Histopathologic and Genomic Characterization of a Novel Caprine Astrovirus Identified in a Boer Goat Kid in Illinois, United States
Jingyi Li, Wes Baumgartner, Leyi Wang

TL;DR
A new astrovirus was found in a sick goat in the U.S., causing severe intestinal damage and showing genetic links to viruses in Asia and Europe.
Contribution
This is the first report of a caprine astrovirus in the U.S. associated with gastrointestinal disease in a goat.
Findings
The virus was identified in a Boer goat kid with severe necrotizing enterocolitis.
The virus showed 86% nucleotide identity to a sheep astrovirus strain from China.
Phylogenetic analysis grouped the virus with small ruminant astroviruses from Asia and Europe.
Abstract
Astroviruses are non-enveloped, positive-sense single-stranded RNA viruses known to infect various mammals and birds, including humans, often causing gastrointestinal disorders. In recent years, astroviruses have also been linked to neurological and respiratory diseases across several species, including ruminants, mink, deer, and other mammals. Notably, astrovirus infections in goats have been documented in countries such as Switzerland and China, where novel genotypes have been identified in fecal samples. However, their role in the context of disease remains unclear, and reports focusing solely on goat astrovirus in the United States have not been published. A necropsy case of a Boer goat kid with a history of diarrhea was submitted for investigation following death in January 2025. Fresh tissues were received and used for histopathology and enteric pathogen testing, including…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
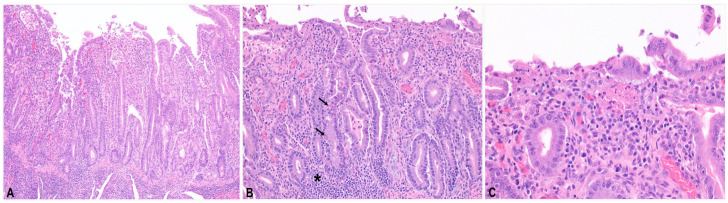 Figure 1
Figure 1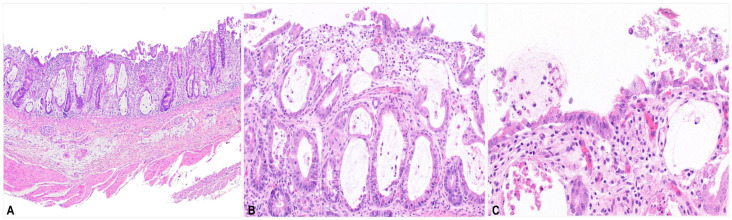 Figure 2
Figure 2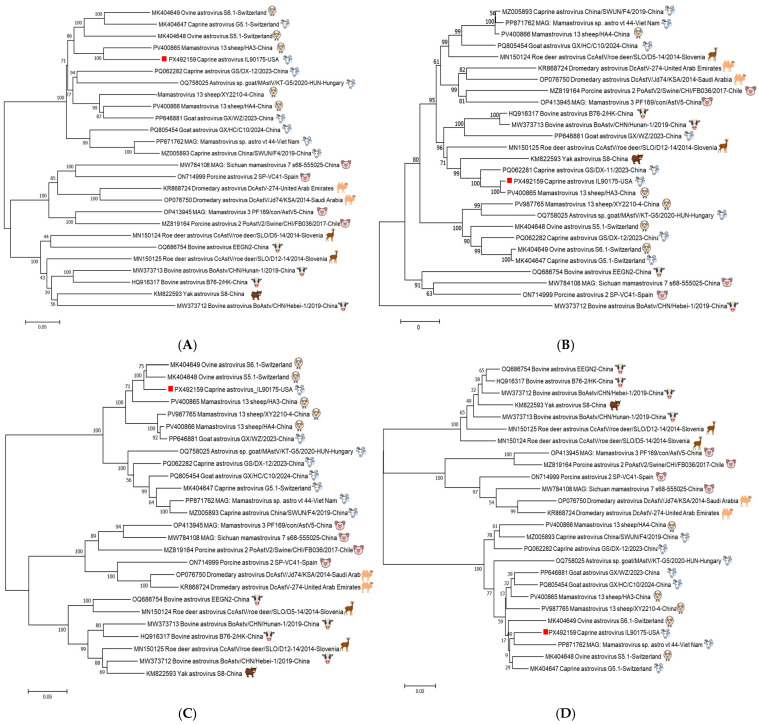 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsViral gastroenteritis research and epidemiology · Virus-based gene therapy research · Viral Infections and Outbreaks Research
1. Introduction
Astroviruses are small, non-enveloped, positive-sense single-stranded RNA viruses belonging to the family Astroviridae [1]. Within the family, the two genera Mamastrovirus and Avastrovirus encompass viruses infecting mammals and birds, respectively [2]. The genome of astroviruses is approximately 6–8 kb long, contains 5′ and 3′ untranslated regions, and three open reading frames (ORFs): ORF1a, ORF1b, and ORF2. The 5′ end is not capped, and ORF1a and ORF1b encode the astrovirus nonstructural proteins, including a serine protease, a viral genome-linked protein (VPg), and an RNA-dependent RNA polymerase (RdRp). At the 3′ poly (A) tail, ORF2 encodes the astrovirus capsid protein [3].
Human astrovirus in children with diarrhea was first described in 1975 [4]. Since then, more cases have been reported in a variety of species, including sheep, cattle, chickens, pigs, dogs, cats, red deer, ducks, mice, turkeys, mink, guinea fowl, bats, cheetahs, sea lions, and rats [1]. Astrovirus infection in different hosts is associated with gastroenteritis, nephritis, hepatitis, encephalitis, polioencephalomyelitis syndrome, and respiratory illness [1,5,6,7].
Transmission primarily occurs via the fecal–oral route. In most species, including humans and sheep, infections are typically asymptomatic or cause mild, self-limiting diarrhea, but in rare cases, infections may lead to severe diarrhea [8]. In turkeys, for example, astrovirus infection is associated with increased mortality and severe diarrhea, despite only mild histopathological lesions [9]. However, the causal relationship between astrovirus infection and diarrhea remains controversial in many species, in that astroviruses have been detected in both healthy and diarrheic animals, namely cattle, goats, roe deer, and pigs [10,11,12,13]. Nevertheless, a 2021 study demonstrated a strong correlation between astrovirus infection and diarrhea in cattle, both as a single agent and during co-infections [14]. In addition to gastrointestinal disease, astroviruses have also been implicated in neurological disorders in animals such as cattle and sheep, encephalomyelitis in pigs [2,15,16,17], as well as “shaking mink syndrome” [18]. Viral respiratory disease has been reported in white-tailed deer [6], pigs [19], and cattle [5].
Caprine astrovirus was identified initially in goat fecal samples from Switzerland in 2019, where three genotypes were discovered: caprine astrovirus G5.1, caprine astrovirus G3.1, and MAstV–34 [20]. In 2021, a novel genotype was detected in goats in China [21]. Subsequent surveillance in China in 2022 and 2024 identified three additional genotypes from goat feces (MAstV–13 and MAstV–24), both with unique genetic and epidemiological profiles [11,22]. These discoveries highlight the circulation of multiple, genetically diverse caprine astrovirus genotypes worldwide. To date, no research has characterized the genetics of goat astrovirus and their correlation with gastrointestinal disease in the United States. In this study, we report the first identification of a novel caprine astrovirus in the United States.
2. Materials and Methods
2.1. Necropsy Samples
In January 2025, necropsy tissues (lung, heart, spleen, kidney, small and large intestines, mesenteric lymph node, abomasum, skeletal muscle, rumen) from a 4-week-old female Boer goat kid were submitted to the University of Illinois Urbana–Champaign Veterinary Diagnostic Laboratory (UIUC-VDL) for histopathological examination and microbiological identification.
2.2. Histopathology Preparation
The necropsy tissues were fixed in 10% neutral buffered formalin, trimmed into cassettes, and routinely processed. All tissues were evaluated with routine hematoxylin and eosin (H&E) staining.
2.3. Routine Laboratory Testing
Both the gross lesions and histopathological findings suggested infectious causes, and intestinal tissues were submitted for pathogen testing, including bacterial culture, Salmonella PCR (DuPont Qualicon BAX^®^ System, Hygiena, Wilmington, DE, USA), rotavirus antigen assay (SAS™ Rota Test, SA Scientific, Ltd., San Antonio, TX, USA), and fecal flotation (Sheather’s solution) for parasites. Both Salmonella PCR and rotavirus antigen assay were performed following the manufacturer’s directions.
2.4. Intestinal Content Suspension Preparation and Nucleic Acid Extraction
Nucleic acids were extracted from intestinal content. The intestine lumen was swabbed with a cotton-tipped wooden applicator. The swab tip was then placed in a microcentrifuge tube containing 1000 µL PBS buffer, agitated, and removed. The tube was vortexed for 20 s and centrifuged for 2 min at 6000× g and room temperature. The nucleic acid extraction was performed using the MagMAX Pathogen RNA/DNA Kit (ThermoFisher, Waltham, MA, USA) on a KingFisher Flex machine (ThermoFisher, Waltham, MA, USA) [23].
2.5. Metagenomics-Based and Targeted-Based Next-Generation Sequencing (NGS), Assembly, and Analysis
2.5.1. Metagenomics-Based NGS (mNGS)
The extracted nucleic acid was subjected to sequence-independent, single primer amplification (SISPA) as previously described [24]. It was first reverse transcribed into complementary DNA (cDNA) using Superscript III reverse transcriptase (ThermoFisher, Waltham, MA, USA) with a random octamer primer containing a defined sequence tag. Double-stranded DNA (dsDNA) was then synthesized using Klenow polymerase (NEB, Ipswich, MA, USA). Subsequent amplification was carried out with a single primer corresponding to the sequence tag using the Advantage 2 PCR kit (Takara Bio, Ann Arbor, MI, USA). The resulting PCR amplicons were purified with the QIAquick PCR Purification Kit (QIAGEN, Germantown, MD, USA) and quantified with the Qubit high-sensitivity assay kits (ThermoFisher, Waltham, MA, USA).
Library preparation was performed using the Nextera XT DNA Library Preparation Kit (Illumina, San Diego, CA, USA) according to the manufacturer’s instructions. The procedure included tagmentation, PCR amplification, PCR cleanup, library normalization, and final pooling. The sample was sequenced using an Illumina MiSeq v2 sequencing kit (300 cycles) on MiSeq (Illumina, San Diego, CA, USA).
2.5.2. Targeted-Based NGS (tNGS)
The assembled genome had two contigs with a 61 bp gap within the capsid region. To close the gap and confirm the capsid sequence, three pairs of primers were designed and used for amplicon-based sequencing of the whole capsid region (Table 1). One-step RT-PCR was performed using the SuperScript™ III One-Step RT-PCR System with Platinum™ Taq DNA Polymerase (ThermoFisher, Waltham, MA, USA) and 5 µL RNA in a 25 µL reaction volume. Amplicon was purified using the QIAquick PCR Purification Kit (QIAGEN, Germantown, MD, USA) and sequenced on MiSeq as described in Section 2.5.1.
2.5.3. Bioinformatic Sequence Analysis
Raw FASTQ files from MiSeq were initially analyzed using Kraken taxonomic classification software (Kraken 2 version) [25] using a database built through a standard method and then assembled using the de novo SPAdes assembler (v4.0.0) [26]. The resulting contigs were analyzed through a local BLAST (BLASTn and BLASTx, https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 1 March 2025) search to identify viral sequences. Using the online closest astrovirus strain S6.1 (MK404649) as the reference, reference-based mapping and extraction of consensus sequences of astrovirus were performed using CLC Genomics Workbench (Qiagen, Germantown, MD, USA). Sequence alignment was performed using MAFFT version 7 [27], and phylogenetic tree construction was carried out with Molecular Evolutionary Genetics Analysis (MEGA) version 7 [28], while sequence identity calculations were performed using the BioEdit alignment editor (version 7.0.5.3).
3. Results
3.1. Clinical Signs in Affected Goat Kids
The animal had a history of light yellow, watery diarrhea. Several other goat kids on the same farm had also died exhibiting similar signs. The referring veterinarian reported scant, watery intestinal contents within the small intestine and no other gross lesions.
3.2. Histopathology
Microscopically, the small and large intestines exhibited severe necrotizing enterocolitis. In the small intestine (Figure 1), the villi were markedly blunted and crypts were hyperplastic. The epithelium showed multifocal ulcerations with eosinophilic and karyorrhectic debris, pyknotic or karyorrhectic enterocytes and leukocytes, and fine cocci bacteria. Few coccidian intraenterocytic parasites were present in the crypts, ranging from schizonts to zoites. Many crypts were dilated by necrotic debris and eosinophilic mucoid material. The lamina propria contained infiltrates of macrophages, lymphocytes, and plasma cells.
In the large intestine (Figure 2), the mucosa was irregular and thin with ulcerations and erosions. Eosinophilic and karyorrhectic debris, mucus, and colonies of fine cocci bacteria occurred along the surface. Crypts were irregular and often dilated by mucus and necrotic enterocytes and lined by either necrotic or hyperplastic epithelium. Many necrotic cells had vacuolated cytoplasm. The lamina propria was fibrotic with macrophages, lymphocytes, and plasma cells scattered throughout. Other tissues lacked significant lesions.
3.3. Routine Laboratory Testing Results
Fecal flotation did not reveal organisms. There was heavy growth of Escherichia coli and Enterococcus hirae. Salmonella PCR and rotavirus antigen assays were negative (Table 2).
3.4. Metagenomic NGS (mNGS) and Targeted NGS (tNGS) and Genome Analysis
3.4.1. Identification of Goat Astrovirus with Complete Genome
Kraken taxonomical analysis of mNGS data revealed that astrovirus reads (14% of total reads, 111,252 reads in total belonging to Astroviridae) were present in the intestinal content (Supplemental Figure S1). The assembled sequence of goat astrovirus IL90175 has two contigs with a 61 bp gap in the capsid gene, as compared to other genomes available on GenBank. In March of 2025, an online NCBI BLAST of the assembled IL90175 astrovirus nucleotide sequence showed that IL90175 had the highest identity (84%) to an ovine astrovirus S6.1 strain from Switzerland with 87% query coverage and 78% identity to a goat strain GS/DX-11/2023 from China (this strain only had capsid gene sequence at GenBank, PQ062281) (Supplementary Figure S2). Further amplicon-based tNGS generated the complete capsid sequence, confirming the capsid sequence derived from mNGS. The complete genome sequence of caprine astrovirus IL90175 strain is 6283 bp in length and deposited into GenBank (accession number: PX492159) and was used for phylogenetic and identity analysis.
3.4.2. Genomic Analysis
In November of 2025, an online NCBI BLAST of the complete genome sequence indicated that caprine astrovirus IL90175 had the highest nucleotide identity (86%) with a newly deposited sheep astrovirus strain, sheep/HA3 (PV400865), with 100% query coverage. This was higher than the identities for the second and third hits: 84% to an ovine astrovirus strain S6.1 (MK404649) and 83% to strain S5.1 (MK404648). Both of these hits showed only 87% query coverage. Whole-genome nucleotide identity analysis using BioEdit revealed that IL90175 shared less than 80% identity with all strains except the highest-identity match, sheep/HA3 (Table 3).
Further analysis of the capsid amino acid sequence using BioEdit showed that caprine astrovirus IL90175 had the highest identity (96.3%) with sheep/HA3, followed by GS/DX-11/2023 (PQ062281) at 86.2%, but markedly lower identities with two ovine strains, S5.1 (62.8%) and S6.1 (60.4%), from Switzerland (Table 3). By contrast, in ORF1a, caprine astrovirus IL90175 showed the highest amino acid sequence identities (94.3%) with both ovine strains S5.1 and S6.1, followed by sheep/HA3 at 91.5%. In ORF1b, IL90175 shared the highest identity with the goat astrovirus G5.1 from Switzerland (96.4%), followed by sheep/HA3 (96.2%) (Table 3).
Phylogenetic tree analysis showed that IL90175 clustered together with the China sheep/HA3 strain for both complete nucleotide and capsid amino acid sequences (Figure 3A,B). However, IL90175 was more closely related to two sheep strains from Switzerland, S5.1 and S6.1, in the ORF1a amino acid phylogeny, and to the Vietnam goat strain astro_vt_44 in the ORF1b amino acid phylogeny (Figure 3C,D).
4. Discussion
Astroviruses are associated with a broad spectrum of diseases: enteric [4,8,9], respiratory [6,19], and neurologic [2,15,16,17,18] across various species. Diarrhea is a major cause of morbidity and mortality in ruminants, particularly in neonates, characterized by dehydration, weight loss, and, in severe cases, death. The description of gastrointestinal disease caused by caprine astrovirus is limited. A study by Wang et al. (China) reported no significant difference in astrovirus detection rates between diarrheic and non-diarrheic samples, and lesions were not described [11]. Additionally, genome characterization of caprine astrovirus is sparse, with only a few strains reported in Switzerland and China in 2019, 2021, 2022, and 2024 [11,20,21,22]. To date, there have been no reports of any genome identification of goat astrovirus in the United States. This study represents the first genome characterization of caprine astrovirus associated with gastrointestinal disease in the U.S.
In this case, co-infection was evident, including coccidia and E.coli, which are associated with neonatal diarrhea. The contribution of E. hirae to disease is uncertain, as it is uncommonly reported, although a heavy culture growth is seen; this organism is likely an opportunist. The histopathology findings were not typical of bacterial and coccidian infections and suggested viral disease, which spurred metagenomic investigation. It is plausible that the pathogens in this case may play contributory or synergistic roles in disease pathogenesis. Histologic descriptions of astrovirus infection are rarely documented in the literature, especially in ruminants. A study conducted in 1979 by D. R. Snodgrass et al. described histologic lesions in astrovirus-infected lambs, which were limited to mild small intestine lesions and resolved by 5 days post-infection [8]. The villi were atrophied, the crypts were dilated, the enterocytes were lost or necrotic, and the lamina propria was expanded by moderate numbers of macrophages, neutrophils, and lymphocytes [8]. In pigs, the lesions were also found in the small intestine, with villous atrophy, crypt hyperplasia, and neutrophils and macrophages in the villi [29]. Similar histological lesions were observed in our case; however, our findings were more severe, with lesions in the large intestine. Further characterization using in situ hybridization techniques and additional sample examination will better characterize this disease and is being pursued.
New virus/variant discoveries often encounter a significant bottleneck in bioinformatic analysis. The alignment of the initial genome sequence using NCBI online BLAST revealed a large gap in the capsid region of IL90175; however, it mapped to the capsid protein gene of a goat strain, GS/DX-11/2023 (Figure S2). Moreover, mapping raw FASTQ data from mNGS to the closest strain, ovine astrovirus S6.1, using CLC Genomics Workbench revealed a large gap in the capsid region (Figure S3A). To close the sequence gap and verify the capsid sequence, amplicon-based targeted NGS was performed to obtain the complete capsid gene. Using the Qiagen CLC Genomics Workbench, mapping of the raw FASTQ data from mNGS to the complete genome of IL90175 revealed high coverage across the entire genome, including the capsid protein region (Figure S3B). When the closest reference strain sequence was distantly related to the raw FASTQ sequence data, not all astrovirus sequences in the raw reads mapped to the reference (ovine astrovirus S6.1), resulting in gaps in highly divergent regions. These results highlighted the limitations of the online NCBI BLAST program and CLC genomics workbench mapping when analyzing highly divergent sequences.
Recent reports increasingly show that mutational events and recombination events are commonly observed between and within the astrovirus genotypes, especially sheep and goat astroviruses [21,30,31]. In our study, using both NCBI BLAST and BioEdit, the complete genome showed the highest sequence identity to the Mamastrovirus 13 sheep strain recently reported in China in April 2025. Prior to this release, our sequence showed the greatest similarity to the ovine astrovirus S6.1 strain from Switzerland. In the phylogenetic tree, the genome clustered closely with Mamastrovirus 13 sheep and other ovine and caprine strains from Switzerland, China, and Hungary. This finding suggests that the U.S. caprine astrovirus strain shares a common evolutionary origin with astroviruses circulating in Europe and Asia, potentially reflecting cross-species transmission or the global spread of related lineages. In addition to whole-genome comparisons, the capsid amino acid is not only related to caprine astrovirus in China but also closely related to Mamastrovirus 13 in sheep in China, yak astrovirus in China, and roe deer astrovirus in Slovenia. The capsid protein encoded by ORF2 is the most variable region in the genome of mammalian and avian astroviruses, and frequent interspecies transmission and recombination, especially near the ORF1b/ORF2 junction, can disrupt host-specific clustering [32]. The nonstructural genes encoded by ORF1a and ORF1b (including the viral protease and RNA-dependent RNA polymerase) are relatively conserved across related host species [11,30]. The ORF1a/b region in our study supports this concept, clustering with ovine and caprine astroviruses from China, Switzerland, and Vietnam.
In summary, this study reports the first detection of caprine astrovirus in the United States, representing a novel strain of the virus. The affected diarrheic goat exhibited chronic necrotizing enteritis and colitis. However, a significant correlation between astrovirus infection and gastrointestinal disease in goats requires further characterization, as secondary bacterial and protozoal infections were also present in this case. Additional case investigations using RNAscope, broader pathogen screening, continued histopathological analysis, and genome characterization would help clarify the causal relationship between neonatal gastrointestinal disease and astrovirus and reveal the genetic characteristics and diversity of the virus.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1De Benedictis P. Schultz-Cherry S. Burnham A. Cattoli G. Astrovirus infections in humans and animals-molecular biology, genetic diversity, and interspecies transmissions Infect. Genet. Evol.2011111529154410.1016/j.meegid.2011.07.02421843659 PMC 7185765 · doi ↗ · pubmed ↗
- 2Li L. Diab S. Mc Graw S. Barr B. Traslavina R. Higgins R. Talbot T. Blanchard P. Rimoldi G. Fahsbender E. Divergent astrovirus associated with neurologic disease in cattle Emerg. Infect. Dis.2013191385139210.3201/eid 1909.13068223965613 PMC 3810933 · doi ↗ · pubmed ↗
- 3Cortez V. Meliopoulos V.A. Karlsson E.A. Hargest V. Johnson C. Schultz-Cherry S. Astrovirus Biology and Pathogenesis Annu. Rev. Virol.2017432734810.1146/annurev-virology-101416-04174228715976 · doi ↗ · pubmed ↗
- 4Appleton H. Higgins P.G. Letter: Viruses and gastroenteritis in infants Lancet 19751129710.1016/S 0140-6736(75)92581-748925 · doi ↗ · pubmed ↗
- 5Ng T.F. Kondov N.O. Deng X. Van Eenennaam A. Neibergs H.L. Delwart E. A metagenomics and case-control study to identify viruses associated with bovine respiratory disease J. Virol.2015895340534910.1128/JVI.00064-1525740998 PMC 4442534 · doi ↗ · pubmed ↗
- 6Wang L. Shen H. Zheng Y. Schumacher L. Li G. Astrovirus in White-Tailed Deer, United States, 2018 Emerg Infect Dis 20202637437610.3201/eid 2602.19087831961307 PMC 6986847 · doi ↗ · pubmed ↗
- 7Arruda B. Arruda P. Hensch M. Chen Q. Zheng Y. Yang C. Gatto I.R.H. Ferreyra F.M. Gauger P. Schwartz K. Porcine Astrovirus Type 3 in Central Nervous System of Swine with Polioencephalomyelitis Emerg. Infect. Dis.2017232097210010.3201/eid 2312.17070329148383 PMC 5708247 · doi ↗ · pubmed ↗
- 8Snodgrass D.R. Angus K.W. Gray E.W. Menzies J.D. Paul G. Pathogenesis of diarrhoea caused by astrovirus infections in lambs Arch. Virol.19796021722610.1007/BF 01317493116623 PMC 7086881 · doi ↗ · pubmed ↗