Biochemical and Genetic Characterization of Ergot Alkaloid Biosynthesis in Aspergillus aspearensis
Jessica L. Fuss, Daniel G. Panaccione

TL;DR
A new fungus, Aspergillus aspearensis, produces ergot alkaloids, which are important as toxins and pharmaceuticals, and may be useful for research and production.
Contribution
Discovery of a new non-Clavicipitaceae fungus producing ergot alkaloids with functional gene clusters.
Findings
A. aspearensis produces lysergic acid α-hydroxyethylamide and other ergot alkaloids.
The fungus produces ergot alkaloids during insect pathogenesis and completes its life cycle in insects.
The genome contains two functional ergot alkaloid synthesis gene clusters without pseudogenization.
Abstract
Ergot alkaloids derived from lysergic acid have impacted humankind significantly as toxins in agriculture and as the foundations of several pharmaceuticals. Few fungi capable of producing lysergic acid derivatives have been found outside the family Clavicipitaceae. Based on its phylogenetic placement, we hypothesized the recently described fungus Aspergillus aspearensis (Aspergillaceae) would synthesize lysergic acid amides. Cultures of A. aspearensis produced abundant lysergic acid α-hydroxyethylamide (LAH) and lesser amounts of other lysergic acid derivatives. Conidia contained high concentrations of ergot alkaloids, whereas sclerotia contained significantly less. Approximately half of the ergot alkaloids produced were secreted into the culture medium. When spores of A. aspearensis were injected into larvae of the model insect Galleria mellonella, larvae died at a significantly faster…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
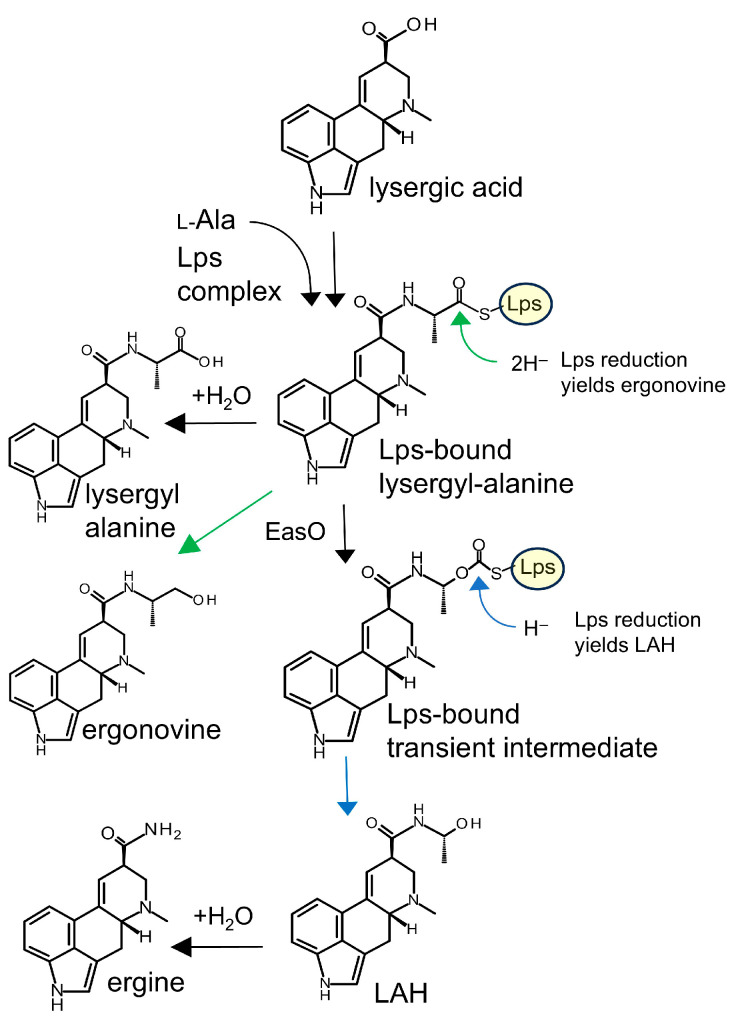 Figure 1
Figure 1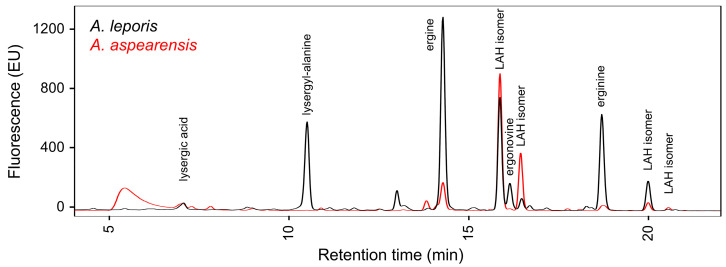 Figure 2
Figure 2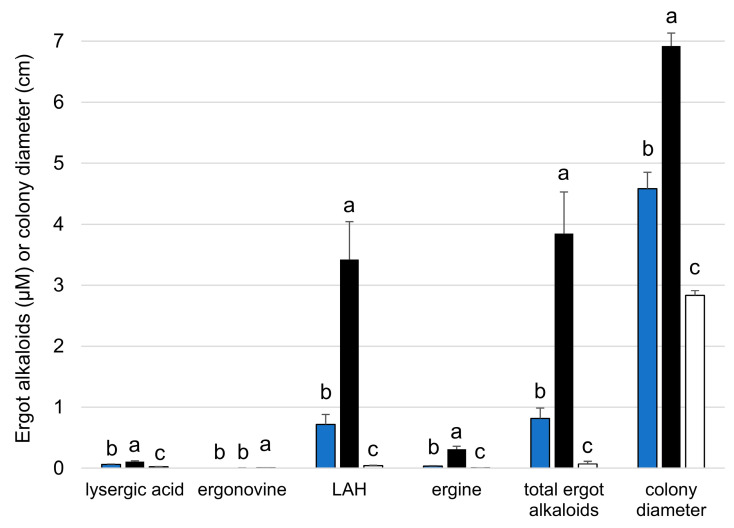 Figure 3
Figure 3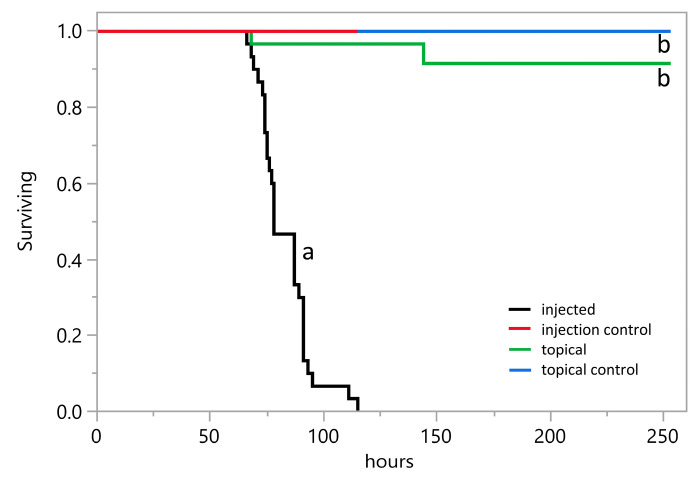 Figure 4
Figure 4 Figure 5
Figure 5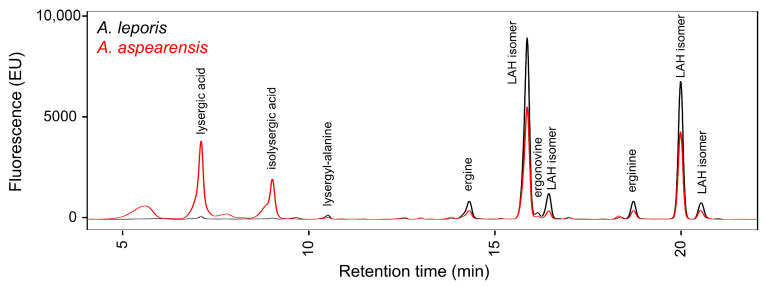 Figure 6
Figure 6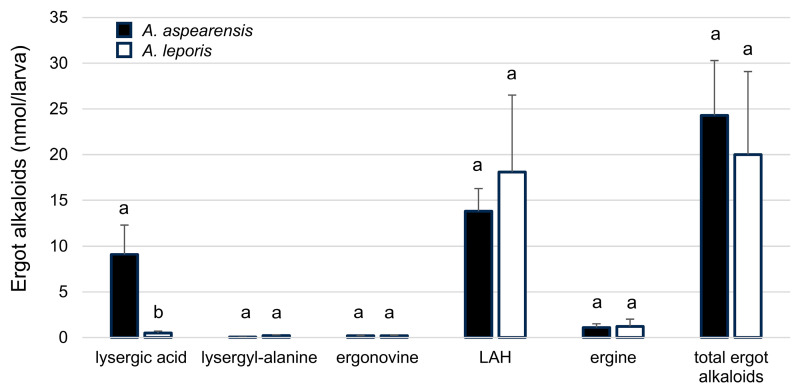 Figure 7
Figure 7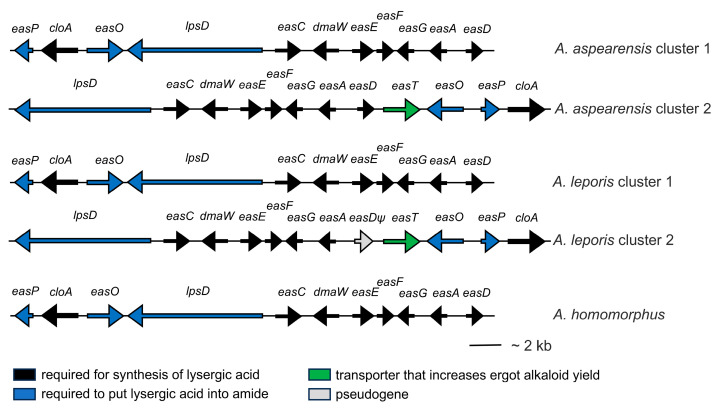 Figure 8
Figure 8- —National Institute of General Medical Sciences
- —National Institute of Food and Agriculture
- —Corinne Hazel (West Virginia University)
- —Clinical & Translational Science Institute (CTSI)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and fungal interactions · Botanical Research and Chemistry · Advanced Synthetic Organic Chemistry
1. Introduction
Ergot alkaloids produced by fungi are significant in agriculture and medicine in multiple ways. Ways in which ergot alkaloids affect agriculture include the historical poisoning of people through ingestion of ergot sclerotia of Claviceps purpurea that contaminated infected grain crops. Consumption of ergot alkaloids found in high concentrations in these fungal structures led to a condition called ergotism that resulted in significant human suffering and death in certain cultures through much of recorded history [1,2]. Similar ergot alkaloids produced by Epichloë species that are endophytic symbionts of grasses continue to plague animal agriculture by reducing animal growth, health, and reproduction [3,4]. Conversely, the anti-herbivore activities of ergot alkaloids reduce damage by insect pests of grasses associated with Epichloë symbionts [5,6] and also may be factors in the success of related Metarhizium species as insect biocontrol agents [7,8]. In medicine, ergot alkaloids have been the foundations of numerous pharmaceuticals prescribed to treat dementia, migraines, Parkinson’s disease, and a variety of other conditions [9,10,11]. The potent pharmacology of ergot alkaloids is exemplified in the activities of the semi-synthetic ergot alkaloid derivative LSD [12]. For these reasons, knowledge of the distribution of ergot alkaloid biosynthetic capability in fungi of different lineages and occupying different ecological niches is important.
Different classes of ergot alkaloids are produced from alternate branches of a highly complex pathway [10,11]. The class of ergot alkaloids most closely associated with the toxicological and pharmaceutical properties described above comprises the lysergic acid-derivatives. The Claviceps, Epichloë, and Metarhizium species discussed above are all members of the family Clavicipitaceae and all produce lysergic acid-derived ergot alkaloids from a shared pathway encoded by genes comprising a biosynthetic gene cluster referred to as an eas cluster [11,13]. Outside of fungi in the Clavicipitaceae, the ability to produce lysergic acid-derived ergot alkaloids has only been documented in four fungi representing two additional, diverse orders of fungi. These include three species of Aspergillus (order Eurotiales) [14] and a distantly related species of Dicyma (order Xylariales) [15]. Molecular and bioinformatic analyses of the three Aspergillus species revealed independent evolution of some pathway steps and a gene with novel biosynthetic capacity [14]. No genetic data are available for the Dicyma species. The Aspergillus species accumulate the lysergic acid amides ergonovine and lysergic acid α-hydroxyethylamide (LAH), along with hydrolysis products lysergyl-alanine and ergine (Figure 1), with LAH typically being produced in the greatest abundance.
The three Aspergillus species documented to produce lysergic acid-derived ergot alkaloids include: A. leporis, which was originally isolated from rabbit dung in Wyoming [16], a western US state, and also has been found in the rhizospheres of plants in that same geographic region [17]; A. hancockii from agricultural soils and commodities in Australia [18]; and, A. homomorphus from soil near the Dead Sea of Israel [19] as well as on millet in Yemen [20]. Aspergillus leporis and A. hancockii are close relatives in Aspergillus section Flavi, whereas A. homomorphus is in a separate Aspergillus section, Nigri. A noteworthy difference in the eas clusters of the Aspergillus species compared to those of the Clavicipitaceae is the inclusion of a single, two-module peptide synthetase-encoding gene in the Aspergillus species as opposed to two or three separate types of peptide synthetase genes whose products must work in different combinations to make lysergic acid amides (such as ergonovine and LAH) or ergopeptines in the Clavicipitaceae [11,21]. Phylogenetic analyses indicated that the ability to produce lysergic acid evolved once in an ancestor to both the Aspergillus species and the members of the Clavicipitaceae, but the ability to assemble derivatives of lysergic acid evolved independently in the two lineages [14].
Recently, a new species of Aspergillus, A. aspearensis Houbraken, Frisvad, Arzanlou & Samson, was described after being isolated from soil from Aspear Island in Urmia Lake of Iran [22]. Based on phylogenetic analyses of a concatenated dataset derived from genes encoding β-tubulin, calmodulin, and the second largest subunit of RNA polymerase II, A. aspearensis grouped as a sister species to A. leporis in a clade within section Flavi that contained A. leporis and A. hancockii as its only other members [22]. Considering the phylogenetic placement of this new species, we hypothesized that A. aspearensis would produce ergot alkaloids and tested that hypothesis by chemical and genomic approaches.
2. Results
2.1. Accumulation of Lysergic Acid Amides in Cultures of A. aspearensis
Cultures of A. aspearensis isolate CBS 143673 grown on malt extract agar for two weeks at 30 °C were analyzed for the presence of ergot alkaloids. Extracts were derived from 400-μL samples containing all fungal material present in that region of the culture dish (hyphae, conidia, and, potentially, immature sclerotia) as well as the agar medium (to the bottom of the Petri dish) on which the sampled portion of the culture had grown. Extracts of A. aspearensis contained abundant ergot alkaloids of the lysergic acid amide class—most abundantly the stereoisomers of LAH and its hydrolysis product, ergine—as assessed by comparison to extracts of A. leporis in high-performance liquid chromatography (HPLC) analyses (Figure 2). Analytes from the A. leporis extracts had been characterized previously as the specified lysergic acid derivatives by additional instrumental approaches [14].
Ten other culture media were assessed for their ability to support LAH accumulation in A. aspearensis in a survey conducted with individual cultures. Among media tested, only pyrithiamine medium [23] and acetamide medium [24] supported accumulation of LAH to concentrations similar to those obtained on malt extract agar (Table S1). All other tested media resulted in accumulation of LAH to concentrations less than 20% of that obtained on malt extract agar. Malt extract agar, as opposed to either of the other high-yielding media, was chosen for conducting all other culture-based experiments because of its low cost and its support of production of abundant conidia and sclerotia.
Quantitative analyses of ergot alkaloids and the effect of temperature on ergot alkaloid accumulation and colony diameter were investigated in cultures grown on malt extract agar for 12 days. Cultures grown at 30 °C had significantly greater colony diameter and significantly greater concentrations of LAH and other ergot alkaloids with the exception of ergonovine, which was not a significant component of the ergot alkaloid profile at any temperature (Figure 3). The increase in ergot alkaloid accumulation at 30 °C suggests that either ergot alkaloid synthesis gene expression or enzyme activity is greatest at that temperature. Our data do not allow us to discriminate between these two possibilities.
Since ergot alkaloids were detected in complex samples of A. aspearensis containing hyphae, conidia, sclerotia (in some cases), and the agar medium on which the fungus had grown, we tried to separate some of these components to better assess where ergot alkaloids were accumulating. Conidia harvested separately from the remainder of their source culture contained abundant ergot alkaloids, whereas sclerotia separated from their source culture contained notably lower yields of ergot alkaloids per g dry weight than observed in conidia (Table 1). During the preparation of conidial extracts we weighed dried samples containing enumerated conidia, allowing us to calculate the average mass of an individual conidium of A. aspearensis as 4.5 ± 1.0 pg (mean ± standard error). That mass is intermediate between the masses calculated previously for conidia of A. fumigatus (2.9 ± 0.3 pg) and A. nidulans (8.8 ± 1.8 pg) [25].
To separate culture medium from the fungus, static test tube cultures were grown in 500 μL of malt extract broth. In the static liquid cultures, 46% of the total ergot alkaloids were secreted into the culture medium with the remaining 54% associated with the solid phase (hyphae, conidia, and perhaps immature sclerotia) (Table 2). Lysergic acid and ergonovine, which were minor contributors (in terms of quantity) to the ergot alkaloid profile, appeared to be secreted at higher rates than LAH or ergine. In a statistical test, however, ergot alkaloid species only marginally affected secretion in a one-way ANOVA (p = 0.06). Overall, the data indicate that about half of the total ergot alkaloids of A. aspearensis are secreted and that the half remaining in the solid phase is distributed mainly between hyphae and conidia (with very little in sclerotia). Our experimental approach did not allow us to cleanly assign separate alkaloid concentration values to hyphae, because we could not obtain hyphal samples that were free of conidia.
2.2. Infection of Galleria mellonella Larvae by A. aspearensis Is Associated with Lethality and the Accumulation of Lysergic Acid Amides
Because the related fungus A. leporis parasitized insects and produced ergot alkaloids in insects [26], we tested the pathogenic potential of A. aspearensis in similar assays. Injection of conidia of A. aspearensis into larvae of the model lepidopteran G. mellonella resulted in death of all inoculated larvae over 3 to 4 days with an LT_50_ (median time to lethality) over three trials of 80 ± 6 hr (mean ± standard deviation) (Figure 4). The LT_50_ of A. aspearensis was similar to that observed with A. leporis (85 ± 10 hr) in previous trials conducted with similar concentrations of conidial inoculum [26]. No mortality was observed over this same time period in larvae that had been mock inoculated with a control solution of phosphate-buffered saline (PBS). The difference in survival curves for A. aspearensis-inoculated larvae and control larvae was statistically significant in a log rank test (p < 0.0001, with Bonferroni-adjusted alpha set at 0.0083).
Aspergillus aspearensis emerged from several dead larvae and conidiated and formed sclerotia on cadavers (Figure 5), indicating that the fungus can complete its asexual life cycle on an insect host. When larvae of G. mellonella were inoculated topically with A. aspearensis by placing them on sporulating cultures, only 2 of 30 larvae (among three trials) died over a subsequent 10-day observation period (Figure 4). The survival curves for larvae topically inoculated with A. aspearensis compared to non-inoculated controls did not differ significantly in a log rank test (p = 0.15, with Bonferroni-adjusted alpha set at 0.0083). These data suggest that although A. aspearensis has significant pathogenic potential, it lacks an effective mechanism to penetrate the insect’s cuticle. In the absence of such a mechanism, the fungus would need to infect through wounds or be vectored by some agent capable of cuticle penetration. Effective insect pathogens rely on appressoria and/or cuticle-degrading enzymes to penetrate an insect hosts’ cuticle [7].
Ergot alkaloids accumulated in insect cadavers consumed by A. aspearensis. The same lysergic acid amides detected in cultures of A. aspearensis (and A. leporis) were observed in infected larvae (Figure 6). The concentrations of most ergot alkaloids that accumulated in G. mellonella infected with A. aspearensis and A. leporis were similar (Figure 7). One exception was a significantly higher concentration of lysergic acid in the larvae infected with A. aspearensis. Previous studies have demonstrated the significance of LAH to insect pathogenesis by A. leporis [26] and M. brunneum [8]. Any potential contribution of lysergic acid to insect pathogenesis has not been tested and should not be inferred from these results.
2.3. Ergot Alkaloid Synthesis Genes in A. aspearensis
A draft genome assembly of Illumina technology-based reads of A. aspearensis CBS 143673 yielded a genome of 41.3 Mb over 2085 contigs, with 77-fold coverage, an N_50_ of 396 kb, and an L_50_ of 26. The genome contained two separate contigs with ergot alkaloid synthesis (eas) gene clusters that each contained all the necessary genes to encode the biosynthesis of LAH (Figure 8). One contig (GenBank accession JBIGLF010000097) contained an ergot alkaloid synthesis (eas) cluster that was similar to, and syntenic with, eas cluster 1 of A. leporis and the lone eas cluster of Aspergillus homomorphus [14]. The second contig (GenBank accession JBIGLF010000037) contained an eas cluster that was syntenic with eas cluster 2 of A. leporis [14], including a homolog of the major facilitator superfamily transporter gene, easT [27], which had until now not been identified in other eas clusters. One noteworthy difference between A. aspearensis eas cluster 2 and eas cluster 2 of A. leporis, is that the gene easD, which is a pseudogene in A. leporis eas cluster 2, has no frame shifts or premature stop codons in the second eas cluster of A. aspearensis and thus appears to encode a functioning protein.
In addition to the two LAH-associated gene clusters, the previously studied genome of A. leporis is unusual in having two satellite clusters that encode some of the genes required to make products of two different ergot alkaloid pathway branches: fumigaclavine A and rugulovasines A/B [28]. In order to be functional, these satellite clusters in A. leporis must rely on activities of some of the core eas genes encoded in the LAH-associated clusters in A. leporis. Based on the similarity of A. aspearensis to A. leporis, we searched the genome of A. aspearensis for evidence of full or partial biosynthetic gene clusters for these alternate ergot alkaloid pathway products. Blastn searches with easH or easQ from the rugulovasine satellite cluster of A. leporis identified a contig (GenBank accession JBIGLF010000052) containing homologs of easH and easQ adjacent to each other and oriented to be transcribed divergently as they are in A. leporis [28]. We obtained no evidence of accumulation of rugulovasines in A. aspearensis. Conditions under which rugulovasines are produced in A. leporis are not well defined; in multiple analyses in A. leporis only a single culture was observed to contain the stereoisomeric pair rugulovasine A and B [28]. Sequences flanking easH and easQ in A. aspearensis were explored by blastx searches but no additional ergot alkaloid synthesis genes were discovered within 20 kb adjacent on either side of the easH and easQ mini-cluster. Unlike the genome of A. leporis, no cluster, complete or partial, capable of contributing to fumigaclavine A synthesis was detected in the genome of A. aspearensis. Critical genes in this branch for which there were no reasonable homologs in A. aspearensis included reductase allele of easA, and the fumigaclavine A branch-associated genes easM and easN. There were no convincing homologs for ergot alkaloid synthesis genes in the genome of A. aspearensis outside of the clusters detailed above.
3. Discussion
The production of lysergic acid amides by A. aspearensis is a rare property among free-living, soil-inhabiting fungi. Aspergillus aspearensis shares this exceptional ability with three other soil-inhabiting Aspergillus species from different geographic regions: A. leporis (from the western USA), A. hancockii (Australia), and A. homomorphus (Israel and Yemen) [14]. Our results also add to the noteworthy association of lysergic acid amides—LAH in particular—with soil inhabiting fungi that have the potential to parasitize insects. In addition to the Aspergillus species, several Metarhizium species produce LAH (most prominently among other lysergic acid amides) and are insect-infecting, rhizosphere inhabitants [7,29]. In another example, Periglandula clandestina, though it symbiotically inhabits the shoots of morning glories, sends much of its lysergic acid amides to the roots of its host plant [30] where they may contribute to resistance to biotic stresses [31,32]. By contrast the other major class of lysergic acid derivatives, ergopeptines, has been documented mainly from plant shoot or seed inhabiting organisms such as Epichloë and Claviceps species and exert their effects on organisms that interact with those above-ground tissues [2,5,6].
The profile of ergot alkaloids observed in cultures A. aspearensis was qualitatively similar to that observed in A. leporis, though fungal cultures were grown on different media (as was necessary to get significant yields of ergot alkaloids from the different species). Cultures of A. aspearensis retained much of their ergot alkaloids in LAH, whereas A. leporis tended to have larger shares of lysergyl-alanine and ergine—hydrolysis products derived from an intermediate step and from LAH, respectively—than did A. aspearensis (Figure 2). In insects, where it was possible to directly compare quantities of ergot alkaloids of the two species on the same substrate, both species contained mainly LAH but A. aspearensis accumulated significantly more lysergic acid than did A. leporis in this context.
Our analyses of ergot alkaloid accumulation in cultures of A. aspearensis revealed that the lysergic acid amides accumulated in more than one part of the culture. Approximately half of the ergot alkaloid yield was secreted into the culture medium, and the other half was retained in the solid phase. Jones et al. [14] reported 66% of the ergot alkaloid content of A. leporis was secreted, a value similar to what we measured in A. aspearensis. Conidia of A. aspearensis contained relatively high concentrations of lysergic acid amides: 3.2 µmol/g dry weight. This concentration is on the same order of magnitude with the 8.1 µmol/g dry weight value observed for fumigaclavines—products of a different branch of the ergot alkaloid pathway—in A. fumigatus, a fungus renowned for concentrating its ergot alkaloids in its conidia [33]. Sclerotia of A. aspearensis, however, contained very little of the lysergic acid amides: 7.7 nmol/g dry weight (almost 500-fold lower per unit weight than in conidia). This observation is somewhat surprising because sclerotia are the primary site of accumulation of ergot alkaloids in ergot fungi (Claviceps species), where typical concentrations of ergot alkaloids have been documented to be approximately one-thousand-fold higher (in the 1 to 10 µmol/g range) in C. purpurea and C. africana [34,35] than in A. aspearensis sclerotia. Sclerotia are stress resisting structures and may contain alkaloids to protect them from biotic stresses. Frisvad et al. [22] noted accumulation of several other classes of specialized metabolites (an aflavinine, kojic acid, mevinolins, and paspalinines) in A. aspearensis, raising the possibility that these fungal structures may be protected by alternate classes of chemicals.
Aspergillus aspearensis demonstrated good pathogenic potential in the model insect G. mellonella, killing insects at a rate comparable to that previously established with A. leporis [26]. Aspergillus aspearensis was only an effective pathogen, however, when injected into the hemocoel, suggesting that it does not have a good mechanism to breach the insect cuticle. In previous experiments [26] insects topically inoculated with A. leporis died significantly faster than uninoculated control larvae; though injection was still a more effective inoculation method for A. leporis. The production of conidia and sclerotia of A. aspearensis on dead insects was striking and indicated that A. aspearensis can complete its life cycle on an insect. Moreover, the ergot alkaloids of A. aspearensis accumulated to very high concentrations—approximately 24 nmol per larva. Considering an approximate volume of 200 µL per larva, the measured quantity of ergot alkaloids translates to approximately 100 µM. This insect-associated concentration is approximately 25-fold higher than the concentration observed in malt extract cultures of A. aspearensis of about the same age. An increase in lysergic acid amide concentration in insects relative to cultures also has been documented in Metarhizium species [29].
The genome of A. aspearensis contained two complete biosynthetic gene clusters, each of which had the capacity to encode the biosynthesis of LAH. The presence of the lysergic acid amides predicted by the gene sequences shows that at least one of each homologous gene pair is functional; assessment of functionality of genes from both eas clusters in this context is theoretical and based on the lack of frame shifts or premature stop codons in their coding sequences. In other sequenced examples in which ergot alkaloid producers had two eas clusters, at least one of the duplicated genes was pseudogenized. These examples include pseudogenization of easD in eas cluster 2 of A. leporis [14] and pseudogenization of a copy of lpsB in one of two eas clusters in Epichloë coenophiala [36]. Notably eas cluster 1 and eas cluster 2 of A. aspearensis are not duplicates of each other in terms of gene synteny or even gene content. This point is true for the two clusters of A. leporis, as well. Thus, the two eas clusters most likely did not arise by a recent duplication event in either of the Aspergillus species. Cluster 1 is syntenic with the lone eas cluster of A. homomorphus, whereas cluster 2 has an additional gene—easT encoding a major facilitator superfamily transporter—and has two changes in gene order or gene position relative to eas cluster 1. Previously, easT was characterized functionally in A. leporis and found to contribute to ergot alkaloid synthesis (quantitatively) but not secretion [27]. The pseudogenization of easD in cluster 2 of A. leporis presumably happened after A. leporis and A. aspearensis diverged from a recent common ancestor. The lack of pseudogenization of any of the A. aspearensis eas genes may be due simply to random probability (since only one gene is pseudogenized in A. leporis) or to some unidentified selective pressure that differs between the two fungi due to their ecological niches or geography. Interestingly, A. hancockii, which also is part of the A. leporis phylogenetic clade [22], contains several pseudogenized eas genes in its genome though it maintains at least one functional copy of each essential gene and makes lysergic acid amides [14]. The genome of A. aspearensis lacked some of the complexity of ergot alkaloid synthesis genes noted previously in A. leporis [28] in that it lacked the partial, satellite gene cluster involved in making fumigaclavine A in A. leporis. Differences in specialized metabolites other than ergot alkaloids were noted in previous studies of A. aspearensis and A. leporis [22]. Aspergillus leporis, produced six classes of specialized metabolites that were not present in similarly cultivated A. aspearensis; conversely, A. aspearensis produced two other classes of specialized metabolites that were not detected in A. leporis.
Lysergic acid amides have now been documented in all three members (A. aspearensis, A. leporis, and A. hancockii) of the A. leporis clade, a small clade embedded within the large section Flavi of the highly speciose genus Aspergillus [22]. The ergot alkaloid synthesis gene clusters of these Aspergillus species are complex and diverse. Reasons why these particular species harbor these gene clusters and produce these bioactive alkaloids whereas their phylogenetic relatives do not may be a fruitful area for future phylogenetic and chemical ecology-based investigations. Most other producers of lysergic acid-based ergot alkaloids are plant-associated members of the Clavicipitaceae, many of which do not grow well or produce ergot alkaloids well away from their plant hosts. Thus, fast-growing saprotrophic ergot alkaloid producers like A. aspearensis, with their wealth of genetic resources, also may have translational significance for research and development relevant to understanding, modifying, controlling, or overexpressing this important group of agricultural and pharmaceutical chemicals.
4. Materials and Methods
4.1. Fungi and Culture Conditions
Aspergillus aspearensis Houbraken, Frisvad, Arzanlou & Samson strain CBS 143673 was obtained from The Westerdijk Fungal Biodiversity Institute, Utrecht, The Netherlands, and maintained on malt extract agar (per liter: 6 g malt extract, 1.8 g maltose, 6 g dextrose, 1.2 g yeast extract, 15 g agar [34]). Unless time or temperature were varied as part of a specific experiment (in which case, details are provided for that experiment), cultures were grown for two weeks at 30 °C in an incubator with ambient oxygen. Aspergillus aspearensis was grown on additional media (24 days at room temperature), to test for ergot alkaloid accumulation, including pyrithiamine medium [23], acetamide medium [24], bar maintenance medium [37], coconut agar [38], Rose Bengal agar [39], Czapek-Dox agar [40], Neurospora minimal medium [40], sucrose-yeast extract agar [29], Luria–Bertani agar [41], and Gamborg’s medium [42]. Aspergillus leporis States & M. Chr. strain NRRL 3216 was obtained from the USDA Agricultural Research Service Culture Collection (NRRL) at Peoria, Illinois, USA, and was cultivated as a source of reference material for lysergic acid amides. Aspergillus leporis was grown on sucrose-yeast extract medium [29], which was previously shown to promote excellent accumulation of lysergic acid amides in that fungus [14].
To collect conidia of A. aspearensis separately from hyphae and sclerotia, malt extract agar plates containing one-month-old cultures of the fungus were inverted (such that the lid was down) and tapped several times to dislodge conidia so that they would collect on the inner surface of the lid. Any sclerotia that landed among the conidia were removed with forceps. Conidia were then harvested from the inner surface of the lid by washing it with 50% methanol. An aliquot was counted on a hemacytometer after diluting in water containing 0.1% tween 20. Conidia were then collected on a pre-weighed 0.2-µm-pore nylon centrifugal filter and dried in a speed-vac before weighing. Sclerotia were manually collected separately from conidia and hyphae, dried in a speed-vac, and weighed prior to extraction of ergot alkaloids.
To compare accumulation of ergot alkaloids in culture fluids as opposed to the solid phase (hyphae, conidia, and, potentially, immature sclerotia) of the fungus, cultures were grown from 50,000 conidia in 0.5-mL of malt extract medium (lacking agar) contained in a 2-mL screw-cap tubes with the lids unscrewed one-quarter turn to provide aeration. After two weeks at 30 °C, the liquid phase of the culture was removed with a micropipette and the volume was measured. The solid phase of the culture was dried in a speed-vac and its dry weight determined. Ergot alkaloids were extracted from liquid by the addition of an equal volume of methanol and from the solid phases by the additional of 1 mL of methanol and bead beating as described immediately below.
Ergot alkaloids were extracted by bead beating cultured fungi or infected insects in methanol as previously described [8,14,26,29]. Briefly, samples were extracted in 2-mL screw-cap microcentrifuge tubes (capped with lids containing rubber O-rings) containing 10 three-mm glass beads. Samples were pulverized by bead beating at 6 m/s for 30 s in a FastPrep 101 (Bio101, Carlsbad, CA, USA) and clarified by centrifugation prior to analysis.
4.2. Inoculation of Insects
To test whether the fungi could grow and produce ergot alkaloids in live insects, conidia (60,000 conidia in 20 μL) in modified phosphate-buffered saline (PBS) [33] were injected into the hindmost proleg of larvae of the greater wax moth, Galleria mellonella, according to methods we have used routinely for several species of fungi [8,26,29,33]. Additional larvae were inoculated topically by placing them on a sporulating culture of A. aspearensis for 24 h before removing them to an empty Petri dish. Survival of insects inoculated with A. aspearensis by either method compared to those inoculated with a sterile PBS solution was monitored and recorded every few hours till all treated larvae were dead or to a maximum of 10 days. All experiments with insects were conducted at room temperature (20 °C to 22 °C). Data were collected from three trials with 10 larvae per treatment per trial.
4.3. Analysis of Ergot Alkaloids
HPLC was based on the methods described previously [8,14,29,37] on a Waters (Milford, MA, USA) Arc HPLC system with fluorescence of analytes monitored with excitation at 310 nm and emission at 410 nm. The solid phase was a Prodigy (Phenomenex, Torrance, CA, USA) C_18_ column with the following properties: length of 150 mm, inside diameter of 4.6 mm, and particle diameter of 5 μm. The mobile phase a was multilinear binary gradient combining 5% acetonitrile + 95% 50 mM ammonium acetate (solution A) and 75% acetonitrile + 25% 50 mM ammonium acetate (solution B) at a flow rate of 1 mL/min. The gradient originated at 100% solution A and ramped linearly to 85% solution A + 15% solution B at 8 min, then to 50% solution A/50% solution B at 25 min, and 100% solution B at 30 min. The flow was held at 100% solution B for 5 min before returning to 100% solution A over 5 min and rinsing with 100% solution A for an additional 5 min before the next injection. Identities of peaks containing lysergic acid, lysergyl-alanine, ergine, LAH, and ergonovine were defined by retention times relative to peaks in reference strains that had been previously defined by additional instrumental analyses [8,14,28,29,37]. Ergot alkaloids were quantified by peak areas relative to an external standard curve prepared with commercially available ergonovine (Sigma, St Louis, MO, USA) and thus values (with the exception of values for ergonovine) must be considered as relative to ergonovine as opposed to absolute.
4.4. Statistical Analysis of Results
Statistical analyses of ergot alkaloid quantities were conducted as follows. Datasets were first analyzed for unequal variances by Brown-Forsythe tests. Datasets that passed a Brown-Forsythe test (p > 0.05) were subsequently analyzed by ANOVA and, when treatment was a significant factor, means were separated in Tukey’s test. Datasets that did not pass a Brown-Forsythe test (p < 0.05) were analyzed nonparametrically by a Wilcoxon’s test and, when treatment was a significant factor, a Steel–Dwass multiple comparison test. Survival data from insect pathogenesis experiments were plotted on Kaplan–Meier survival curves and analyzed by log rank tests with a Bonferroni-adjusted alpha set at 0.0083. All statistical analyses were conducted in JMP edition 18 (SAS, Cary, NC, USA).
4.5. Genomic Sequence Acquisition and Analysis
The genome of A. aspearensis was sequenced by Illumina (San Diego, CA, USA) NextSeq technology with the assistance of the West Virginia University and Marshall University Genomics Core Facilities and assembled at the Marshall University Bioinformatics Core Facility. Raw reads were trimmed in Trimmomatic version 0.39 [43] by application of the following tools of that software package: the ILLUMINACLIP tool was used with default parameters to remove sequencing adapters from the reads, the SLIDINGWINDOW tool was used with a window size of 4 bases and a minimum quality score of 15 to remove low-quality base calls, and finally, the MINLEN tool was used to filter out reads less than 25 bases in length. Trimmed reads were assembled in SPAdes version 3.15.4 [44] with default settings. To estimate genome coverage, we mapped the trimmed reads back to the assembled genome using bbmap version 39.06 [45]. Genome sizes, N50, and L50 statistics were computed in Python version 3.10.13 by parsing the fasta files. The genome has been deposited at GenBank under accession number JBIGLF000000000.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Matossian M.K. Poisons of the Past: Molds, Epidemics, and History Yale University Press New Haven, CT, USA 1989
- 2Haarmann T. Rolke Y. Giesbart S. Tudzynski P. Ergot: From witchcraft to biotechnology Mol. Plant Pathol.20091056357710.1111/j.1364-3703.2009.00548.x 19523108 PMC 6640538 · doi ↗ · pubmed ↗
- 3Caradus J.R. Card S.D. Finch S.C. Hume D.E. Johnson L.J. Mace W.J. Popay A.J. Ergot alkaloids in New Zealand pastures and their impact N. Z. J. Agri. Res.20226514110.1080/00288233.2020.1785514 · doi ↗
- 4Klotz J.L. Activities and effects of ergot alkaloids on livestock physiology and production Toxins 201572801282110.3390/toxins 708280126226000 PMC 4549725 · doi ↗ · pubmed ↗
- 5Potter D.A. Stokes J.T. Redmond C.T. Schardl C.L. Panaccione D.G. Contribution of ergot alkaloids to suppression of a grass-feeding caterpillar assessed with gene-knockout endophytes in perennial ryegrass Entomol. Exp. Appl.2008126138147
- 6Hudson D. Mace W. Popay A. Jensen J. Mc Kenzie C. Cameron C. Johnson R. Genetic manipulation of the ergot alkaloid pathway in Epichloë festucae var. lolii and its effect on black beetle feeding deterrence Toxins 2021137610.3390/toxins 1302007633498584 PMC 7909537 · doi ↗ · pubmed ↗
- 7St Leger R.J. Wang J.B. Metarhizium: Jack of all trades, master of many Open Biol.20201020030710.1098/rsob.20030733292103 PMC 7776561 · doi ↗ · pubmed ↗
- 8Steen C.R. Sampson J.K. Panaccione D.G. A Baeyer-Villiger monooxygenase gene involved in the synthesis of lysergic acid amides affects the interaction of the fungus Metarhizium brunneum with insects Appl. Environ. Microbiol.202187 e 00748-2110.1128/AEM.00748-2134160271 PMC 8357275 · doi ↗ · pubmed ↗