Three New Species and Three New Records of Arthoniaceae (Ascomycota, Arthoniales) from China
Chengyue Hao, Shuhua Jiang, Zefeng Jia

TL;DR
This study discovers three new lichen species and reports three new records in China, using both physical and genetic analysis.
Contribution
The paper introduces three new lichen species and provides the first molecular data for the genus Eremothecella.
Findings
Three new species of lichenized fungi were identified in Yunnan, China.
Three species are reported as new records for China.
Molecular data for the genus Eremothecella is presented for the first time.
Abstract
Based on morphological and phylogenetic analyses, during a study on the biodiversity of the lichenized fungi family Arthoniaceae in Yunnan, China, three new species of Arthonia, Eremothecella, and Synarthonia (Arthonia yunnanensis sp. nov., A. pararubella sp. nov., Eremothecella pruinocarpa sp. nov.) and three new Chinese records (A. rubella, E. calamicola, Synarthonia inconspicua) were reported in this present study. A phylogenetic analysis using maximum likelihood and Bayesian inference based on a two-gene dataset (mtSSU and RPB2) revealed that all these species form distinct independent clades. In addition, this study represents the first report of molecular sequences for the genus Eremothecella. Detailed descriptions, ecological and chemical characteristics, and illustrations for each species are provided.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
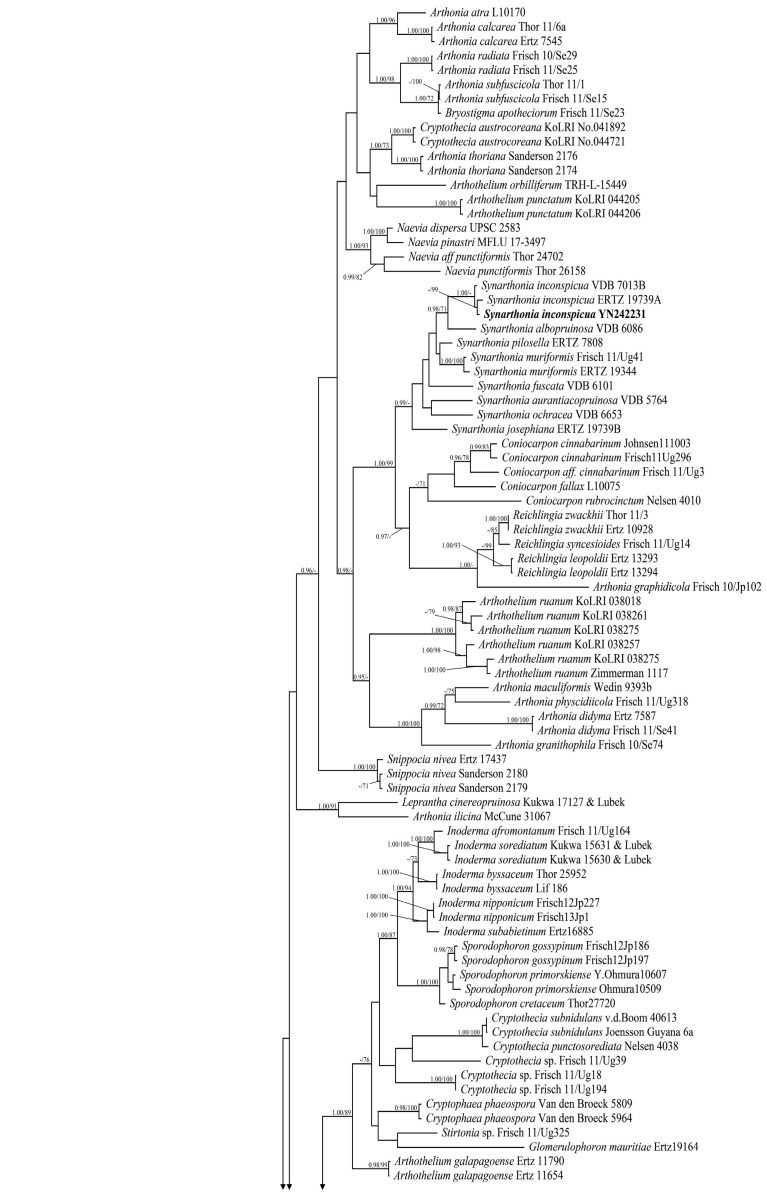 Figure 1
Figure 1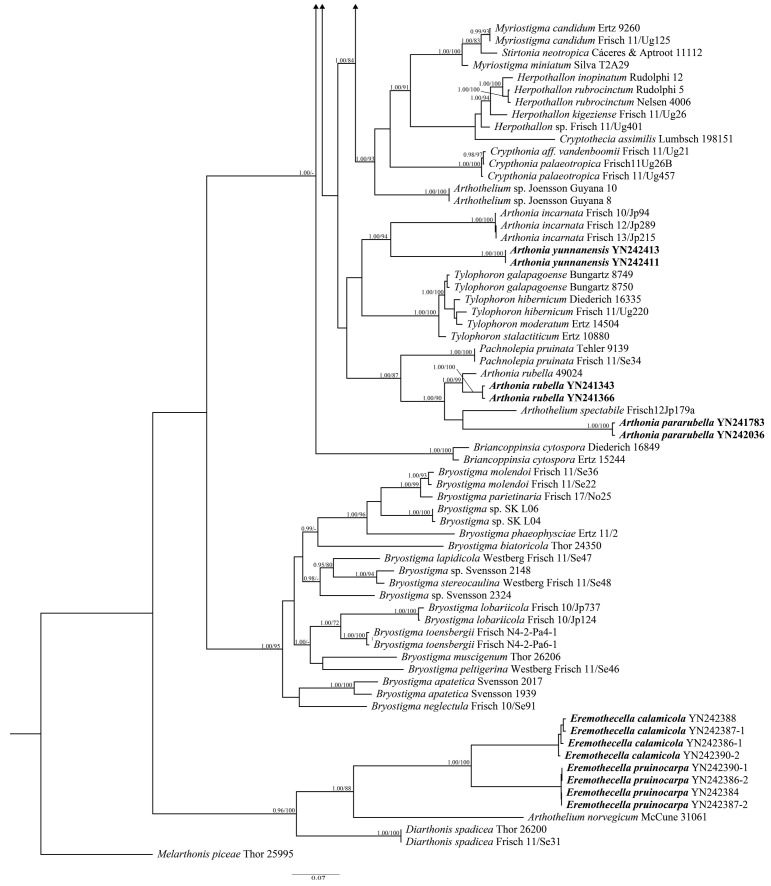 Figure 2
Figure 2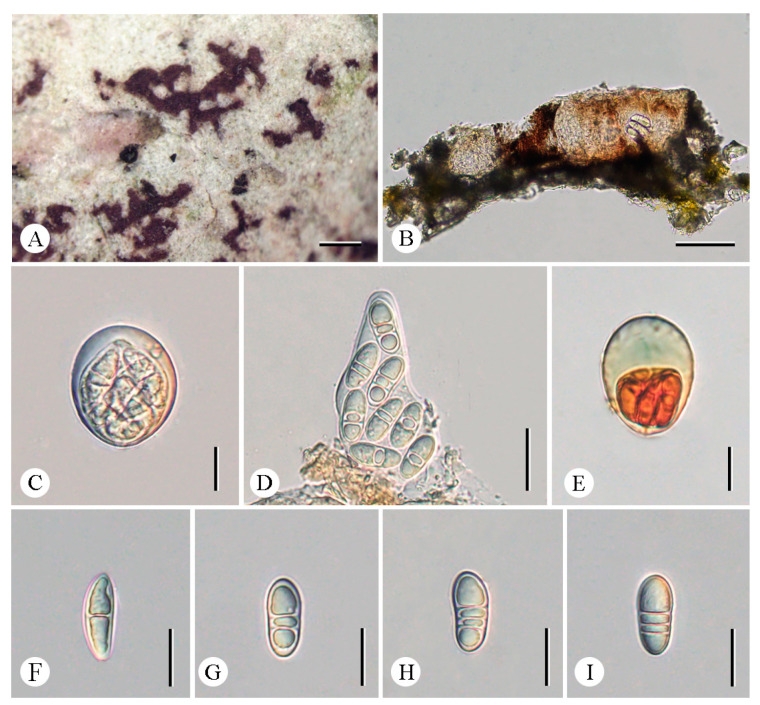 Figure 3
Figure 3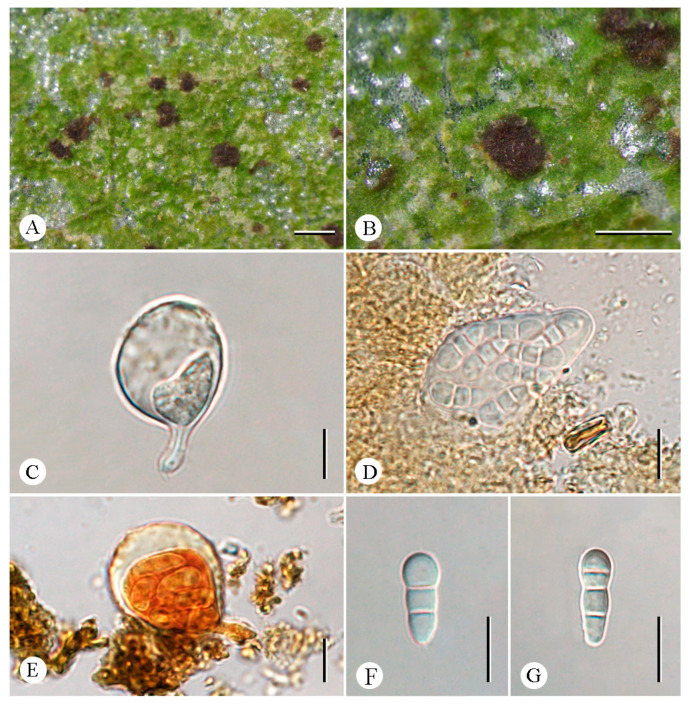 Figure 4
Figure 4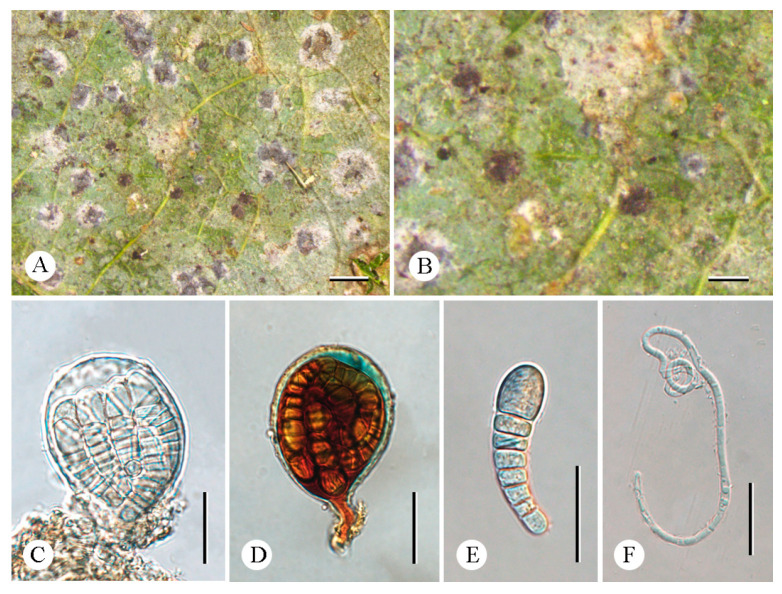 Figure 5
Figure 5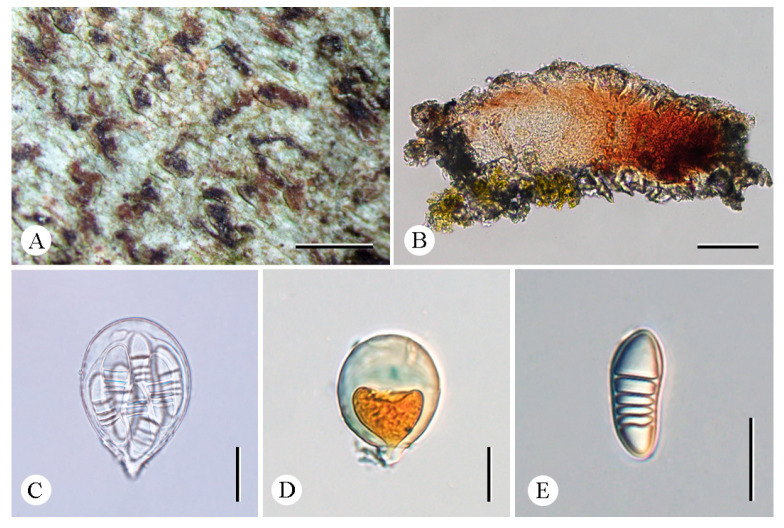 Figure 6
Figure 6 Figure 7
Figure 7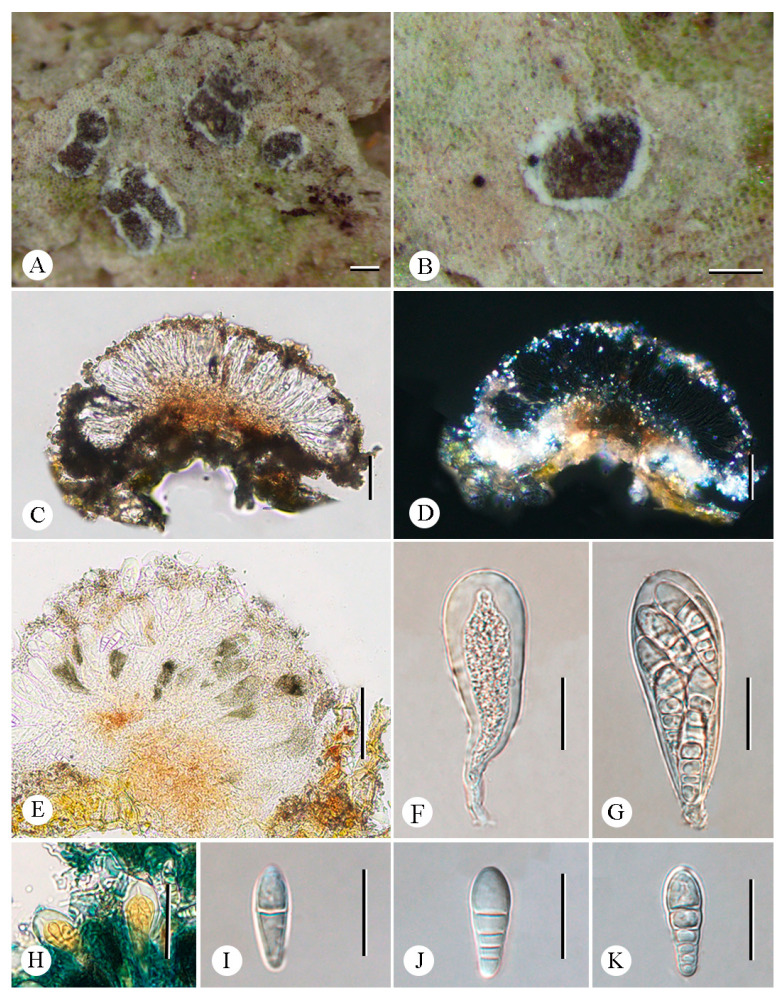 Figure 8
Figure 8- —National Natural Science Foundation of China
- —Chinese Academy of Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLichen and fungal ecology · Mycorrhizal Fungi and Plant Interactions · Biocrusts and Microbial Ecology
1. Introduction
The family Arthoniaceae Rchb. (belonging to Ascomycota, Arthoniales) is one of the largest families of lichenized fungi, and was formally delineated by Reichenbach [1]. It exhibits two important morphological features: a reduction of the ascomatal borders and globose to clavate asci with a strongly thickened apical wall, and it is not monophyletic [2,3,4,5]. This family is mainly prominent in tropical regions with abundant corticolous and foliicolous species, and it constitutes a major component of the lichen flora across diverse forest types [6]. Including genera such as Arthonia Ach., Eremothecella Syd., and Synarthonia Müll. Arg., the family currently comprises 20 genera and approximately 750 species [7,8].
Arthonia (established by Acharius in 1806) remains one of the most understudied taxa even though it encompasses approximately 500 species in the world [9]. This genus is considered to be a heterogeneous group, according to Grube et al. [10]. The lichenized representatives of Arthonia are always crustose with irregular or stellate (rarely rounded) ascocarps and have hyaline to brownish, muriform, or transversely septate (often unequally) ascospores [11]. The genus (at least in its traditional sense) comprises all different life forms: lichens, non-lichenized fungi, lichen parasites, and parasymbionts [5].
The genus Eremothecella, established by Sydow [12], shows distinctive morphological characteristics, including dark brown to black ascomata with loose paraphysoids, globose asci frequently emerging through the ascomata surface as small warts, and applanate pycnidia producing elongated filiform conidia with multiple septa, which provide taxonomic separation from Arthonia [13,14]. The genus was merged into Arthonia by Santesson [15], but subsequent revisions by Sérusiaux [13] recognized the generic distinctness of Eremothecella and formally accepted four species within this classification: viz*. E. calamicola* Syd., E. macrosperma (Zahlbr). Sérus., E. palmulacea (Müll. Arg.) Sérus., and E. variratae (Aptroot & Sipman) Sérus. Lücking (2008) further affirmed its taxonomic autonomy based primarily on the presence of Eremothecella-type pycnidia [14]. Currently, ten species are recognized: eight foliicolous and two corticolous in this genus [16].
Synarthonia was established in 1891. This small genus was the subject of a detailed discussion and report by Van Den Broeck et al. [17]. It is characterized by solitary ascomata becoming mono- to pluri-carpocentral synascomata embedded in a slightly elevated to immersed pseudostroma, with a thin white thalline margin, and Arthonia-type asci producing transversely septate ascospores, with enlarged apical cell or muriform ascospores [18]. However, the phylogeny of Synarthonia was unclear, and the synonymy of Reichlingia Diederich & Scheid had been suggested, but Ertz et al. (2020) pointed out that Synarthonia can be placed in a well-supported monophyletic lineage [18,19].
During our ongoing studies on Arthoniaceae in China, specimens belonging to three genera—Arthonia, Eremothecella, and Synarthonia—were collected. Integrative taxonomic analyses based on combined phenotypic and molecular datasets were conducted [20]. For molecular phylogenetic analyses, the mitochondrial small subunit (mtSSU) and the RNA polymerase subunit II (RPB2) were used, following the methods described by Frisch et al. (2014) and Thiyagaraja et al. (2020) for Arthoniaceae [5,6]. Three new species and three new records of Arthoniaceae from China are reported here.
2. Materials and Methods
2.1. Morphological and Chemical Analyses
Specimens were collected from Yunnan Province, China, and were deposited in the Fungarium of the College of Life Sciences, Liaocheng University (LCUF). Freshly collected specimens were first subjected to DNA extraction or preserved at −20 °C. The extracted DNA was typically stored at −80 °C, and all specimens were ultimately deposited in the herbarium. An Olympus dissecting microscope (Olympus SZX16, Olympus Corporation, Tokyo, Japan) and an Olympus light microscope (Olympus BX53, Olympus Corporation, Tokyo, Japan) were used for morphological and anatomical studies. Measurements were taken from mature vertical sections of fruit bodies mounted in water to observe whether they contained crystals: the shape and size of the photobiont, the width of the hyphae, and the color, shape, and size of the asci, ascospores, pycnidia, and conidia. The amyloidity of ascospores was tested with Lugol’s solution. Spot tests with K (10% aqueous solution of potassium hydroxide), C (saturated solution of aqueous sodium hypochlorite), P (saturated solution of p-phenylenediamine in 95% ethylalcohol), and UV (fluorescence at 365 nm) were performed on the surface of the thallus [21]. The lichen substances were detected and identified using a UV light source and thin-layer chromatography (TLC) in solvent C [22].
2.2. DNA Extraction and PCR Sequencing
Genomic DNA was extracted from the ascomata and thallus of the specimens using a DNA secure Plant Kit (Tsingke, Beijing, China) according to the manufacturer’s instructions. The mtSSU and RPB2 regions were amplified using the primer pair mtSSU1/3R [23] and RPB2-7cF/11aR [24]. Polymerase Chain Reaction (PCR) cycling conditions were set to an initial denaturation at 95 °C for 5 min, followed by 35 cycles of denaturation at 95 °C for 45 s, annealing at 50 °C for 1 min, extension at 72 °C for 1.5 min, and a final extension at 72 °C for 10 min [25]. The PCR products were checked on 1% electrophoresis gels, stained with ethidium bromide, and sequenced by Tsingke Biotech Co., Ltd. (Beijing, China).
2.3. Phylogenetic Analyses
The new sequences were assembled via Geneious v.9.0.2 (Biomatters Ltd., Auckland, New Zealand) and potential sample contamination were confirmed by BLASTn searches on the NCBI website (http://www.ncbi.nlm.nih.gov/BLAST/, accessed on 17 April 2025). Then, high-similarity sequences of Arthoniaceae were selected for further analysis (Table 1), with Melarthonis piceae Frisch & G. Thor (belonging to Chrysotrichaceae, Arthoniales) as the outgroup [5]. The assembled sequences were aligned with the online version of MAFFT v.7 (https://mafft.cbrc.jp/alignment/server/, accessed on 17 April 2025). Ambiguously aligned regions were delimited using Gblocks v.0.91b with the least stringent selection [26]. Geneious v.9.0.2 was used to concatenate the mtSSU and RPB2 genes and produce a two-locus dataset. Maximum likelihood (ML) analysis and Bayesian inference (BI) were used to infer phylogenetic trees based on the concatenated mtSSU and RPB2 datasets. The maximum likelihood analysis was conducted with RAxML-HPC 2 on ACCESS v.8.2.12 employing a GTRGAMMA approximation with a rapid bootstrap analysis of 1000 replicates on the CIPRES Scientific gateway portal (http://www.phylo.org/portal2/, accessed on 17 April 2025) for verification [27,28]. For BI analysis, PartitionFinder 2 [29] was used to determine the best-fit model for each partition. Based on the results, we used the GTR+I+G model for mtSSU and the HKY+I+G model for RPB2. Bayesian Inference phylogenies were inferred using MrBayes v.3.2.7a under a partition model [30]. We ran 2 million generations, in which the initial 25% of sampled data was discarded as burn-in. Bayesian inference posterior probability (BIPP) equal to and above 95% was considered significant and shown at the nodes. Generated phylogenetic trees were visualized under Figtree v.1.4.4 and edited in Adobe Illustrator CC2019 software.
3. Results
3.1. Phylogenetic Results
A total of 150 specimens comprising 227 DNA sequences (139 mtSSU sequences and 88 RPB2 sequences), including fifteen mtSSU sequences and eight RPB2 sequences that were newly generated were used in this study (Table 1). For the two-locus (mtSSU and RPB2) dataset taxon set, the concatenated alignment contained 1544 characters (787 for mtSSU and 757 for RPB2). The RAxML tree was constructed with a final ML optimization likelihood value of −27,812.975243. The parameters for the GTR+I+G model of combined mtSSU and RPB2 were as follows: estimated base frequencies: A = 0.290705, C = 0.231758, G = 0.228597, T = 0.248940; substitution rate AC = 1.017805, AG = 5.448594, AT = 1.720312, CG = 1.388442, CT = 5.952238, and GT = 1.000000. The resulting topology was similar for maximum likelihood (ML) and Bayesian analysis. Therefore, only the Bayesian tree is presented, with bootstrap support ≥70% for ML analysis and posterior probabilities ≥0.95 for Bayesian analysis (Figure 1).
In the phylogenetic tree (Figure 1), the main well-supported lineages of Arthoniaceae follow the results obtained by Frisch et al. (2014) and agree well with previous hypotheses on the polyphyletic origins of parasitic Arthonia [6]. Our phylogenetic results also support that Eremothecella forms an independent clade within the family Arthoniaceae based on the sequence of Eremothecella pruinocarpa (YN242390-1, YN242386-2, YN242384, YN242387-2) and E. calamicola (YN242388, YN242387-1, YN242386-1, YN242390-2), which were newly generated in this study. These are also the first reported molecular sequences of species in the genus Eremothecella.
3.2. Taxonomy
Arthonia pararubella C.Y. Hao, S.H. Jiang, and Z.F. Jia, sp. nov. (Figure 2).
Fungal Name: FN 572511.
Diagnosis. The new species externally resembles Arthonia rubella (Fée) Nyl. but differs in the shorter ascospores and fewer septa.
Type. China. Yunnan Province: Mengla County, Xishuangbanna Tropic Botanical Garden, Chinese Academy of Sciences, Baihuayuan, 21°55′26″ N, 101°15′02″ E, alt. 560 m, on bark, 18 December 2024, C.Y. Hao (LCUF YN242036—holotype).
**Description. **Thallus greenish gray to whitish gray. Prothallus brownish-black line. Ascomata reddish-brown to blackening, innate, maculate to lirellate, sometimes branched or stellate, thinner at the branched ending, 0.2–1.65 × 0.05–0.2 mm. Excipulum carbonized, 21.5–30 μm thickness. Hymenium 67–77 µm tall, pale brown to brown, K/I+ blue. Asci clavate to subglobose, 47.5–66.5 × 28–44 µm ( = 55 × 35 μm), Arthonia-type, eight-spored, I+ orange-red, apical part thickened, usually with a distinct ocular chamber, K/I−. Ascospores oblong–ovoid, (2–)3-septate, upper and lower cells enlarged, ellipsoidal locules in immaturity, colorless, 25.5–29.5 × 10–13.5 μm ( = 28 × 12.5 μm), I+ orange-red.
Chemistry. Thallus K-, C-, KC-, UV-. No substances were detected by TLC.
Ecology and distribution. The new species is currently only known in China on the bark of trees in a tropical forest in Yunnan Province.
Etymology. The specific epithet pararubella refers to the similarity to Arthonia rubella.
Additional specimens examined. China. Yunnan Province: Jinghong City, Mengyang Town, Wild Elephant Valley, 22°09′58″ N, 100°51′49″ E, alt. 800 m, on bark, 16 December 2024, C.Y. Hao (LCUF YN241783).
Notes. Arthonia pararubella can be distinguished from similar species by its hyaline ascospores (25.5–29.5 × 10–13.5 μm) with two to three septa. Arthonia rubella is morphologically similar to this new species, but the former can be distinguished by longer ascospores with more septa (3–5-septate vs. (2–)3-septate; 25–43 × 10–15 μm vs. 25.5–29.5 × 10–13.5 μm) [31,32,33]. Further, both are far apart in the phylogenetic tree (Figure 1). Another similar species, A. compensatula Nyl. differs from this new species by its ascospores with more septa (4–5-septate vs. (2–)3-septate) and without amphicephalic [33,34].
Arthonia yunnanensis C.Y. Hao, S.H. Jiang, and Z.F. Jia, sp. nov. (Figure 3).
Fungal Name: FN 572510.
Diagnosis. The new species externally resembles Arthonia incarnata Kullh. ex Almq, but differs in the subglobose asci, color of the thallus, and that it lives on leaves.
Type. China. Yunnan Province: Mengla County, Xishuangbanna Tropic Botanical Garden, Chinese Academy of Sciences, lvshilin, 21°54′17″ N, 101°16′28″ E, alt. 625 m, on leaves, 18 December 2024, C.Y. Hao (LCUF YN242413—holotype).
Description.* *Thallus uncontinuous, yellowish green to green, thin, matte to weakly glossy. Prothallus absent. Photobiont trentepohlioid, cells globose to long elliptical. Ascomata dispersed, maculate, usually slightly irregularly roundish to short elliptical, adnate, flat to weakly convex, pale and often brownish to dark brown, 0.25–0.4 mm in diameter. Hymenium near hyaline to pale orange-brown, K/I+ blue. Asci subglobose, 27–32 × 18–23 μm ( = 29 × 20.5 μm), Arthonia-type, thin lateral walls, stipitate, eight-spored, usually with a distinct ocular chamber, K/I−. Ascospores clavate, 2(–3)-septate, usually with an enlarged distal cell, not curved, with slight constrictions at septa, colorless, 12.5–15.5 × 4.5–6.5 μm ( = 14 × 5.5 μm).
Chemistry. Thallus K-, C-, KC-, UV-. No substances were detected by TLC.
Ecology and distribution. The new species is currently only known in China on the leaves of trees in a tropical rainforest in Yunnan Province.
Etymology. The specific epithet yunnanensis refers to the type locality, Yunnan Province, Mengla County.
Additional specimens examined. China. Yunnan Province: Mengla County, Xishuangbanna Tropic Botanical Garden, Chinese Academy of Sciences, Greenstone Forest, 21°54′17″ N, 101°16′28″ E, alt. 625 m, on leaves, 18 December 2024, C.Y. Hao (LCUF YN242411).
Notes. Arthonia yunnanensis, the only species from southern China’s tropical region inhabiting leaves, is characterized by subglobose asci. It resembles Arthonia incarnata, A. accolens Stirt., and A. lividula Vain. in having 2-septate ascospores [4,15,35,36]. However, A. incarnata resembles the new species, having the same size ascospores (11.7–15.3 × 4.2–5.4 μm vs. 12.5–15.5 × 4.5–6.5 μm), but its thallus is often inapparent or visible as a whitish to pale olive-gray discoloration of bark; further, its asci are longer and narrower (40–50 × 14–20 μm vs. 27–32 × 18–23 μm) [4]. In the phylogenetic tree (Figure 1), although A. yunnanensis forms a sister clade to A. incarnata, the two clades are distinctly separated by a long genetic distance and exhibit substantial divergence. Therefore, A. yunnanensis as an independent lineage also supports its status as a species distinct from A. incarnata. Arthonia accolens can be distinguished by larger ascomata (often >0.5 mm diam.) [15,36]. Arthonia lividula differs from this new species in having shorter asci and smaller ascospores without constrictions at the septa (9–12 × 3–5 μm vs. 12.5–15.5 × 4.5–6.5 μm) [15,35]. Morphologically, A. yunnanensis was revealed as significantly different from all known species of Arthonia (see notes below), and therefore we describe this species here as a new species.
Eremothecella pruinocarpa C.Y. Hao, S.H. Jiang, and Z.F. Jia, sp. nov. (Figure 4).
Diagnosis. The new species externally resembles Eremothecella ajaysinghii Jagad. Ram & G.P. Sinha but differs in its fewer septa of ascospores.
Type. China. Yunnan Province: Jinghong City, tropical rainforests, 21°54′31″ N, 101°10′58″ E, alt. 635 m, on leaves, 17 December 2024, C.Y. Hao (LCUF YN242390-1—holotype).
**Description. **Thallus crustose, greenish gray, dispersed or continuous, matte. Prothallus absent. Photobiont trentepohlioid, cells in radiate plates, rectangular, ca. 11 × 4 μm. Ascomata rounded to irregular, dark brown to black, ± moderately to densely grayish pruinose, pruina sometimes increasingly dense along the margin, 0.2–1.15 mm diam. Asci obovate to globose, 45–60 × 36.5–45.5 μm ( = 54.5 × 41 μm), stipitate, eight-spored, K/I+ red, usually with a distinct ocular chamber, K/I+ blue. Ascospores colorless, clavate, curved, (6–)7(–9)-septate, enlarged distal cell, very slight constrictions at septa, 30–49 × 8–11.5 μm ( = 38 × 9.5 μm), I+ orange-red. Pycnidia numerous, ± rounded, irregular, black, 0.2–0.45 mm diam. Conidia colorless, filiform, multiseptate, 80–130 × 1.5–2 µm ( = 100 × 2 μm).
Chemistry. Thallus K-, C-, KC-, UV-. No substances were detected by TLC.
Ecology and distribution. The new species is currently only known in China on the leaves of trees in a tropical rainforest in Yunnan Province.
Etymology. The specific epithet pruinocarpa refers to the densely whitish gray ascomata.
Additional specimens examined. China. Yunnan Province: Jinghong City, tropical rainforests, 21°54′31″ N, 101°10′58″ E, alt. 635 m, on leaves, 17 December 2024 X. Li (LCUF YN242384; LCUF YN242386-2; LCUF YN242387-2).
Notes. Eremothecella pruinocarpa differs from similar species in its grayish pruinose ascomata and (6–)7(–9)-septate ascospores. Eremothecella ajaysinghii resembles the new species E. pruinocarpa in having grayish pruinose ascomata but differs in having ascospores with more septa [8–10(–11)-septate vs. (6–)7(–9)-septate] [16]. Eremothecella macrosperma is another similar species, particularly in ascospore size, but its ascomata are non-pruinose [15]. Eremothecella cyaneoides Lücking differs from this new species in having ascospores with fewer septa (3–5-septate vs. (6–)7(–9)-septate) [37]. Phylogenetically, four specimens of Eremothecella pruinocarpa form a strongly supported monophyletic clade, which also supports that it is a new species (Figure 1).
Arthonia rubella (Fée) Nyl. (Figure 5).
Fungal Name: FN 119459.
Basionym. Graphis rubella Fée, Essai Crypt. Exot. (Paris): 43 (1825) [1824].
**Description. **Thallus greenish gray to green, or whitish gray. Prothallus line brownish black. Photobiont trentepohlioid, cells globose. Ascomata reddish-brown and black, innate, maculate to lirellate, flexuous to irregularly branched, 0.25–1.2 mm long, disk occasionally opening. Asci clavate to subglobose, Arthonia-type, eight-spored, apical part thickened, usually with a distinct ocular chamber. Ascospores oblong–ovoid, (3–)4(–5)-septate, upper and the lower cell enlarged, colorless, 29–35 × 12–15 μm.
Chemistry. Thallus K-, C-, KC-, UV-. No substances were detected by TLC.
Ecology and distribution. Growing on exposed trees in tropical forests. Initially reported from America (as ‘Graphis rubella’) [31]. Newly reported in China.
Additional specimens examined. China. Yunnan Province: Puer City, Jingdong County, Jinping Town, Fork River Bridge, 24°37′30″ N, 100°45′22 ″ E, alt. 1320 m, on bark, 16 August 2024, C.Y. Hao (LCUF YN241343; LCUF YN241366).
Notes. The morphological and anatomical characteristics of this study align with Arthonia rubella from America as described by Fée (as ‘Graphis rubella’) [31]. The vast majority of records document this species on bark substrates in tropical South America [31,38]. It is closely related to A. compensatula in morphology, but the latter species has no amphicephalic ascospores [31,33,34]. It is also similar to A. acanthotheciicola Ertz & Common in ascospore morphology and septation, with both featuring enlarged upper and lower cells; however, the latter typically has smaller ascospores (14.5–17 × 6–6.5 µm) [39]. The newly generated A. rubella sequences from this study are clustered with the published sequences of the same species in the phylogenetic tree (Figure 1).
Eremothecella calamicola Syd. (Figure 6).
Fungal Name: FN 145347.
**Description. **Thallus crustose, dispersed or continuous, smooth, green, thin, matte. Prothallus absent. Photobiont trentepohlioid, cells rectangular to irregular, ca. 9 × 5 μm. Ascomata rounded to irregular in outline, 0.3–1 mm diam., dark brown to blackish brown, non-pruinose. Asci obovate to globose, 37.5–52.5 × 33.5–40.5 µm, stipitate, eight-spored, usually with a distinct ocular chamber, K/I+ pale blue. Ascospores colorless, clavate, curved, 5–6-septate, enlarged distal cell, with very slight constrictions at septa, (28.3–)30.5–38 × 8.5–10.5 μm, I+ orange-red.
Chemistry. Thallus K-, C-, KC-, UV-. No substances were detected by TLC.
Ecology and distribution. Grows exposed trees in the shaded parts of evergreen forests. Previously reported from the Philippines [12,15], New Guinea [15], Mexico, Costa Rica, Panama, Guyana, French Guiana, Ecuador, Peru, Brazil [14], and India [16]. Newly reported in China.
Additional specimens examined. China. Yunnan Province: Jinghong City, tropical rainforests, 21°54′31″ N, 101°10′58″ E, alt. 635 m, on leaves, 17 December 2024, X. Li (LCUF YN242386-1; LCUF YN242387-1; LCUF YN242388); C.Y. Hao (LCUF YN242390-2).
**Notes.**Eremothecella calamicola resembles Arthonia palmulacea (Müll. Arg.) R. Sant. but the latter has shorter ascospores with fewer septa (16–25 × 4–8 μm vs. (28.3–)30.5–38 × 8.5–10.5 μm; 3–5-septate vs. 5–6-septate) [15,16]. Eremothecella variratae is another similar species regarding ascospore size (28–42 × 7–10 µm vs. (28.3–)30.5–38 × 8.5–10.5 μm), but its ascomata often feature orange-yellow pruinose [15,16]. Another morphologically similar species, E. nicobarica Jagad. Ram and G. P. Sinha. can be distinguished by larger ascospores with more septa ((14–)15–17-septate vs. 5–6-septate; 60–72 × 10–13 µm vs. (28.3–)30.5–38 × 8.5–10.5 μm) [15,16]. In the phylogenetic tree, a well-supported monophyletic clade comprising the E. calamicola specimens is sister to the new species E. pruinocarpa (Figure 1).
Synarthonia inconspicua (Stirt.) Van den Broeck and Ertz. (Figure 7).
Fungal Name: FN 825152.
Basionym. Arthonia inconspicua Stirt., Proceedings of the Philosophical Society of Glasgow 11: 319. 1879 [40].
**Description. **Thallus whitish to greenish-gray, smooth, continuous to cracked. Photobiont trentepohlioid, cells 9 µm in diam., rounded, usually in chains. Ascomata usually solitary, 0.25–0.6 × ca. 0.2 mm, rounded to lirellate, often forming irregular clusters, slightly elevated above thallus level, scattered more or less evenly over the thallus; disk heavily white pruinose in margin and thinner pruinose in the middle, light brown when pruina removed, flat to convex. Excipulum 16.5–21.5 µm wide, loosely intricate hyphae, inspersed with orange-brown granules which are K+ completely dissolving. Epihymenium 3.5–24.5 µm tall, and adspersed with orange-brown granules which are K+ completely dissolving. Hymenium 41.5–57.5 µm tall, hyaline, not inspersed, K/I+ blue. Paraphysoids loosely intricate around the asci. Hypothecium 17–34 µm-thick, yellowish brown, composed of loosely intricate hyphae, inspersed with orange granules which are K+ completely dissolving, K/I+ blue. Asci 49–65 × 17–24 µm, clavate, obovoid to ellipsoid or globose, stipitate, occasionally with a distinct ocular chamber, K/I−. Ascospores 17.5–24 × 6.5–8 µm, hyaline, with enlarged apical cell, oblong–ovoid, (1–)3–4(–5)-septate, spore ontogeny macrocephalic, unidirectional.
Chemistry. Thallus K-, C-, KC-, UV+. No substances were detected by TLC.
Ecology and distribution. Growing on exposed trees in tropical forests. Previously reported in the tropics of Tamil Nadu [40], India [41,42,43], Cuba, D.R. Congo, India, Madagascar, Netherlands Antilles, Rwanda, Sierra Leone, Tanzania, Uganda, and USA [17]. Newly reported in China.
Additional specimens examined. China. Yunnan Province: Mengla County, Mengla Fairyland, Menglazi Naturn Reserve, 21°42′44″ N, 101°22′32″ E, alt. 700 m, on bark, 19 December 2024, C.Y. Hao (LCUF YN242231).
Notes. The similar morphological species Synarthonia albopruinosa Van den Broeck and Ertz. differs from S. inconspicua in smaller ascospores with fewer septa (12.5–17.5 × 5–6.5 µm vs. 17.5–24 × 6.5–8 µm; (1–)2–3-septate vs. (1–)3–4(–5)-septate) [17]. Additionally, S. fuscata Van Den Broeck and Ertz. can be distinguished from S. inconspicua by the absence of pruina on the ascomata and the presence of a K/I+ blue ring-like structure in the asci [17]. In terms of molecular structure, newly generated sequences in this study of S. inconspicua were clustered with the published sequences of this species in the phylogenetic tree (Figure 1).
4. Discussion
Molecular analyses are indispensable for the classification and species identification of many lichen species within a genus due to their scarce morphological distinctions [44,45]. Employing morphological and phylogenetic analyses, this study identified three new species: Arthonia yunnanensis, A. pararubella, and Eremothecella pruinocarpa, and three new records for China: A. rubella, E. calamicola, and Synarthonia inconspicua from Yunnan Province, China. This study indicates the high species diversity of the family Arthoniaceae in Yunnan Province, China.
In the family Arthoniaceae, many taxa possess only phenotypic data but lack genotypic information, which has led to confusion regarding the taxonomic status; for example, Eremothecella. It needs to be pointed out that previous studies on this genus were mainly based on morphology, but this research provides the first molecular data for Eremothecella. Our molecular results also show that Eremothecella (E. pruinocarpa and E. calamicola) forms an independent clade within Arthoniaceae. Furthermore, to improve the clarity of generic relationships within the Arthoniaceae, sustained taxonomic investigations should be conducted in the future. Thorough surveys of the regional lichen flora will undoubtedly yield additional specimens for analysis. Empirical evidence indicates that in-depth research will uncover rarer species of Arthoniaceae, along with a considerable number of hitherto undescribed taxa.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Reichenbach H.T.L. Der Deutsche Botaniker Her barienbuch D. Arnoldische Buchandlung Leipzig, Germany Dresden, Germany 18411240
- 2Tehler A. A new approach to the phylogeny of Euascomycetes with a cladistic outline of Arthoniales focussing on Roccellaceae Can. J. Bot.1990682458249210.1139/b 90-312 · doi ↗
- 3Grube M. Classification and Phylogeny in the Arthoniales (Lichenized ascomycetes)Bryologist 199810137739110.1639/0007-2745(1998)101[377:CAPITA]2.0.CO;2 · doi ↗
- 4Frisch A. Thor G. Moon K.H. Ohmura Y. Arthonia incarnata (Arthoniaceae), a rare and poorly known old-growth forest lichen new to Asia Nord. J. Bot.20173558759410.1111/njb.01560 · doi ↗
- 5Thiyagaraja V. Lücking R. Ertz D. Wanasinghe D.N. Karunarathna S.C. Camporesi E. Hyde K.D. Evolution of non-lichenized, saprotrophic species of Arthonia (Ascomycota, Arthoniales) and resurrection of Naevia, with notes on Mycoporum Fungal Divers.202010220522410.1007/s 13225-020-00451-9 · doi ↗
- 6Frisch A. Thor G. Ertz D. Grube M. The Arthonialean challenge: Restructuring Arthoniaceae Taxon 20146372774410.12705/634.20 · doi ↗
- 7Aptroot A. Cáceres M.E.D.S. Santos L.A.D. The taxonomy of sterile Arthoniaceae from Brazil: White crusts on overhanging tropical trees can be named Lichenologist 20245611310.1017/S 0024282924000021 · doi ↗
- 8Lücking R. Hodkinson B.P. Leavitt S.D. The 2016 classification of lichenized fungi in the Ascomycota and Basidiomycota—Approaching one thousand genera Bryologist 201711936110.1639/0007-2745-119.4.361 · doi ↗