Viruses Infecting Cuban Honey Bees and Evolution of Deformed-Wing-Virus Variants
Poppy J. Hesketh-Best, Anais R. Luis, Declan C. Schroeder, Stephen J. Martin

TL;DR
This study explores the evolution of deformed wing virus (DWV) in Cuba's isolated honey bee population, revealing unique viral patterns similar to those in the USA and Europe.
Contribution
The study reports the first detection of Lake Sinai Virus variants and the dominance of DWV-B in Cuba, highlighting unique viral evolution in an isolated bee population.
Findings
Two variants of Lake Sinai Virus were detected in Cuban honey bee samples.
DWV-B is the dominant variant in Cuba, unlike neighboring regions where DWV-A is more common.
A DWV-B/A recombinant was identified, suggesting viral evolution in Cuba mirrors that of the USA and Europe.
Abstract
Cuba is in a unique situation in which it has a large (220,000 managed colonies) and isolated honey bee population due to a 60+ year ban on the importation of bees. Despite this, the ectoparasitic mite Varroa destructor arrived in 1996, and with it came deformed wing virus (DWV). In 2018, an island-wide survey detected varroa and DWV in 91% of colonies. In this study, we conducted a full-virome analysis on some of these samples, along with additional samples collected in 2021. For the first time, we detected two variants of Lake Sinai Virus and confirmed the absence of the normally widespread black queen cell virus in Cuba. We also detected both DWV-A and DWV-B master variants, with DWV-B being the dominant variant. Interestingly, the DWV-B/A recombinant was also detected, indicating that despite Cuba’s isolated nature, the pattern of DWV evolution mirrors that found in the USA and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
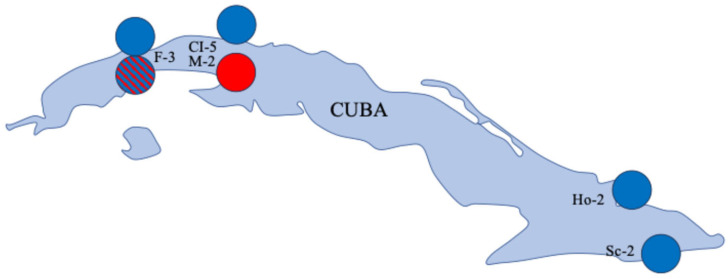 Figure 1
Figure 1 Figure 2
Figure 2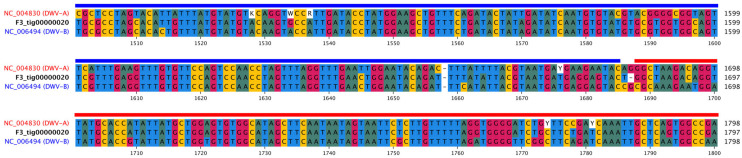 Figure 3
Figure 3- —Leverhulme
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect and Pesticide Research · Plant Virus Research Studies · Insect and Arachnid Ecology and Behavior
1. Introduction
Cuba, the largest Caribbean Island, is currently home to over 220,000 managed Apis mellifera (Apidae, Hymenoptera) honey bee colonies, in addition to a large free-living population [1]. An over-60-year ban on the importation of honey bees into Cuba has prevented the arrival of Africanized bees from neighboring countries. Studies using allozyme markers [2] and mitochondrial haplotypes (microsats) [3] have confirmed that the Cuban honey bee population is European (M and C lineages). The microsatellite data also showed that there is a homogeneous population of managed honey bees across the country without any regional differences, confirming the isolated nature of the population.
Despite the ban, in 1996, the ectoparasitic varroa mite, Varroa destructor (Varroidae, Mesostigmata), was first detected in the Matanzas province of Western Cuba and subsequently spread across the entire island. After initial losses of both managed and free-living colonies, natural varroa resistance arose, so no mite control methods have been used over the past 20+ years. This makes the Cuban population the largest varroa-resistant European honey bee population in the world [1]. Honey bees in Cuba [1], like those in many other countries [4], reduce the number of varroa mites by increasing their ability to detect and remove mite-infested worker cells [5]. This reduces the viral load within the colony due to there being fewer viral vectors (mites) [6].
An island-wide survey conducted in 2018 detected deformed wing virus (DWV, Iflavirus aladeformis, Iflaviridae, Picornavirales) in 91% of the analyzed colonies and 100% of the apiaries [7]. DWV has transformed from a minor honey bee pathogen to the most widespread and intensively studied insect pathogen in the world [8]. The dramatic rise in DWV’s prevalence is solely due to its association with the varroa mite. The mite introduced a new viral transmission route, i.e., inoculation directly into the hemolymph during mite feeding rather than the natural DWV transmission via food and mating. The lethal association between varroa and DWV has caused the death of millions of honey bee colonies and changed the viral landscape pertaining to honey bees. DWV, when transmitted by varroa, reduces the life expectancy of the infested honey bee pupae and leads to colony death when sufficient mite numbers are present. DWV is a multi-strain virus comprising four master variants, the common DWV-A and DWV-B variants, the rare DWV-C variant [8], and the potentially extinct DWV-D variant [9]. The master variant DWV-D was isolated from a dead pre-varroa colony from Egypt in 1977, and despite extensive searches of several hundred RNA-sequencing libraries, it now appears to have become extinct [9]. Its closest-related master variant is DWV-C, which has only been detected occasionally in viral surveys of varroa-infested honey bee colonies [10]. It is more common in stingless bees [11], small hive beetles [12], and, interestingly, in two varroa-free island populations of honey bees in the Azores [13]. In varroa-infested honey bees, the two major master variants, DWV-A and DWV-B, are almost always detected in honey bee surveys [8,14]. DWV-A was the first master variant to be detected in varroa-infested colonies [15]. However, longitudinal studies [10,16,17,18] have shown that the DWV-B master strain initially replaced DWV-A. During this process, various DWV-B/A recombinants emerged, which now appear to be dominant in both the US [19] and Europe [20]. In Latin America [11,18], China [21], and Japan [22], the DWV-A master variant remains dominant.
Here, we provide the first report of which DWV master variants are present in the isolated Cuban honey bee population.
2. Materials and Methods
Twelve samples, each containing around 50 adult workers, were collected from across Cuba in 2018 [7] and supplemented by new samples collected 2021 and stored in a freezer before being prepared for Oxford Nanopore Technologies (ONT) sequencing, as described in [19]. Briefly, DNA libraries were generated using Template-Switching RT Enzyme Mix (New England Biolabs, Ipswich, MA, USA) with an N6 TS–modified random primer (Thermo Fisher Scientific, Waltham, MA, USA) for first-strand synthesis and PrimeSTAR GXL polymerase (Takara Bio, San Jose, CA, USA) for second-strand synthesis [23,24]. Clean cDNA was then used to prepare Oxford Nanopore Technologies (ONT) sequencing libraries following the Native Barcoding Sequencing Kit (SQK-NBD114-96) protocol, with FPPE DNA Repair omitted. Pooled libraries were sequenced for 24 h on an R10 flow cell using a GridION (Oxford Nanopore Technologies, New York, NY, USA).
Reads were base-called and demultiplexed with Guppy v6.4 (high-accuracy model). Nanopore barcodes and adapters were trimmed using PoreChop v0.2.4. [25]. Trimmed reads were corrected and assembled with Canu v2.2 [26] using the following flags: nanapore maxInputCoverage = 2000 corOutCoverage = all corMinCoverage = 10 corMhapSensitivity = high minoverlap = 50 minread = 200 genomesize = 5000. Contigs ≥ 2 kb were manually binned in the anvi’o v7.1 interactive interface [27]. Anvi’o was used to profile contigs using Prodigal v2.6.3 [28]. Reads were mapped with Minimap2 v2.24 [29], and BAM files were generated with SAMtools v1.19.2 [30]. Coverage profiles were merged into a single database. Additional functional and taxonomic information was added using HMMER v3.4 against VOGs (https://vogdb.org/, accessed 31 December 2024), along with standard anvi’o HMMs, NCBI COGs, and KEGG Kofams [31,32]. Gene-level taxonomy was assigned using Kaiju v2.9 against the NCBI nr and RVDB databases (accessed 31 December 2023) [33]. Manual binning was guided by sequence composition (anvi’o dendrogram) and VOG HMMER hits.
3. Results
Due to RNA degradation caused by regular power cuts to the freezers in Cuba, we were only able to assemble eight (contigs) from the high-quality sequences obtained from the five samples, but these were located across Cuba (Table 1 and Figure 1), with contig lengths of between 2036 and 8448 (Table 1). Overall, the genomes obtained were fragmented, likely due to RNA degradation, limiting the completeness of the assemblies. We successfully sequenced a near-complete DWV-A genome in one sample (M2), DWV-B in four samples, and a DWV-B/A recombinant co-infection with DWV-B in one sample (Table 1, Figure 1 and Figure 2). The putative breakpoint for the recombinant genome occurred within the GP1 region (Figure 3). In addition, in one sample (CI-5, Table 1), we also detected two variants of lake Sinai virus (LSV, Sinhaliviridae, Nodamuvirales) for the first time in Cuba. No black queen cell virus (Dicistroviridae, Cripavirus) was detected in any of the samples.
4. Discussion
In this study, we did not detect black queen cell virus, which was detected in a previous viral survey of Cuba [7]. This is unusual since this virus is one of the most globally detected viruses in honey bee surveys [34,35]. Its absence may again reflect the isolated nature of the Cuban honey bee population. For the first time, we detected LSV in Cuba, which, unlike DWV, appears not to be an emerging disease but rather a highly variable multi-strain virus that has a stable association with A. mellifera [36]. The dominance of the DWV-B variant and the presence of the DWV-B/A recombinant in Cuban honey bees mirror the situation that was previously observed in Europe [20,37], the USA [19], and many other countries [14], with the DWV-A variant being replaced by DWV-B. The recombinant DWV-B/A sample detected in Cuba had DWV-B structural genes (VP1-VP4), meaning that previous RT-qPCR assays, e.g., [11], targeting the RdRP gene incorrectly indicated the presence of DWV-A rather than DWV-B/A recombinants. Interestingly, in all countries except Japan [22], where DWV-A is currently dominant, DWV-B has been detected but at very low prevalences, such as in Brazil (11%, n = 26) [11], Mexico (0.7–2%, n = 364) [18], and China (0.8%, n = 117) [38]. Although this study is based on only five samples, DWV-B was detected in 75% of the samples, with similar high prevalences of DWV-B detected in the UK (100%, n = 249) [10], US (56%, n = 217) [10], and Europe (82%, n = 50) [20]. This provokes us to ask why and how DWV-A has remained dominant over many decades in some countries and why it has been replaced by DWV-B, a development followed by the rise in DWV-B/A recombinants in other regions such as the USA and Europe. One proposed explanation is that superinfection exclusion by DWV-A may block DWV-B via the process of inter-genotype recombination meltdown [18], but this idea needs to be empirically tested.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Luis A.R. Grindrod I. Webb G. Perez A. Martin S.J. Recapping and mite removal behaviour in Cuba: Home to the world’s largest population of Varroa-resistant European honeybees Sci. Rep.2022121559710.1038/s 41598-022-19871-536114263 PMC 9481617 · doi ↗ · pubmed ↗
- 2Pérez A. Demedio J. Estatus racial e índices de infestación de colmenas (Apis mellifera L.) por Varroa destructor (Anderson y Trueman) en Mayabeque, Cuba. [Racial status and index of hive (Apis mellifera L.) infestation by Varroa destructor (Anderson and Trueman) in Mayabeque, Cub]Cuban J. Agric. Sci.2017512
- 3Yadró-Garcia C.A. Rodríguez-Luis A. Pérez-Pineiro A.M. Pérez-Morfi A. Invernizzi C. Tomasco I.H. Cuban honey bees: Significant differentiation from European honey bees in incomplete isolation J. Apicult. Res.20216037538410.1080/00218839.2020.1841460 · doi ↗
- 4Martin S.J. Hawkins G.P. Brettell L.E. Reece N. Correia-Oliveira M.E. Allsopp M.H. Varroa destructor reproduction and cell re-capping in mite-resistant Apis mellifera populations Apidologie 20205136938110.1007/s 13592-019-00721-9 · doi ↗
- 5Grindrod I. Martin S.J. Parallel Evolution of Varroa Resistance in Honey Bees; a common mechanism across continents?Proc. R. Soc. B 20212882021137510.1098/rspb.2021.1375 PMC 833483934344183 · doi ↗ · pubmed ↗
- 6de Souza F.S. Allsopp M.H. Martin S.J. Deformed wing virus prevalence and load in honeybees in South Africa Arch. Virol.202116623724110.1007/s 00705-020-04863-533136209 PMC 7815608 · doi ↗ · pubmed ↗
- 7Luis A.R. Garcia C.A.Y.G. Invernizzi C. Branchiccela B. Piñeiro A.M. Morfi A.P. Zunino P. Antúnez K. Nosema ceranae and RNA viruses in honey bee populations of Cuba J. Apicult. Res.20205946847110.1080/00218839.2020.1749451 · doi ↗
- 8Martin S.J. Brettell L.E. Deformed wing virus in honeybees and other insects Ann. Rev. Virol.20196496910.1146/annurev-virology-092818-01570031185188 · doi ↗ · pubmed ↗