Chemical Investigation of the Global Regulator veA-Overexpressed Mutant of an Arctic Strain Aspergillus sydowii MNP-2
Qing Gong, Wei Wang, Yujie Zhao, Xiaoying Wang, Xuelian Bai, Huawei Zhang

TL;DR
Overexpressing the veA gene in a cold-loving fungus activated hidden chemical pathways, leading to new and known compounds.
Contribution
Overexpression of veA in Aspergillus sydowii activated silent gene clusters, producing a novel polyhydroxy anthraquinone and expanding natural product diversity.
Findings
Overexpression of veA in Aspergillus sydowii MNP-2 led to the discovery of a novel polyhydroxy anthraquinone and nine known metabolites.
Compounds 1 and 10 were biosynthetically derived from T2PK and NRPS pathways previously silent in the wild-type strain.
Compound 3 showed weak antifungal activity against Candida albicans, and compound 4 had weak cytotoxicity against HCT116 cells.
Abstract
A growing body of evidence indicates that artificial manipulation of transcriptional regulation is a powerful approach to activate cryptic biosynthetic gene clusters (BGCs) of secondary metabolites (SMs) in fungi. In this study, one mutant strain MNP-2-OE::veA was constructed by overexpressing the global transcription regulator veA in an Arctic-derived strain Aspergillus sydowii MNP-2. Chemical investigation of the mutant OE::veA resulted in the isolation of one novel polyhydroxy anthraquinone (1) together with nine known metabolites (2–10), which were unambiguously characterized by various spectroscopic methods including 1D and 2D NMR and HR-ESI-MS as well as via comparison with literature data. Biosynthetically, compounds 1 and 10 as new arising chemicals were, respectively, formed by type II polyketide synthase (T2PK) and non-ribosomal peptide synthetase (NRPS), which were silent in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
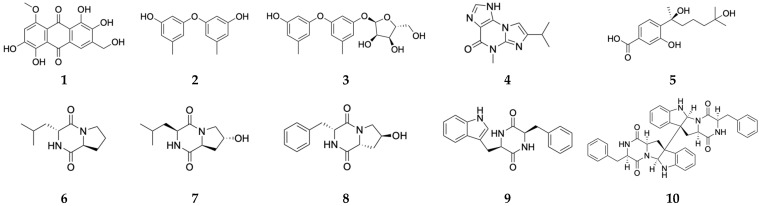 Figure 1
Figure 1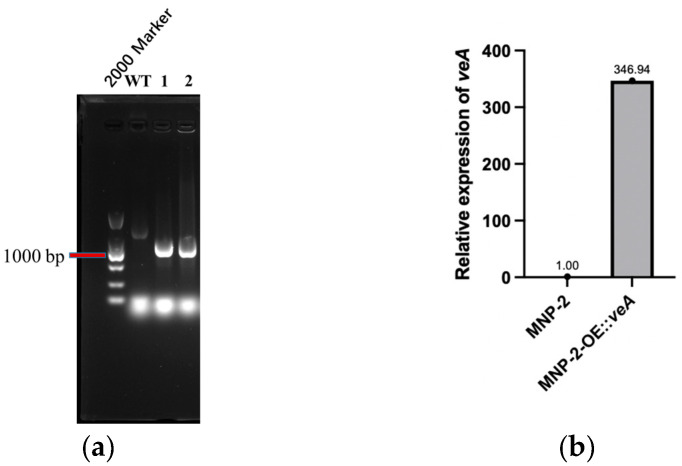 Figure 2
Figure 2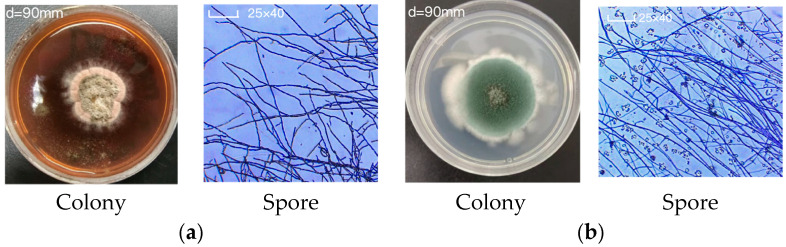 Figure 3
Figure 3 Figure 4
Figure 4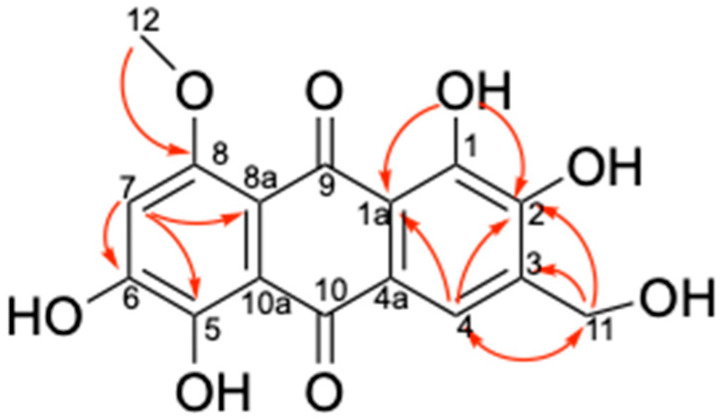 Figure 5
Figure 5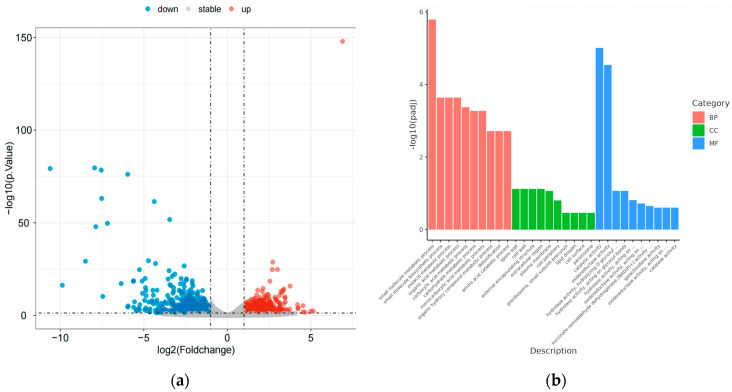 Figure 6
Figure 6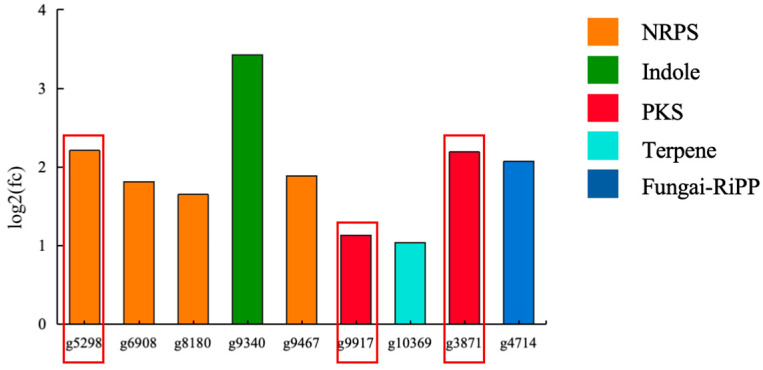 Figure 7
Figure 7- —the National Key Research and Development Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Natural Products and Biosynthesis · Fungal and yeast genetics research · Microbial Metabolic Engineering and Bioproduction
1. Introduction
Marine-derived Aspergillus fungi produce a diverse array of structurally unique and biologically active secondary metabolites (SMs), representing a hotspot in current field of natural product research [1,2,3,4,5]. A. sydowii, a ubiquitous fungus widely distributed across various ecosystems, is renowned for its extraordinary environmental adaptability and impressive secondary metabolic potential [6]. Genome sequencing and bioinformatics analyses suggest that the number of SM biosynthetic gene clusters (BGCs) in A. sydowii strains far exceeds the chemical diversity of compounds produced by them [7,8]. Therefore, the vast majority of these BGCs remain transcriptionally silent, making the discovery of novel structures challenging. In the past two decades, multiple strategies, such as epigenetic regulation, promoter engineering, global regulation, and others, have been developed and employed to activate silenced BGCs, thereby activating fungal secondary metabolic capabilities and discovering diversified SMs [9,10,11]. Of these approaches, global regulatory factor modification is considered one of the most promising strategies due to its ability to simultaneously affect multiple BGCs and its straightforward operability. veA is one of the most prevalent global regulators within the Aspergillus genus, playing a pivotal role in modulating morphogenesis and secondary metabolism [12,13,14].
In our previous studies, we performed whole-genome analysis and metabolic profiling of an Arctic-derived fungus A. sydowii MNP-2 using the global natural product social molecular network (GNPS) approach, suggesting that this strain harbors biosynthetic potential to produce diverse SMs with complex structures and significant biological activities [15]. The chemical study of the wild-type (WT) strain MNP-2 had afforded 11 phenolic bisabolane sesquiterpenes (PBSs) from its rice and potato dextrose broth (PDB) fermentation extracts [16]. To awaken the latent BGCs in this strain to produce other bioactive SMs, one global regulator veA-overexpressed mutant OE::veA was successfully constructed and shown to make ten substances (1–10, Figure 1) including one novel polyhydroxy anthraquinone (1). This work highlights the construction of the veA-overexpressed mutant OE::veA and the isolation and elucidation of its derived SMs, as well as their biological properties.
2. Results and Discussion
2.1. Construction and Morphological Characterization of the Mutant Strain
Following genome sequencing and gene annotation analysis, the global regulator veA from A. sydowii MNP-2 (1755 bp in full length) was identified and selected as the target of this study. Specific primers were employed for the amplification of the veA gene, and the overexpression plasmid pOE::veA was subsequently generated (Figures S1 and S2). The overexpression mutant OE::veA was constructed using Agrobacterium tumefaciens-mediated transformation (ATMT) [17,18]. Furthermore, the Hyg resistance gene band could be specifically amplified from the transformed strains harboring the recombinant plasmid (Figure 2a). Real-time quantitative polymerase chain reaction (RT-qPCR) was performed to determine the expression level of veA in the OE::veA strain, with β-actin serving as the internal reference. Compared with the WT MNP-2, the veA gene expression level in the OE::veA strain was significantly increased by 346.94-fold (Table 1 and Figure 2b). Collectively, these findings confirmed the successful construction of the veA-overexpressing mutant OE::veA.
The WT strain A. sydowii MNP-2 was observed to produce significantly higher levels of red pigment, whereas the veA-overexpressing mutant OE::veA exhibited a marked reduction in pigment biosynthesis. Compared to the WT strain, OE::veA displayed three distinct colony phenotypic alterations: more pigmented spores, reduced surface wrinkling, and significantly larger colony diameters (Figure 3a). At the microscopic level, the WT strain MNP-2 formed dense, complex mycelial networks characterized by fewer conidia and minimal branching at hyphal tips. In contrast, OE::veA produced an increased number of conidia and developed longer, dendritic hyphal branches (Figure 3b).
2.2. LC-MS/MS Analysis of Mutant-Derived Crude Extracts
The liquid chromatography–mass spectrometry (LC-MS) data are shown in Figure 4. When cultured in modified Martin medium, the molecular weights of SMs from the WT strain MNP-2 were primarily concentrated with in the m/z 250–500 range, whereas those produced by the mutant strain OE::veA were concentrated in the m/z 200–400 range.
2.3. Isolation and Identification of OE::veA-Derived Secondary Metabolites
A total of ten compounds (1–10) were isolated from the modified Martin medium culture extract, comprising one novel compound (1) and nine known metabolites (2–10), including a rare diphenyl ether glycoside (3). Using HR-ESIMS and ^1^H NMR as well as comparison with literature data, these compounds were, respectively, identified as follows: 1,2,5,6-tetrahydroxy-3-(hydroxymethyl)-8-methoxy-9,10-anthraquinon (1) [19], diorcinol (2) [20], diorcinol-3-O-α-d-ribofuranoside (3) [21], acremolin B (4) [22], 10-hydroxysydonic acid (5) [23], cyclo (Leu-Pro) (6) [24], cyclo (L-Phe-4-OH-L-Pro) (7) [25], cyclo (L-(4-OH)-Pro-L-Leu) (8) [26], cyclo (L-Phenyl-L-tryptophyl) (9) [27], and WIN 64821 (10) [28].
Compound 1, obtained as a deep red powder that easily dissolves in DMSO-d_6_, is slightly soluble in methanol, and insoluble in chloroform. The molecular formula of 1 was established as C_16_H_12_O_8_ based on the sodium adduct molecular ion peak m/z 355.0433 [M+Na]^+^ obtained from HR-ESIMS, indicating a degree of unsaturation of 11. The ^1^H NMR spectrum (Figure S3) of 1 exhibits a methyl signal at δH 3.90 (3H, s), a methylene signal at δH 4.58 (2H, s), two aromatic protons at δH 6.99 (1H, s) and δH 7.88 (1H, s), and five active hydrogen signals at δH 13.97 (1H, brs), δH 13.87 (1H, brs), δH 11.28 (1H, brs), δH 10.51 (1H, brs), and δH 5.37 (1H, brs) (Table 2). Further analysis of its ^13^C NMR spectrum (Figure S4) indicates that compound 1 contains two ketone carbonyl signals at δC 185.4 and 187.2, along with twelve sp^2^-hybridized carbon signals at δC 157.1, 155.5, 149.4, 149.3, 147.6, 134.5, 121.9, 117.6, 115.6, 114.9, 108.7, and 106.3 ppm, which are similar to those of asperthecin, an anthraquinone pigment obtained from Aspergillus nidulans [19]. The key HMBC correlations from H-11 (δH 4.58) to C-2 (δC 149.4), C-3 (δC 134.5), and C-4 (δC 117.6), the oxygen methyl (δH 3.90) to C-8 (δC 157.1) and from H-7 (δH 6.99) to C-5 (δC 147.6), C-6 (δC 155.5), and C-8a (δC 108.7) support the assignment of the hydroxymethyl at the C3 position and the methoxy substitution at the C8 position (Figure 5). Therefore, compound 1 was unambiguously determined as 8-methoxyl-asperthecin.
Compound 2: Light yellow oily substance; soluble in methanol and chloroform. It is commonly distributed in soil fungi (serving as defensive metabolites against pathogens). Previous studies have documented its antimicrobial activity against Gram-positive bacteria Staphylococcus aureus, Bacillus subtilis, and the yeast Candida albicans [20].
Compound 3: Colorless oily substance; soluble in methanol and chloroform. Initially isolated from marine microbes, it exhibits cytotoxicity against multiple cancer cell lines, including human tissue cell lymphoma, human prostate cancer, and human colorectal adenocarcinoma cell lines [21].
Compound 4: White crystals; soluble in methanol and chloroform. It is primarily distributed in deep-sea-derived fungi [22].
Compound 5: White amorphous powder; soluble in methanol and chloroform. First isolated from marine fungi, it shows obvious inhibitory activities against pathogenic bacteria Escherichia coli, Edwardsiella tarda, Vibrio harveyi, and Vibrio parahaemolyticus [23].
Compounds 6–8: Colorless crystals; soluble in methanol and chloroform. Originally isolated from deep-sea sediments, prior bioinformatic analysis indicated that their biosynthesis is closely associated with four genes (jatA–D), which encode core non-ribosomal peptide synthetases and acetyltransferases [24,25,26].
Compound 9: White amorphous powder; soluble in methanol and chloroform. It was previously isolated from an unidentified Penicillium sp. [27].
Compound 10: White amorphous powder; soluble in methanol, slightly soluble in chloroform. It was initially derived from Aspergillus sp. It acts as a substance P (SP)-competitive antagonist binding to the human NK1 receptor, and also possesses significant NK2 receptor antagonistic activity [28].
2.4. Transcriptomic Analysis
To investigate the role of global regulatory factor veA in the biosynthesis of natural products in the mutant OE::veA, differential expression gene (DEG) analysis was carried out using transcriptomics technology. The results suggested that the mutant OE::veA had 608 upregulated genes and 635 downregulated genes compared with the WT strain MNP-2. Volcano plot analysis further validated the statistical significance of their gene expression differences (Figure 6a). Gene ontology (GO) enrichment analysis indicated that overexpression of the veA gene significantly affected metabolic process-related pathways in the strain, suggesting that this gene may be involved in regulating natural product biosynthesis through the modulation of metabolic networks (Figure 6b).
Among these greatly upregulated (p < 0.05) genes in the mutant OE::veA, nine SM biosynthesis-related genes were identified and shown to encode two polyketide synthases (PKSs), four non-ribosomal peptide synthetases (NRPSs), one terpene, one ribosomally synthesized and post-translationally modified peptide (RiPP), and one indole (Figure 7). Biosynthetically, compounds 1 and 10, as new arising SMs from the mutant OE::veA, were presumably formed by T2PK and NRPS, respectively. By Blastp-based homology alignment analysis, two upregulated PKS genes (g3871 and g9917) had a respective similarity of 56.6% and 46.43% with mdpG, which was essential for the biosynthesis of T2PK natural product xanthones [29]. The core gene (g5298), which exhibited upregulated expression in NRPS, shared 67% sequence similarity with dtpA, a key gene within the BGC responsible for the production of TDKPs [30]. Therefore, the overexpression of the veA gene activated the PKS and NRPS genes in the mutant OE::veA and resulted in the production of compounds 1 and 10.
2.5. Biological Activity of SMs
Antimicrobial assay revealed that compound 3 showed weak activity against C. albicans (MIC = 64 μg/mL). The cytotoxicity results for compounds (1–10) indicate that compound 4 with an IC_50_ value of 44.47 μM against HCT116 cells.
3. Materials and Methods
3.1. Fungal Material and Fermentation
The fungal strain A. sydowii MNP-2 was isolated from Arctic marine sediments. In this study, an overexpression strain of the global regulator veA, designated, was constructed from the WT MNP-2. For seed culture, OE::veA was inoculated into the modified Martin medium and incubated in the dark at 28 °C with shaking at 180 rpm for 15 days. For hyphal observation, 1 μL of a spore suspension (1 × 10^7^ spores/mL) from both the WT and OE:: veA strains was aseptically placed into the gap between a PDA plate and a sterile coverslip. The plates were incubated at 28 °C in the dark for 2 days. The coverslips were then carefully removed with sterile forceps, placed onto glass slides, and stained with crystal violet. Hyphal growth and morphology were examined under an optical microscope.
3.2. Construction of Overexpression Mutant
Using the newly extracted genomic cDNA of A. sydowii MNP-2 as a template, primer pairs containing the target gene were designed for veA gene amplification. Following the PCR amplification of the target gene fragment, the veA gene was cloned downstream of the gpdA promoter in the fungal expression vector via restriction enzyme digestion and ligation, constructing the recombinant plasmid pOE::veA. The validated overexpression vector was introduced into Agrobacterium tumefaciens AGL-1 via freeze–thaw transformation. The successful transformant colonies were isolated and co-cultured with A. sydowii MNP-2. After the resistance screening, overexpressing positive transformants were obtained. RNA was extracted from positive transformants, reverse transcribed into cDNA, and validated via RT-qPCR. After ten generations of stable passage on antibiotic plates, RNA was extracted from overexpressing mutant strains and WT strains, was reverse transcribed into cDNA templates, and subjected to RT-qPCR to detect veA gene expression levels in the WT and mutant OE::veA strains. In the RT-qPCR analysis for assessing the expression level of the veA gene in the OE::veA mutant and WT MNP-2 strains, β-actin was utilized as the internal reference gene. Specifically, the ΔCt value for each sample was calculated as the difference between the cycle threshold (Ct) of the target gene (veA) and that of the internal reference gene (β-actin). Subsequently, the WT MNP-2 strain served as the control: its average ΔCt value was first determined, and the ΔΔCt value was then derived by subtracting this average ΔCt value from the ΔCt value of the OE::veA mutant. Finally, the relative overexpression level of veA in the OE::veA mutant was calculated using the 2^−ΔΔCt^method [31], which directly reflects the fold increase in veA expression in the mutant strain relative to the WT counterpart.
Using sterile pipettes, spore suspensions (spore concentration 1 × 10^7^ spores/mL) of the WT and mutant strains were inoculated onto PDA plates. Plates were incubated at 28 °C for 8 days under dark conditions, with daily growth records taken for both strains. The positive mutant strain OE::veA maintained normal growth after the tenth generation, exhibiting significant phenotypic differences compared to the MNP-2 strain. The primers used in this study are detailed in Table S1.
3.3. General Experimental Procedures
^1^H NMR and ^13^C NMR spectra were recorded at 600 and 150 MHz, respectively, using Bruker Avance DRX600 instruments (Bruker, Fällande, Switzerland). ESIMS were obtainedwith an Agilent 6210 LC/TOF-MS spectrometer (Agilent Technologies, Santa Clara, CA, USA). Reverse phase HPLC wascarried out on an Essentia LC-16P apparatus (Essentia, San Diego, CA, USA) fitted with apreparative HPLC column (Phenomenex Gemini-NX C18 column 50 mm × 21.2 mm, 5 μm; Phenomenex, Inc., Torrance, CA, USA). It operated at a flow rate of 1.0 mL/min. All the chemicals used were of analytical grade.
3.4. Liquid Chromatography–Mass Spectrometry Analysis
A small amount of fermented crude extract was dried by blowing air, and then was adjusted to a concentration of 1 mg/mL. Centrifugation was performed at 12,000 rpm and 4 °C for 20 min, and the supernatant was collected into a sample vial. The instrumentation used was a SCIEX X500B quadrupole time-of-flight (QTOF) high-resolution mass spectrometer (HRMS) coupled with a SCIEX ExionLC ultra-high-performance liquid chromatograph (UPLC; SCIEX, Framingham, MA, USA). The liquid chromatography conditions were as follows: the analytical column was a Phenomenex Kinetex C18 column (100 Å, 150 × 2.1 mm, 5 μm; Phenomenex, Inc., Torrance, CA, USA), with gradient elution performed using a methanol/water (MeOH/H_2_O) mobile phase system ranging from 10% to 100% MeOH at a constant flow rate of 0.3 mL/min. Mass spectrometry (MS) conditions were configured as follows: electrospray ionization (ESI) was employed as the ion source, with full-scan mode performed for both positive and negative ion modes. The mass-to-charge ratio (m/z) range was set at 100–1500 for both full MS scan and MS/MS fragmentation analysis.
3.5. Extraction and Isolation
About 10.9 g of crude extract obtained from the fermentation broth of mutant strain OE::veA cultivated in modified Martin medium using ethyl acetate as extraction solvent was subjected to semi-preparative HPLC (Gemini NX-C18 110Å, 50 × 21.2 mm, 5 μm; Phenomenex, Inc., Torrance, CA, USA) using an acetonitrile–water gradient elution (30:70 to 100:0, v/v), yielding seven fractions (Fr.1–Fr.7). Fr.3 was further purified by gel column chromatography with a mobile phase of methanol/dichloromethane (1:1, v/v), affording four subfractions (Fr.3.1–Fr.3.4). Fr.3.2 was subsequently purified by semi-preparative HPLC (Gemini C18 110Å, 250 × 4.6 mm, 5 μm; Phenomenex, Inc., Torrance, CA, USA) using acetonitrile/water (15:85, v/v) as the mobile phase, yielding compounds 6 (5.4 mg, t_R_ = 23.0 min), 7 (8.0 mg, t_R_ = 11.2 min), and 8 (7.8 mg, t_R_ = 17.9 min). Fr.4 was subjected to semi-preparative HPLC (Gemini NX-C18 110Å, 250 × 21.2 mm, 5 μm; Phenomenex, Inc., Torrance, CA, USA) using an acetonitrile–water gradient elution (20:80 to 100:0, v/v), yielding seven subfractions (Fr.4.1–Fr.4.7). A red solid precipitated upon the vacuum concentration of Fr.4.2, which was isolated as compound 1 (11.2 mg) after filtration. Fr.4.3 was further purified via semi-preparative HPLC (Gemini C18 110Å, 250 × 4.6 mm, 5 μm; Phenomenex, Inc., Torrance, CA, USA) using acetonitrile/0.1% aqueous formic acid (30:70, v/v) as the mobile phase, producing compound 5 (7.2 mg, t_R_ = 9.7 min). Fr.4.4 was purified by semi-preparative HPLC (Gemini C18 110Å, 250 × 4.6 mm, 5 μm; Phenomenex, Inc., Torrance, CA, USA) to yield compounds 2 (7.9 mg, t_R_ = 17.5 min, acetonitrile/water, 42:58, v/v) and 10 (19.1 mg, t_R_ = 14.0 min, acetonitrile/0.1% aqueous formic acid, 45:55, v/v). Fr.4.5 was further subjected to purification using semi-preparative HPLC (Gemini C18 110Å, 250 × 4.6 mm, 5 μm; Phenomenex, Inc., Torrance, CA, USA) with a mobile phase of acetonitrile/water, yielding compounds 3 (3.1 mg, t_R_ = 23.0 min, 28:82, v/v), 4 (5.8 mg, t_R_ = 16.0 min, acetonitrile/water, 18:82, v/v), and 9 (7.4 mg, t_R_ = 43.2 min, acetonitrile/water, 22:78, v/v).
3.6. Transcriptomic Analysis
For each sample, three sets of biological replicates were prepared. Library construction and Illumina sequencing were performed at Shanghai Personal Biotechnology Co. Ltd. (Hangzhou, China). The genes of differential expression were analyzed by DESeq (v1.38.3) with screen conditions as follows: expression difference multiple |log2FoldChange| > 1, significant p-value < 0.05. TopGO (v2.50.0) was used to perform GO enrichment analysis on the differentially expressed genes (all DEGs/up DEGs/down DEGs), to calculate the p-value using the hypergeometric distribution method (the standard of significant enrichment is p-value < 0.05), and to identify the GO terms with significantly enriched differential genes to determine the main biological functions performed by these differential genes. Sequence similarity was analyzed using the Blastp algorithm against the NCBI Non-Redundant Protein Database with an E-value cutoff set at 1 × 10^−5^.
3.7. Bioactivity Assay
According to the gradient dilution method [32], the antimicrobial activity of compounds 1–10 was tested against three indicator strains, including Staphylococcus aureus (S. aureus ATCC 25923), Escherichia coli (E. coli ATCC 25922), and Candida albicans (C. albicans ATCC 10231). Three pathogenic strains were obtained from Nanjing Medical University (Nanjing, China). A crude extract solution at a concentration of 10 mg/mL was prepared using DMSO. Ampicillin sodium served as the positive control for E. coli ATCC 25922 and S. aureus ATCC 25923, while amphotericin B was used as the positive control for C. albicans ATCC 10231. DMSO was used as the negative control.
The human breast cancer cell line MCF-7, the human hepatocellular carcinoma cell line HepG2, and the human colorectal cancer cell line HCT116 were purchased from the China Center for Type Culture Collection (CCTCC, Wuhan, China). The cytotoxicity of all chemicals 1–10 against MCF-7, HepG2, and HCT116 tumor cells was evaluated using the CCK-8 assay [33]. MCF-7 and HepG2 cells were cultured in DMEM medium, while HCT116 cells were maintained in RPMI 1640 medium (supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin). Cells were seeded into 96-well plates at a density of 5 × 10^3^ cells/well (100 μL/well) and incubated at 37 °C with 5% CO_2_ for 24 h. Different concentrations of compounds 1–10 were then added, followed by a further incubation for 48 h. After the 48 h incubation, a medium containing 10% CCK-8 was added to each well, and the plates were incubated for an additional 40 min. The optical density (OD) values were measured at a wavelength of 450 nm using a microplate reader.
4. Conclusions
One veA-overexpressing mutant MNP-2-OE::veA was successfully constructed and shown to make one novel polyhydroxy anthraquinone (1) together with nine known metabolites (2–10), which compounds 1 and 10, respectively, biosynthesized by T2PK and NRPS, were newly emerging substances. These results demonstrated the crucial role of veA as a global transcription regulatory factor in activating silent BGCs in fungi, providing experimental evidence and theoretical support for enhancing chemical diversity through target gene regulation. In future, other transcription regulatory factors (such as LaeA and VelB) in the genome of WT strain MNP-2 will be further overexpressed or knocked out for discovery of novel SMs with therapeutic potential.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Song Z. Gao J. Hu J. He H. Huang P. Zhang L. Song F. One new xanthenone from the marine-derived fungus Aspergillus versicolor MF 160003 Nat. Prod. Res.2020342907291210.1080/14786419.2019.159735531009246 · doi ↗ · pubmed ↗
- 2You M. Liao L. Hong S. Park W. Kwon D. Lee J. Noh M. Oh D. Oh K. Shin J. Lumazine peptides from the marine-derived fungus Aspergillus terreus Mar. Drugs 2015131290130310.3390/md 1303129025775424 PMC 4377984 · doi ↗ · pubmed ↗
- 3Hwang J. Lee J. Park S. Lee J. Oh D. Oh K. Shin J. New peptides from the marine-derived fungi Aspergillus allahabadii and Aspergillus ochraceopetaliformis Mar. Drugs 20191748810.3390/md 1709048831438635 PMC 6780696 · doi ↗ · pubmed ↗
- 4Han Y. Zhang Q. Xu W. Hai Y. Chao R. Wang C. Hou X. Wei M. Gu Y. Wang C. Targeted isolation of antitubercular cycloheptapeptides and an unusual pyrroloindoline-containing new analog, asperpyrroindotide A, using LC-MS/MS-based molecular networking Mar. Life Sci. Technol.20235859310.1007/s 42995-022-00157-836713278 PMC 9854410 · doi ↗ · pubmed ↗
- 5Andersen M. Nielsen J. Klitgaard A. Petersen L. Zachariasen M. Hansen T. Blicher L. Gotfredsen C. Larsen T. Nielsen K. Accurate prediction of secondary metabolite gene clusters in filamentous fungi Proc. Natl. Acad. Sci. USA 2013110 E 99E 10710.1073/pnas.120553211023248299 PMC 3538241 · doi ↗ · pubmed ↗
- 6Paulus C. Rebets Y. Tokovenko B. Nadmid S. Terekhova L.P. Myronovskyi M. Zotchev S.B. Rückert C. Braig S. Zahler S. New natural products identified by combined genomics-metabolomics profiling of marine Streptomyces sp. MP 131-18Sci. Rep.201774238210.1038/srep 4238228186197 PMC 5301196 · doi ↗ · pubmed ↗
- 7Bergmann S. Schümann J. Scherlach K. Lange C. Brakhage A. Hertweck C. Genomics-driven discovery of PKS-NRPS hybrid metabolites from Aspergillus nidulans Nat. Chem. Biol.2007321321710.1038/nchembio 86917369821 · doi ↗ · pubmed ↗
- 8Hou X. Liu L. Xu D. Lai D. Zhou L. Involvement of Lae A and Velvet proteins in regulating the production of mycotoxins and other fungal secondary metabolites J. Fungi 20241056110.3390/jof 10080561 PMC 1135536839194887 · doi ↗ · pubmed ↗