗ · pubmed ↗