Characterization of Members of the Fusarium incarnatum–equiseti Species Complex from Natural and Cultivated Grasses Intended for Grazing Cattle in Argentina
María Julia Nichea, Eugenia Cendoya, Vanessa Gimena Zachetti, Luisina Delma Demonte, María Rosa Repetti, Sofia Alejandra Palacios, María Laura Ramirez

TL;DR
This study identifies diverse Fusarium species in Argentine grasses that can produce a mycotoxin linked to cattle contamination.
Contribution
The study reveals high genetic diversity within the FIESC and confirms their ZEA production potential in pasture grasses.
Findings
Phylogenetic analysis identified five known FIESC species and six putative new species.
76% of isolates produced zearalenone (ZEA), including in high quantities.
FIESC species in Argentine grasses are likely sources of ZEA contamination in grazing cattle.
Abstract
The detection of zeranol in grazing cattle could be explained by the metabolization of the mycotoxin, zearalenone (ZEA), which was proven to be naturally contaminating the grasses harboring the Fusarium species. Previous studies have suggested that members of the Fusarium incarnatum–equiseti species complex (FIESC) could be responsible for this contamination. Therefore, the objective of this study is to determine the species composition of FIESC isolates isolated from natural and cultivated pastures previously intended for livestock feed in Argentina and to analyze their ability to produce ZEA. Twenty-five Fusarium isolates were characterized by a phylogenetic analysis of the translation elongation factor 1α, and their ZEA production was quantified by cultivation in rice and subsequent analysis by UPLC-MS/MS. The phylogenetic analysis revealed a high genetic diversity identifying five…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
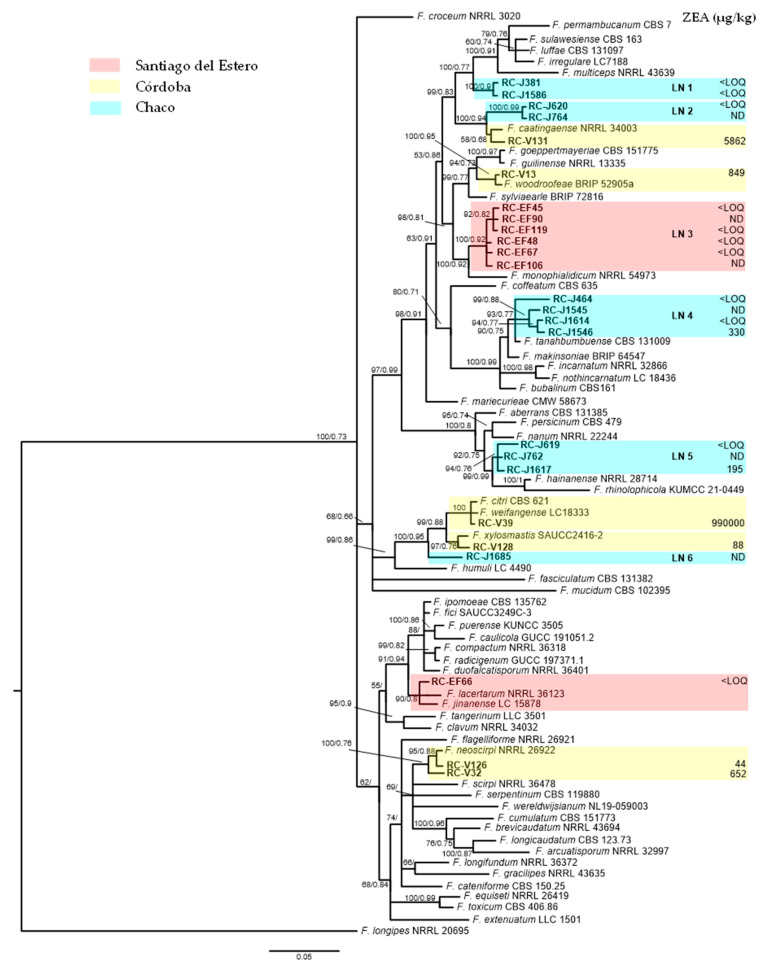 Figure 1
Figure 1- —Agencia Nacional de Promoción Científica y Tecnológica (MINCyT)
- —Consejo Nacional de Investigaciones Científicas y Técnicas
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycotoxins in Agriculture and Food · Plant and fungal interactions · Marine Toxins and Detection Methods
1. Introduction
Livestock activity is one of the main pillars of the Argentine economy, concentrated mainly in the Pampeana region, followed by the Northeast, Patagonia, the Northwest, and Cuyo. These regions have temperate-to-subtropical climates with rain regimes that allow for the development of grasslands, pastures, and greens that represent the nutritional support of livestock. Extensive beef production in Argentina is mostly based on grazing native and cultivated pastures. Native pastures rely on natural, non-cultivated plant species, offering cost savings, high biodiversity, and soil health benefits, but generally with lower yields and nutrient quality. By contrast, cultivated pastures use introduced species for high productivity and improved feed quality but demand ongoing investment, can reduce biodiversity, and have a limited lifespan [1].
Despite the obvious nutritional value of pastures, they can also pose health risks to grazing animals under certain circumstances, such as kikuyu poisoning in cattle [2,3,4] or when the growth of mycotoxigenic fungi leads to mycotoxin build-up in pastures [5,6,7].
Of particular concern to the Argentinean livestock industry is that the mycotoxin, zearalenone (ZEA), is chemically similar to the growth-promoting α-zearalanol (zeranol), which is banned in Argentina and in the European Union (EU). For the past 20 years, zeranol has been detected in bovine urine during routine analysis of beef cattle farms (enrolled as EU exporters) as part of a national residue control plan by the National Service for Health and Food Quality (SENASA), the central governing authority in Argentina. The cattle were raised on those cattle farms through natural grass grazing, without any external inputs. There is evidence of the presence of zeranol in the urine of deer, goats, sheep, cattle, and horses that only received grass-based feeding [8,9,10]. In Australia and New Zealand, the detection of zeranol in cattle could come from the ZEA metabolism, which is a mycotoxin synthesized by fungi belonging to the genus Fusarium and is found naturally in pastures intended for bovine feeding [11,12]. Kennedy et al. [13] and Smith and Morris [14] demonstrated that zeranol (α-ZAL) can be formed from Fusarium metabolites in vivo in bovine rumen. There is, then, a natural source of zeranol in the pastures, so the finding of zeranol in an animal’s urine alone is not sufficient proof that the producer has abused this substance. In 2011 and 2014, we sampled natural grasses belonging to the family Poaceae from two cattle farms in the Chaco Wetlands, where zeranol had been detected in bovine urine. We found that 90% of the samples were contaminated with ZEA at concentrations ranging from 0.7 to 2120 μg/kg d.m. (mean = 84.5 μg/kg). In addition to α-zearlanol and β-zearalenol (derived from ZEA), other metabolites produced by Fusarium, such as toxins T-2 and HT-2, beauvericin (BEA), equisetin, aurofusarin (AUF), neosolaniol (NEO), and diacetoxyscirpenol (DAS), were also detected. The mycological study showed that all samples had Fusarium contamination (60–100%), with the most frequently found species being a novel one within the Fusarium sambucinum species complex (FSASC), F. chaquense. This species is characterized by producing T-2 toxin, T-2 triol, T-2 tetraol, toxin HT-2, DAS, NEO, AUF, and BEA, but not ZEA [15]. The second most frequent isolated species belongs to the Fusarium incarnatum–equiseti species complex (FIESC), and we hypothesized that members of this complex could be responsible for the ZEA present in the grasses.
In recently published systematic reviews on the worldwide occurrence levels of mycotoxins in pastures (cultivated and natural), the data presented indicate that mycotoxins produced by the genus Fusarium were the most frequent ones, being reported in all articles evaluated. ZEA was the most prevalent mycotoxin, followed by trichothecenes [16,17].
We have also carried out other studies in natural and cultivated grasses intended for grazing cattle, searching for Fusarium endophytes [18]. These surveys have revealed that the most commonly isolated species were included in the FIESC. However, some strains in the complex could not be satisfactorily identified to the species level using morphological markers. In order to identify them, molecular biological techniques were used through phylogenetic analysis. Thus, those species are known as “phylospecies”.
Multilocus phylogenetic analyses revealed that the FIESC had more than 40 phylogenetically distinct species which had been divided into the Equiseti clade and the Incarnatum clade [19,20]. They are distributed throughout tropical, subtropical, and temperate regions. Several studies have revealed that members of the FIESC are capable of producing mycotoxins. These fungi can synthesize a range of toxic secondary metabolites, such as nivalenol (NIV) and its acetylated derivative (4-acetyl- nivalenol; 4-ANIV), deoxynivalenol (DON) and its acetylated derivatives (15-acetyldeoxynivalenol and 3-acetyldeoxynivalenol, 15-ADON and 3-ADON), DAS, NEO, ZEA, BEA, and other emerging toxins [21]. Therefore, the aims of this work were to understand the species composition of the endophyte FIESC isolates previously obtained from natural and cultivated grasses used for cattle production in Argentina and to analyze the capability of these isolates to produce ZEA.
2. Materials and Methods
2.1. Origin of Isolates Examined
Twenty-five Fusarium isolates were recovered from the culture collection of the Laboratory of Mycology, Department of Microbiology and Immunology, Universidad Nacional de Rio Cuarto (RC). The isolates were previously isolated as endophytes from the aerial part of natural and cultivated asymptomatic grasses (Poaceae) devoted to cattle grazing obtained from beef cattle farms in Argentina. In the particular case of natural grasses, the selection criteria used was to sample the most palatable ones (Table 1). All of these isolates were putatively assigned to the FIESC based on their similarity to the colony color and micromorphological characteristics described by Leslie and Summerell [22] for F. equiseti and F. semitectum, which represent the diagnostic morphotypes for members of this species complex.
2.2. DNA Extraction, Amplification and Sequencing
Total genomic DNA was extracted using the cetyltrimethylammonium bromide (CTAB; Sigma-Aldrich, St. Louis, MO, USA) method, as described by Leslie and Summerell [22]. In brief, all the isolates were grown in 50 mL of complete medium (CM) and incubated in an orbital shaker (150 rpm) for 3 d at 25 °C. The resulting mycelia were harvested by filtration through non-gauze milk filters (KenAG, Ashland, OH, USA). Excess water was removed by blotting the mycelia between clean paper towels, and the dried mycelia were stored frozen at −20 °C until ground and extracted with CTAB. The quality of the genomic DNA was determined by electrophoresis and quantified using a spectrophotometer (model ND-1000; NanoDrop Technologies, Wilmington, DE, USA).
Partial amplification of the translation elongation factor 1-α (TEF1) gene was carried out with PCR primers EF1 and EF2 using the amplification conditions described by O’Donnell et al. [23]. The PCR products were purified and sequenced by Macrogen, Inc. (Seoul, Republic of Korea), using the same primers used for the PCR amplification. Sequences were edited using the BioEdit Sequence Alignment Editor 7.1.3.0 [24]. Initial identifications of all Fusarium isolates relied on BLAST (https://blast.ncbi.nlm.nih.gov, accessed on 30 July 2025) search comparisons against the Fusarium MLST database (https://fusarium.mycobank.org, accessed on 5 August 2025). These results were then used to produce a reference dataset for the FIESC, using previously deposited sequences obtained from the NCBI nucleotide database. All sequences generated in this study were deposited in GenBank, with the accession numbers provided in Table 1.
2.3. Sequence Analysis and Phylogenetic Analysis
Multiple sequence alignment of the TEF1 gene was performed using the Web-based program MAFFT (https://mafft.cbrc.jp/alignment/server/, accessed on 5 August 2025) [25]. TEF1 sequences from the reference FFSC strains and other Fusarium species obtained from GenBank were included in the analysis (Table 2). Based on this alignment, phylogenetic analyses were performed to selected strains by maximum likelihood (ML), using PhyML 3.1 [26], and Bayesian inference (BI), using MrBayes 3.2.6 [27]. For ML and BI analyses, the best substitution model was determined using jModelTest 2.1.10 [28] and scored following the Akaike information criterion (AIC). The TrN + G model was used. For the ML analysis, the robustness of the best tree was evaluated by 1000 bootstrap replications. For the BI analysis, two runs with four chains each were run for 10 million generations with a sampling frequency of every 100 generations. Trees after the initial 25% of trees for each run were discarded as burn-in. Fusarium longipes NRRL 20695 was used as outgroup.
2.4. Zearalenone Production
All the isolates were cultured in Erlenmeyer flasks (250 mL) containing 25 g of long grain rice. Ten ml of distilled water was added before autoclaving for 30 min at 121 °C in order to reach 40% humidity. The procedure was repeated twice. Each flask was inoculated with a 3-mm diameter agar disk taken from the margin of a colony grown on synthetic nutrient agar (SNA) at 25 °C for 7 days [22]. Flasks were shaken by hand once a day for 1 week. These cultures were incubated for 28 days at 25 °C in dark. Non-inoculated flasks with rice were used as a negative control. At the end of the incubation period, the contents of the flask were dried at 50 °C for 24 h, ground to a fine powder, and then stored at −20 °C until analyzed for mycotoxins.
2.5. Zearalenone Analysis
QuEChERS extraction procedure was followed according to Lacina et al. [29] with the modifications made by Dzuman et al. [30]. The methodology was applied as follows: a 3 g portion of the homogenized sample was weighed in a polypropylene centrifugation tube and acidified Milli-Q water (10 mL, 0.2% formic acid) was added and left to soak the matrix for 30 min. Then, acetonitrile was added (10 mL), and the sample was extracted for 30 min using a laboratory shaker (New Brunswick Scientific, Edison, NJ, USA). Next, 4 g of magnesium sulfate and 1 g of sodium chloride were added and the tube was shaken for 1 min, followed by centrifugation (5 min, 10,000 rpm; Thermo Fisher Scientific Inc., Waltham, MA, USA). The organic upper layer (2 mL) was removed and shaken with 0.1 g of Bondesil-C_18_ and 0.3 g of magnesium sulfate for 2 min, followed by centrifugation (5 min, 10,000 rpm). Finally, 1 mL of purified extract was removed into a vial prior to injection on an LC–MS system.
The detection and quantitation of ZEA was performed using an ACQUITY UPLCTM system (Waters Corporation, Milford, MA, USA) coupled with a triple quadrupole mass spectrometer (MS/MS) equipped with a Z-spray electrospray ionization source (ESI). Chromatographic separation was achieved on a BEH C18 reversed-phase column (1.7 μm, 2.1 × 100 mm) maintained at 40 °C, utilizing a gradient elution with mobile phases comprising ultrapure water with 5 mM NH_4_F and 0.1% formic acid (A) and methanol with similar additives (B). The flow rate was set to 0.4 mL/min, and the injection volume was 10 μL. The MS/MS detection was carried out in positive mode, monitoring transitions m/z 319.1 → 187.0 (quantifier) and 319.1 → 185.0 (qualifier), with collision energies optimized accordingly. Quantification was performed using matrix-matched calibration curves over a concentration range from 1 to 300 ng/mL, following validation parameters including recovery, repeatability, linearity (R^2^ > 0.99) (Figure S1), and limits of detection and quantification at signal-to-noise ratios of 3 and 10, respectively. The method demonstrated the limits of detection and quantification of 3.0 ng/g and 15 ng/g, respectively, and showed high precision, with mean recoveries ranging from 85% to 98%. The entire procedure ensures the accurate, sensitive, and reproducible determination of ZEA in the analyzed samples.
3. Results
3.1. Phylogenetic Analyses
A phylogenetic analysis based on the TEF1 gene showed that the 25 isolates belonged to the FIESC (Figure 1). Part of the isolates (seven) were identified as the F. caatingaense (RC-V131), F. woodroofeae (RC-V13), F. weifangense (RC-V39), F. xylosmatis (RC-V128), F. neoscirpi (RC-V126 and RC-V32), and F. lacertarum (RC-EF66) phylospecies. Interestingly, we also found that most of the isolates could not be resolved within any phylospecies and form five monophyletic groups, provisionally called LN1 to LN5. The group LN1 (two isolates) clustered closer to the phylospecies F. multiceps, and LN2 (two isolates) shared a clade with the sister species F. caatingaense. The largest group, LN3 (six isolates), clustered closer to F. monophialidicum; LN4 (four isolates) was closely related to F. tanahbumbuense. While LN5 (three isolates) was closely related to F. hainanense; one isolate, RC-J1617, formed a singleton lineage (LN6) (Figure 1).
3.2. ZEA Production
In general, 76% (19/25) of the endophyte isolates belonging to the FIESC isolated from natural and cultivated grasses were able to produce ZEA. The highest level was shown by the F. weifangense isolate RC-V39 (990,000 µg/kg) (Figure S2). Other isolates that also produce large amounts of ZEA included F. caatingaense RC-V131 (5862 µg/kg), F. woodroofeae RC-V13 (849 µg/kg) (Figure S3), both F. neoscirpi RC-V32 and RC-V126, 652 µg/kg and 44 µg/kg, respectively (Figure S4), and F. xylosmatis RC-V128 (88 µg/kg). Most of the remaining isolates included in the novel lineages were able to produce low levels of ZEA (<LOQ > LOD), except for two isolates, RC-J1546 (LN4), which produced 330 µg/kg, and RC-J1617 (LN5), which produced 195 µg/kg (Figure 1).
4. Discussion
This is the first study to elucidate the phylogeny and the potential of ZEA production for 25 endophyte isolates belonging to the FIESC obtained from natural and cultivated grasses intended for grazing cattle in Argentina. The phylogeny suggested the occurrence of five known species in the FIESC (F. caatingaense, F. woodroofeae, F. weifangense, F. xylosmatis, F. neoscirpi, and F. lacertarum) and six lineages (one singleton, the rest with several isolates) representing putatively novel phylogenetic species. Finding several Fusarium species that may represent new taxa requires further confirmation using multilocus phylogenetic analyses combined with morphology. The genetic diversity found in our survey of the FIESC isolates from natural and cultivated grasses was high and confirmed the previous reports on the FIESC that showed high biological variability existing within this species complex isolated from multiple hosts. Members of the FIESC have been frequently associated with native and wild grasses [31,32,33,34,35,36,37] and commonly isolated from wheat, maize, rice, soybean, barley, and oat [38,39,40,41,42]. FIESC species can produce a dozen mycotoxins, mainly trichothecenes and ZEA [21,38,43].
It was noticeable that each novel lineage contained isolates obtained in the same geographical area. For example, LN3 has six members, all isolated from cultivated grass, Megathyrsus maximus (syn. Panicum maximum), from Santiago del Estero province. Additionally, the results showed that natural grasses (isolates from Córdoba and Chaco provinces) exhibit a greater diversity compared to the cultivated grass M. maximus.
In the case of the isolates from natural grasses in the Chaco province, all of them were clustered in five novel lineages (one singleton). These isolates were obtained in a previous study that we conducted in the east of Chaco Province in Argentina, in a wetland ecosystem formed by the Paraná and Paraguay river floodplain that is one of the three most biodiverse biomes in the country. As part of this study, we sampled asymptomatic Poaceae plants (n = 175) representative of 12 grass genera. All the grasses were naturally contaminated with Fusarium mycotoxins, including, mainly, ZEA and the type A trichothecenes, such as T-2 and HT-2 [12]. All the metabolites noted above are reported to be produced by various Fusarium species [21]. The mycological analysis revealed that 60–100% of the sampled plants were contaminated with Fusarium, the most prevalent one being a novel species, F. chaquense, that belongs to FSAMSC. This species was responsible for type A trichothecene grass contamination, since all the isolates were able to produce in vitro this kind of mycotoxin, and we were also able to demonstrate the presence of the biosynthetic genes [15]. The second most frequently isolated species belonged to the FIESC, which is part of the present study. We have demonstrated that 67% of these isolates (8/12) were capable of in vitro ZEA production, and we believe that they are responsible for grass contamination with this mycotoxin.
It is noteworthy that three of the known species found, such as F. weifangense, F. xylosmatis, and F. woodroofeae, have recently been described, and almost all the recorded strains are from China [20,42,44]. In particular, F. weifangense was first isolated from symptomatic tissues of Triticum aestivum [40], has recently been reported from necrotic spots of Prunus salicina [20], and causes fruit rot diseases in the Indian jujube in Thailand [45], always as a pathogen, never as a grass endophyte. Fusarium xylosmatis, which has also been recently described from necrotic spots of Xylosma congesta in China, is closely related to F. weifangense [20]. Neither of these three species has been reported as ZEA producers before. During the present study, all the isolates belonging to these species were able to produce high levels of ZEA, with F. weifangense RC-V39 being the highest producer (990,000 µg/kg).
Megathyrsus maximus is a Poaceae plant originally from Africa and is widely used as forage grass in Brazil and Argentina. There is only one previous report on endophytic Fusarium on M. maximum seeds collected in several geographic locations in Brazil. The two species found were included in the FIESC: F. hainanense and F. duofalcatisporum [34]. In the present study, we did not obtain these species; most of the Fusarium endophytes isolated from M. maximus clustered in the LN3, and one was identified as F. lacertarum. This species has previously been associated with Fusarium head blight on sorghum in the USA [46] and causing wilt in Vigna unguiculata in Brazil [47], and has also been reported in this country as a pathogen in Nopalea cochenillifera [48]. However, mycotoxin characterization in F. lacertarum has not been performed. Our strain (RC-EF66) was able to produce ZEA at very low concentrations.
We observed one isolate (RC-V13) that clustered with the known FIESC phylospecies, F. caatingaense, formally described in 2019 in Brazil, associated with Dactylopius opuntiae (Hemiptera: Dactylopiidae) [49], and also reported in cultivated rice [50]. F. caatingaense strains from rice and insects have been reported as producers of BEA, FUS, AcDON, DON, T-2, DAS, NIV, and ZEA [39,50]. In the present study, the F. caatingaense strain was also capable of producing ZEA at a high level.
Two isolates clustered with F. neoscirpi (RC-V32 and RC-V126), and both isolates were capable of ZEA production. This species was formally described by Xia et al. [51] as a unique single strain from soil.
5. Conclusions
The results of the present study enhance our understanding of FIESC presence as an endophyte in natural and cultivated grasses used for grazing cattle and also the ZEA production by the different species found. It is noticeable in the presence of six putative novel lineages. This survey shows that naturally occurring and cultivated grasses not only harbor a high diversity of known species within the FIESC, which are pathogens of rice, wheat, and other hosts, but novel Fusarium species, which have the capability to produce mycotoxins.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arelovich H.M. Bravo R.D. Martinez M.F. Development, characteristic, and trends for beef cattle production in Argentina Anim. Front.20111374510.2527/af.2011-0021 · doi ↗
- 2Bourke C.A. A review of kikuyu grass (Pennisetum clandestinum) poisoning in cattle Aust. Vet. J.20078526126710.1111/j.1751-0813.2007.00168.x 17615037 · doi ↗ · pubmed ↗
- 3Ryley M. Bourke C. Liew E.C.Y. Summerell B. Is Fusarium torulosum the causal agent of kikuyu poisoning in Australia? Australas Plant Dis. Notes 2007213313510.1071/DN 07053 · doi ↗
- 4Botha C.J. Truter M. Jacobs A. Fusarium species isolated from Pennisetum clandestinum collected during outbreaks of kikuyu poisoning in cattle in South Africa Onderstepoort J. Vet. Res.201481 e 1e 810.4102/ojvr.v 81i 1.80325686222 · doi ↗ · pubmed ↗
- 5Golinski P. Kostechi M. Golinska B.T. Golinsky P.K. Accumulation of mycotoxins in forage for winter pasture during prolonged utilisation of sward Pol. J. Vet. Sci.20036818812817777 · pubmed ↗
- 6Driehuis F. Spanjer M.C. Scholten J.M. Te Giffel M.C. Occurrence of mycotoxins in maize, grass and wheat silage for dairy cattle in the Netherlands Food Addit. Contam. Part B Surveill.20081415010.1080/1939321080223692724784536 · doi ↗ · pubmed ↗
- 7Skládanka J. Nedělník J. Adam V. Doležal P. MoravcováH. Dohnal V. Forage as a primary source of mycotoxins in animal diets Int. J. Environ. Res. Public Health 20118375010.3390/ijerph 801003721318013 PMC 3037059 · doi ↗ · pubmed ↗
- 8Erasmuson A.F. Scahill B.G. West D.M. Natural zeranol (α-zearalanol) in the urine of pasture-fed animals J. Agric. Food Chem.1984422721272510.1021/jf 00048 a 014 · doi ↗