Identification and Expression Analysis of the Cytochrome P450 Genes in Phyllotreta striolata and CYP6TH1/CYP6TH2 in the Involvement of Pyridaben Tolerance
Yongqin Zhu, Zhongting Liu, Wenyong Mai, Xinhua Pu, Haoyue Mo, Benshui Shu, Zhongzhen Wu

TL;DR
This study identifies 94 cytochrome P450 genes in Phyllotreta striolata and shows that CYP6TH1 and CYP6TH2 are key to pyridaben tolerance, offering new targets for pest resistance management.
Contribution
The study identifies CYP6TH1 and CYP6TH2 as novel genes involved in pyridaben tolerance in Phyllotreta striolata.
Findings
94 cytochrome P450 genes were identified and classified in Phyllotreta striolata.
CYP6TH1 and CYP6TH2 were highly expressed in the head and fat body and upregulated by pyridaben exposure.
RNA interference knockdown of CYP6TH1 and CYP6TH2 increased pyridaben susceptibility in P. striolata.
Abstract
Pyridaben is extensively utilized to manage Phyllotreta striolata; nevertheless, the emergence of resistance from continuous use has created difficulties for its efficient control. At present, research on the significance of Cytochrome P450 genes in imparting pesticide resistance in Phyllotreta striolata is scarce. This research discovered 94 cytochrome P450 genes in Phyllotreta striolata, examined their spatiotemporal expression patterns, and demonstrated that CYP6TH1 and CYP6TH2 are crucial for pyridaben tolerance. The striped flea beetle, Phyllotreta striolata, is a worldwide pest that causes severe damage to cruciferous crops. Pyridaben, an inhibitor of mitochondrial complex I, has been widely used for the control of P. striolata. Identifying key genes involved in pyridaben tolerance is vital for establishing effective resistance management strategies. A total of 94 full-length…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
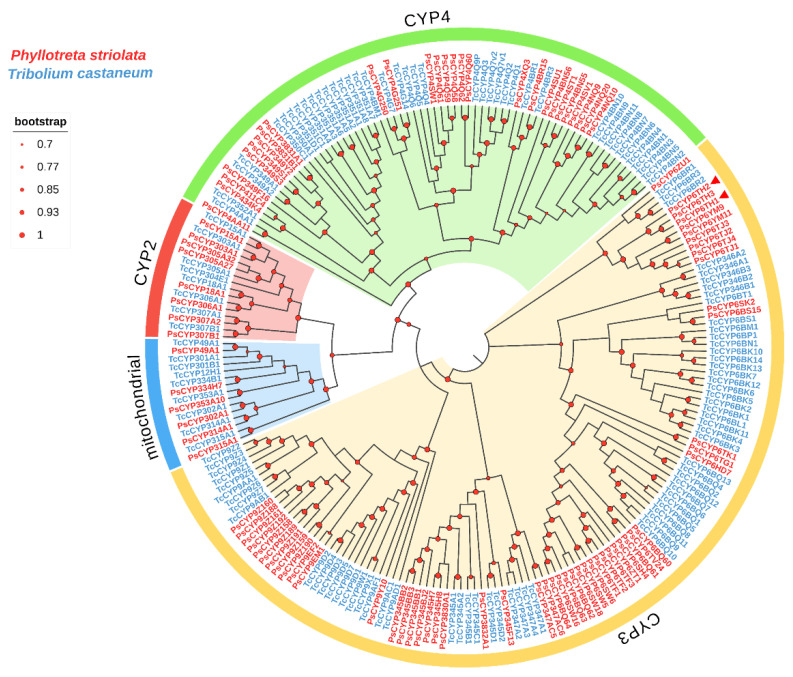 Figure 1
Figure 1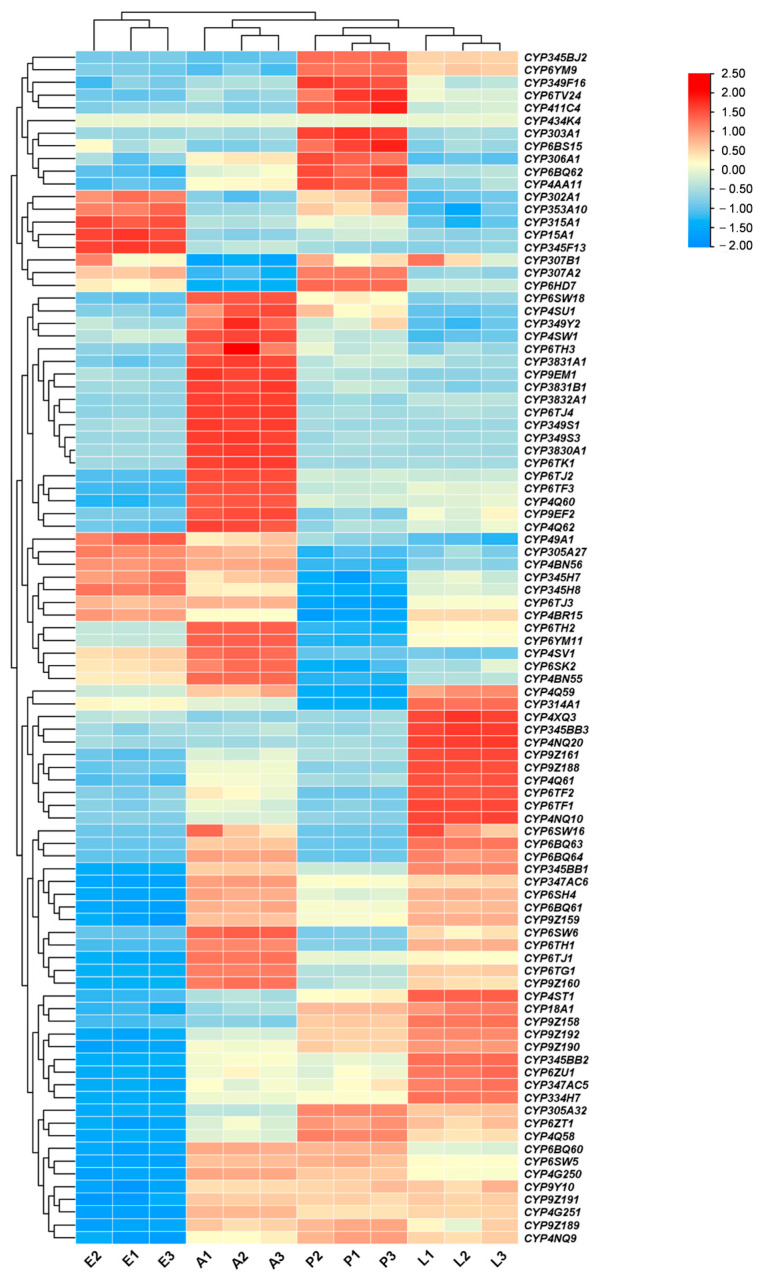 Figure 2
Figure 2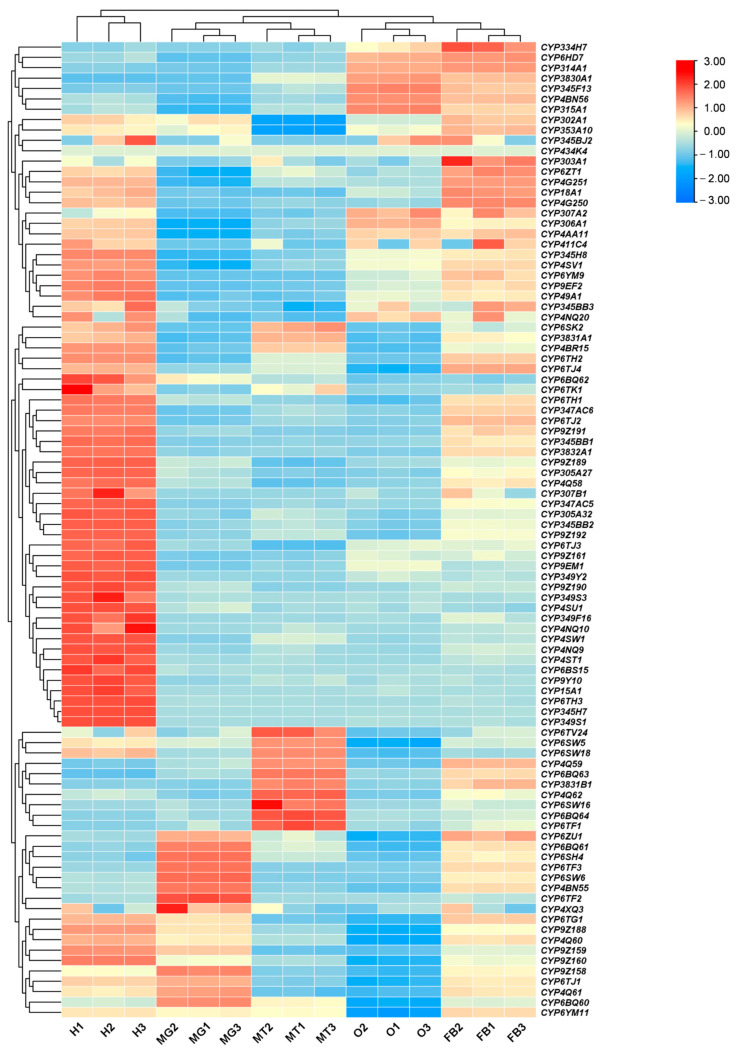 Figure 3
Figure 3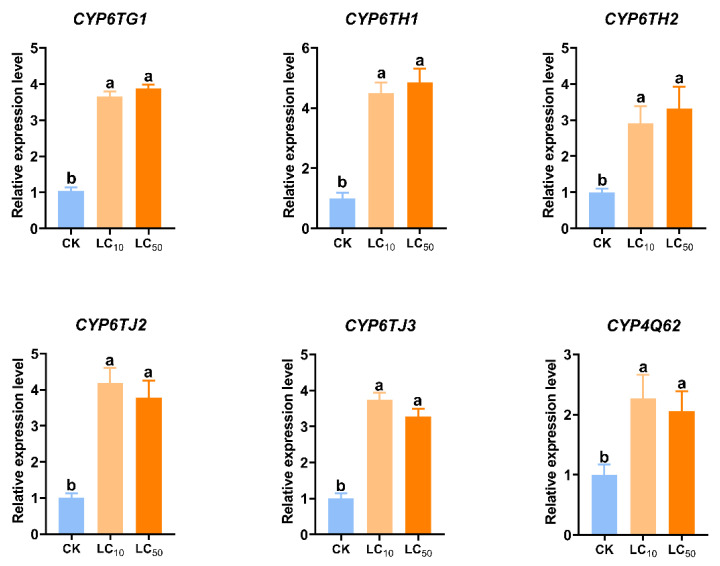 Figure 4
Figure 4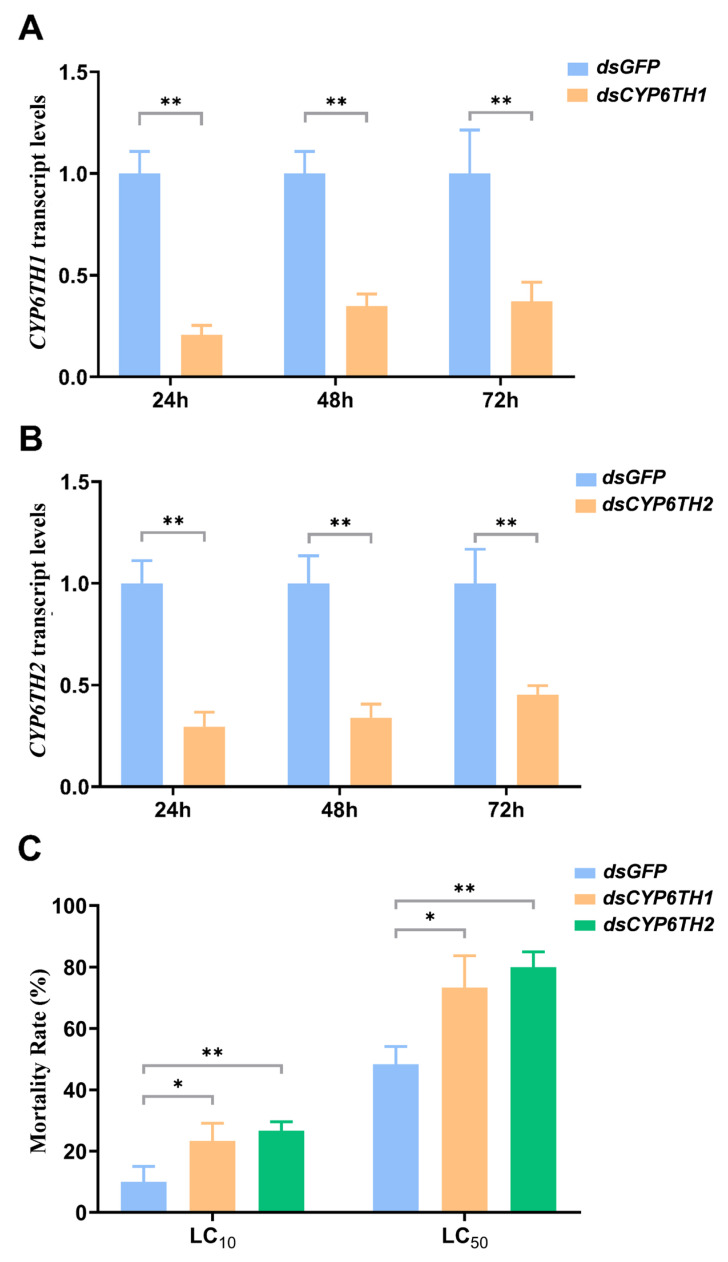 Figure 5
Figure 5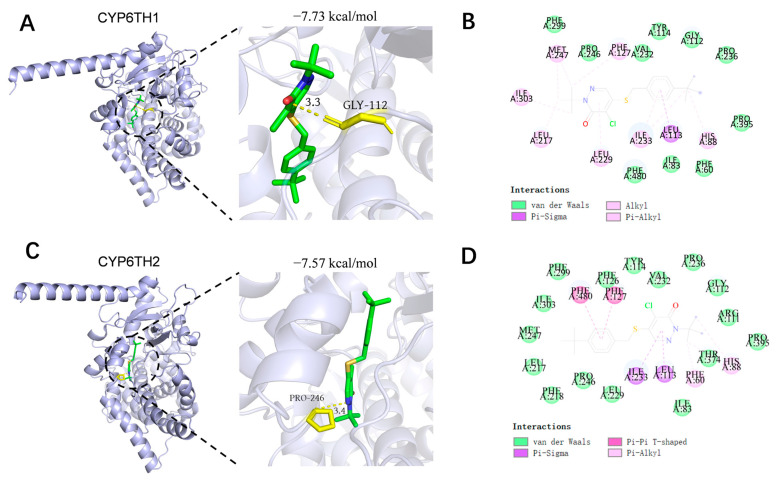 Figure 6
Figure 6- —Guangdong Provincial University Innovation Team Project
- —Guangdong Province key construction discipline Scientific Research Capacity Improvement Program
- —open competition program of top ten critical priorities of Agricultural Science and Technology Innovation for the 14th Five-Year Plan of Guangdong Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect Resistance and Genetics · Insect-Plant Interactions and Control · Neurobiology and Insect Physiology Research
1. Introduction
The striped flea beetle, Phyllotreta striolata (Fabricius) (Coleoptera: Chrysomelidae), is a destructive pest of cruciferous vegetables [1]. Both larvae and adults cause significant damage to plants: adults feed on the leaf mesophyll, producing characteristic “shot-hole” symptoms, and in severe cases, may consume the entire mesophyll layer. Larvae can damage main roots and root hairs, leading to plant wilting and eventual death [2]. Chemical insecticides such as fipronil, chlorpyrifos, phoxim, imidacloprid, and acetamiprid are widely used to control P. striolata [3,4,5]. However, intensive and repeated application has led to the emergence of resistant strains in recent years [6].
Pyridaben, a pyridazinone derivative that inhibits mitochondrial complex I in the electron transport chain [7], is another extensively used insecticide. As a miticide, pyridaben is also extensively used to control P. striolata [8]. For example, field trials have shown that 15% pyridaben EC is among the most effective insecticides recommended for managing this pest [9]. Additionally, combinations such as pyridaben–acetamiprid WP show significant synergy [10]. At the same time, many pest species around the world have been found to be very resistant to pyridaben. For instance, Tetranychus urticae [11] and Panonychus citri [12] have shown strong resistance to pyridaben in fields. In Yunnan Province, China, field populations of Tetranychus cinnabarinus exhibited resistance ratios to pyridaben ranging from 1.50- to 4.68-fold [13]. High levels of resistance of P. striolata to pyridaben have also been observed in Guangdong Province, where pyridaben has been applied extensively for insect pest management [6]. Thus, a detailed understanding of how P. striolata responds to pyridaben at the molecular level is essential.
Cytochrome P450 monooxygenases (P450s or CYPs), one of the largest enzyme superfamilies across living organisms, play crucial roles in metabolizing both endogenous and exogenous compounds, including steroids, fatty acids, and insecticides. The constitutive overexpression of specific P450 genes is a well-documented mechanism underlying insecticide resistance [14]. For example, CYP6DB3 is upregulated in thiamethoxam- and imidacloprid-resistant strains of Bemisia tabaci [15]. Elevated P450 activities and overexpression of CYP6BG1 have been linked to chlorantraniliprole resistance in Plutella xylostella [16]. In Spodoptera exigua, multiple CYP6AE genes, particularly CYP6AE70, are overexpressed and associated with cross-resistance to chlorpyrifos, cypermethrin, and deltamethrin [17]. Additionally, pyridaben exposure upregulates CYP4CF1 and CYP4CL2 in Panonychus citri [18]. However, the involvement of P450 genes in insecticide resistance has been poorly investigated in P. striolata.
In this study, we identified 94 full-length P450 genes and analyzed their expression patterns across different developmental stages and tissues. We further investigated the response of these P450 genes to sublethal pyridaben exposure (LC_10_ and LC_50_). Knockdown of these two genes via RNA interference (RNAi) increased susceptibility of P. striolata to pyridaben. The potential interactions between CYP6TH1/CYP6TH2 and pyridaben were analyzed through molecular docking experiments. Our findings offer new insights into the molecular mechanisms of P450-mediated pyridaben tolerance in P. striolata.
2. Materials and Methods
2.1. Insect Rearing
The P. striolata population used in this study was a laboratory colony originally collected from a crucifer crop field in Fujian province, China. Since 2014, the strain has been maintained on mustard plants (Brassica juncea cv. Bau-Sin) in a greenhouse at Zhongkai University of Agriculture and Engineering without exposure to insecticides. Rearing conditions were as follows: temperature 21–26 °C, relative humidity 60–80%, and soil humidity 95%. Natural daylight was used as the light source in the greenhouse.
2.2. Toxicological Bioassay
The susceptibility of P. striolata adults to pyridaben (analytical standard, Sigma-Aldrich, St. Louis, MO, USA) was determined using the leaf-dip bioassay method. A stock solution of pyridaben was prepared in acetone and diluted to various concentrations for assays. Fresh, uniform radish leaf disks (diameter: 2 cm) were dipped in the corresponding insecticide solutions for 10 s, with control leaf disks dipped in acetone only. The treated leaf disks were air-dried for approximately 2 h to allow the solvent to evaporate completely. For each concentration, 60 actively moving adults (3–5 days old; 20 adults for each replicate, and three replicates were prepared) were starved for 2 h and then introduced into a Petri dish (9 cm diameter) containing the treated leaf disk. The treated leaf disks were replaced daily to ensure fresh food supply and continuous exposure. A moistened cotton ball was placed in the dish to prevent desiccation. Mortality was assessed after 48 h of exposure. Adults were considered dead if they did not move when prodded with a fine brush. The experiment was repeated three times independently. Probit analysis (SPSS version 19.0) was used to calculate the LC_10_ and LC_50_ values along with their 95% confidence intervals.
2.3. Sample Preparation for RNA-Seq
Transcriptomic analyses were performed on three sets of samples from P. striolata: (1) Different developmental stages: eggs, 3rd instar larvae, pupae, and 3-day-old adults. (2) Different tissues of 3-day-old adults: dissected heads, ovaries, midguts, Malpighian tubules, and fat bodies (male/female = 1:1). (3) whole bodies of 3-day-old adults after a 48 h treatment with sublethal doses (LC_10_ and LC_50_) of pyridaben. The LC_10_ dose was employed to simulate a mild stress environment for capturing early responsive genes, while the LC_50_ dose was selected to identify key tolerance genes under high selection pressure, thereby avoiding the physiological collapse associated with higher lethal concentrations. Survivors were collected for RNA extraction. Samples from the developmental stages and tissues included three biological replicates, while the pyridaben-treated groups included two biological replicates, respectively. Each biological replicate comprised 50 individuals (or pooled tissues from 50 individuals), which were immediately frozen in liquid nitrogen and stored at −80 °C until RNA extraction.
2.4. RNA Extraction, Library Construction, and Sequencing
Total RNA was extracted from each sample using TRIzol^®^ Reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s protocol. RNA integrity and concentration were assessed using an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA) and a NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Only samples with an RNA Integrity Number (RIN) > 8.0 were used for subsequent library construction. Sequencing libraries were generated using a NEBNext^®^ Ultra™ RNA Library Prep Kit from Illumina^®^ (New England Biolabs, Ipswich, MA, USA) following the manufacturer’s recommendations. The libraries were sequenced on an Illumina NovaSeq 6000 platform (Illumina, San Diego, CA, USA), generating paired-end reads of approximately 150 bp in length.
2.5. Data Processing and Differential Expression Analysis
Raw sequence reads were processed through a stringent quality control pipeline. Adapter sequences and low-quality reads were removed using Trimmomatic. The clean reads were de novo assembled into transcripts using Trinity software (version 2.8.5). The assembled unigenes were functionally annotated against major databases, including NR, NT, Pfam, KOG/COG, Swiss-Prot, KEGG, and GO. Gene expression levels were estimated by calculating TPM (Transcripts Per Million) values. Differential expression analysis was conducted using the DESeq2 R package (version 1.26.0) with raw read counts as input. Genes with an adjusted p-value (padj) < 0.05 and |log_2_(FoldChange)| > 1 were assigned as differentially expressed genes (DEGs).
2.6. P450 Gene Identification, Phylogeny, and Expression Profiling
Putative P450 genes in P. striolata were identified by performing a local BLAST search against its genome assembly, which was retrieved from NCBI (https://www.ncbi.nlm.nih.gov/datasets/genome/GCA_918026865.1/ (accessed on 1 December 2025)). The search used known insect P450 protein sequences from Tribolium castaneum as queries. Candidate genes were further verified by detecting the conserved P450 domain (PF00067) using InterProScan and the Pfam database. The coding completeness of the candidate P. striolata P450s was assessed before obtaining official designations based on the criteria established by the P450 Nomenclature Committee [19].
The deduced amino acid sequences of the identified P. striolata P450s were aligned with P450s from other model insect species using MAFFT (version 7.1). A phylogenetic tree was reconstructed using the neighborhood-joining method based on FastTree. The robustness of the tree was evaluated with 1000 bootstrap replicates. The resulting tree was visualized and annotated using iTOL (https://itol.embl.de/ (accessed on 20 December 2025)).
To further determine the expression levels of the P450 genes identified in the P. striolata genome, the clean reads were then pseudoaligned to the transcript sequences from the reference genome using Kallisto, and gene expression levels were quantified as TPM (Transcripts Per Million). Heatmaps were generated using the TBtools-II software (version 2.382) to visualize the expression profiles [20].
2.7. qRT-PCR Validation
To validate the transcriptome sequencing results, the expression levels of key P450 genes identified as differentially expressed were analyzed using qRT-PCR. Total RNA was extracted from the whole bodies of surviving adults (dead individuals were excluded) collected after 48 h of exposure (consistent with the bioassay duration) to control and pyridaben-treated (LC_10_ and LC_50_) conditions. First-strand cDNA was synthesized from 1 µg of DNase I-treated total RNA using the PrimeScript RT reagent Kit with gDNA Eraser (Takara Bio Inc., Kusatsu, Shiga, Japan).
Gene-specific primers were designed using Primer Premier 5.0 software, with amplicon lengths ranging from 80 to 200 bp. The β-Actin gene was selected as an internal reference based on its stable expression in the transcriptome data. qRT-PCR was performed in triplicate for each sample using TB Green^®^ Premix Ex Taq™ II (Takara Bio Inc., Kusatsu, Shiga, Japan) on a QuantStudio 5 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). The reaction protocol consisted of an initial denaturation at 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 34 s. A melt curve analysis was performed at the end of each run to confirm the specificity of amplification. The relative expression levels of target genes were calculated using the 2^(−ΔΔCt)^ method.
2.8. RNA Interference and Toxicity Bioassay
Based on their expression profiles, CYP6TH1 and CYP6TH2 were selected for RNA interference (RNAi). Specific double-stranded RNA (dsRNA) primers targeting CYP6TH1, CYP6TH2, and GFP (as a control) were designed using E-RNAi (https://e-rnai.dkfz.de/signaling/e-rnai3/ (accessed on 10 April 2024)), each incorporating a T7 promoter sequence at the 5′ end (see Supplementary Table S3 for primer details). These primers were amplified from P. striolata cDNA samples or from a plasmid harboring the GFP gene. PCR products were confirmed by sequencing and then used as templates for in vitro transcription using the T7 RiboMAX™ Express RNAi System (Promega, Madison, WI, USA), following the manufacturer’s instructions. The synthesized dsRNA (3000–5000 ng/μL) was quantified using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), and its integrity was assessed via agarose gel electrophoresis. dsRNA samples were aliquoted and stored at −80 °C until further use.
Microinjection of dsRNA into insects was performed. Approximately 200 ng of either dsGFP (control) or gene-specific dsRNA was injected into each P. striolata adult. RNAi efficiency was assessed using qPCR at 24, 48, and 72 h post-injection, with cDNA synthesized from isolated total RNA. The impact of dsRNA on target gene expression was analyzed using qPCR with dsGFP-injected insects as the negative control. Separately, adults treated with dsRNA were transferred to fresh B. juncea leaves that had been treated with LC_10_ or LC_50_ doses of pyridaben and allowed to air-dry. Mortality was recorded over a 72 h period. Each RNAi-treated group consisted of 20 adults, and the entire bioassay was independently replicated three times.
2.9. Molecular Docking
The three-dimensional (3D) structures of CYP6TH1 and CYP6TH2 proteins were predicted using AlphaFold3 (https://alphafold.com). The stereochemical quality and structural validity of the predicted models were evaluated using the SAVES v6.1 server (https://saves.mbi.ucla.edu/ (accessed on 15 August 2025)). The 3D structure of pyridaben was obtained from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/ (accessed on 15 August 2025)). The proteins and ligand were first prepared for docking using AutoDock Tools v1.5.4 [21]. Molecular docking simulations were then performed with AutoDockTools-1.5.7 to explore the binding interactions between the proteins (CYP6TH1 and CYP6TH2) and pyridaben. The resulting complexes were visualized and analyzed using PyMOL (version 3.0.3). The interactions between pyridaben and the active sites of CYP6TH1 and CYP6TH2 were visualized using Discovery Studio (version 4.5).
2.10. Statistical Analysis
SPSS version 16.0 (SPSS, Inc., Chicago, IL, USA) was applied to perform statistical analyses. Probit analysis was used to calculate the LC_10_ and LC_50_ values along with their 95% confidence intervals. Student’s t-test was used to evaluate statistically significant differences between two samples. Mortality data among adult insects were analyzed using one-way ANOVA at p < 0.05.
3. Results
3.1. Identification and Phylogenetic Analysis of CYP Genes
Based on the P. striolata genome, 94 full-length PsCYP open reading frames (ORFs) were identified through keyword search and domain analysis (Table S1). The 94 full-length PsCYPs were officially named following the criteria established by the P450 Nomenclature Committee (Dr. D. Nelson, personal communication). According to the standard classification system, these 94 PsCYPs were grouped into four major clans: mitochondrial (6 genes), CYP2 (8 genes), CYP3 (53 genes), and CYP4 (27 genes). These clans were further subdivided into 23 families and 57 subfamilies (Table S2). A neighborhood-joining phylogenetic tree was constructed to infer the orthologous relationships of P450 sequences between T. castaneum and P. striolata (Figure 1). The tree resolved the four major P450 clans: CYP2, CYP3, CYP4, and mitochondrial. The CYP2 and mitochondrial clans comprised highly conserved members with well-established functions, whereas the CYP3 and CYP4 clans showed greater diversification, with many genes having unknown functions.
Among the conserved clans, four genes, three from the mitochondrial clan and one from CYP2, were identified as Halloween genes (CYP302A1, CYP306A1, CYP307A2, CYP314A1, and CYP315A1), which are known to encode enzymes involved in the synthesis of the molting hormone 20-hydroxyecdysone, as previously reported in D. melanogaster [22]. Another gene from the CYP2 clan, CYP18A1, is likely implicated in 20-hydroxyecdysone degradation in insects. Within the CYP3 and CYP4 clans, four and three distinct clusters of P. striolata CYPs were identified, respectively. Members of these clans are typically associated with the metabolism of pesticides, plant secondary metabolites, odorants, and pheromones. The expansion of CYP3 and CYP4 clans in P. striolata may contribute to its high capacity for detoxification and insecticide resistance.
3.2. Tissue and Stage Expression Pattern
To gain insight into the potential functions of CYP genes in P. striolata, we analyzed their expression patterns across different developmental stages and adult tissues using TPM values. Developmental stage expression analysis clustered the 94 CYP genes into distinct clades (Figure 2). Eggs highly expressed a group of 12 genes, including CYP302A1, CYP353A10, and CYP315A1. Another 23 genes, such as CYP6SW18, CYP4US1, CYP349Y2, CYP4SW1, CYP6TH3, and CYP3831A1, showed elevated expression in adult stages. Additionally, 20 genes were predominantly expressed in larvae, while 10 genes exhibited peak expression in pupae.
Tissue-specific expression analysis revealed diverse patterns of CYP gene expression in various tissues (Figure 3). A group of 12 genes showed relatively high expression in the midgut, while another 10 genes were highly expressed in the Malpighian tubules. Thirty-nine genes exhibited elevated expression in the head. Another 5 genes were predominantly expressed in fat bodies. These results reveal that CYP genes are differently expressed in specific tissues of P. striolata, suggesting functional specializations connected to their expression sites. CYPs abundantly expressed in the midgut, Malpighian tubules and fat body are likely involved in the detoxification and metabolism of pesticides or plant secondary chemicals. Those expressed in the head may play roles in neurological or sensory processes.
3.3. Impact of Pyridaben on Insect Survival and P450 Gene Expression
As summarized in Table S4, pyridaben exhibited relatively low insecticidal activity against adult P. striolata. After 24 h of exposure, mortality rates were 1.19 ± 1.19%, 18.75 ± 6.25%, 29.27 ± 8.00%, 43.48 ± 9.32%, and 54.67 ± 5.81% at concentrations of 100, 125, 166.7, 250, and 500 μg/mL, respectively. Based on the mortality data, the probit regression equation was fitted as y = 2.8504x − 2.2559, where y represents the probability of mortality and x denotes the logarithm of concentrations. The LC_50_ and LC_10_ values were 351.2 μg/mL and 180.3 μg/mL, respectively.
The expression levels of CYP genes in adults following 24 h exposure to LC_10_ and LC_50_ pyridaben treatments are shown in Figure S1. Among the identified CYP genes, CYP6TG1, CYP6TH1, CYP6TH2, CYP6TJ2, CYP6TJ3, and CYP4Q62, were upregulated under both doses. Four of them, including CYP6TH1, CYP6TH2, CYP6TJ2, and CYP6TJ3, which are from the same clade, showed more than four-fold upregulation. The upregulation was further validated by RT-qPCR (Figure 4).
3.4. Knockdown of CYP6TH1 and CYP6TH2 Reduces Survival of P. striolata to Pyridaben Treatments
To investigate the role of CYP6TH1 and CYP6TH2 in pyridaben tolerance, RNAi was employed to knock down their expression in P. striolata adults (Figure 5). Compared to the dsGFP-treated controls, dsCYP6TH1 resulted in significant reductions in CYP6TH1 expression, with 79.36%, 65.14%, and 62.92% reduction in transcript levels at 24, 48, and 72 h post dsRNA-injection, respectively (Figure 5A; p < 0.05, Unpaired t-test). dsCYP6TH2 injection also significantly suppressed CYP6TH2 expression, with 70.62%, 66.23%, and 54.71% reduction at the above-mentioned time points (Figure 5B; p < 0.05 Unpaired t-test). Correlated with gene silencing, susceptibility to pyridaben was markedly enhanced: mortality under LC_10_ exposure increased by 12.33% and 16.33% in dsCYP6TH1- and dsCYP6TH2-treated beetles, respectively. Mortality rose even higher following LC_50_ exposure, with 25.00% and 31.67% increases in insect death rates, respectively (Figure 5C).
3.5. Predicted Binding of CYP6TH1 and CYP6TH2 with Pyridaben
To gain more profound insights into the potential interactions between pyridaben and CYP6TH1/CYP6TH2, molecular docking simulations were performed. The results revealed that the binding of pyridaben to CYP6TH1 was primarily stabilized by van der Waals forces and hydrophobic interactions (Figure 6A,B). A total of 16 amino acid residues of CYP6TH1 (Ile83, Arg111, Gly112, Tyr114, Phe126, Leu217, Phe218, Leu229, Val232, Pro236, Pro246, Met247, Phe299, Ile303, Thr374, and Pro395) were found to interact with pyridaben via van der Waals forces. Additionally, 6 residues (Phe60, His88, Leu113, Phe127, Ile233, and Phe480) formed hydrophobic interactions with the insecticide (Figure 6B). Similarly, van der Waals forces and hydrophobic interactions were also identified as the main driving forces for the binding of pyridaben to CYP6TH2 (Figure 6C,D). For instance, 10 residues (Phe60, Ile83, Gly112, Tyr114, Val232, Pro236, Pro246, Phe299, Pro395, and Phe480) participated in van der Waals interactions with pyridaben. Furthermore, the binding free energies of CYP6TH1 and CYP6TH2 with pyridaben were −7.73 kcal/mol and −7.57 kcal/mol, respectively, indicating strong binding affinity between CYP6TH1 and pyridaben and between CYP6TH2 and pyridaben.
4. Discussion
Chemical insecticides remain one of the most direct and effective approaches for pest management. However, insects have developed diverse resistance mechanisms in response to prolonged selection pressure. These include the overexpression of detoxification enzymes and structural mutations that enhance catalytic efficiency through amino acid substitutions [23,24,25]. Key detoxifying enzymes involved in insecticide metabolism are cytochrome P450 monooxygenases (P450s), glutathione S-transferases (GSTs), and carboxylesterases (CarEs) [26]. Among these, P450s are particularly versatile, catalyzing oxidation–reduction reactions critical for numerous physiological and detoxification processes. The advent of high-throughput sequencing has greatly accelerated the identification of P450 genes, including in non-model species lacking reference genomes. To date, P450s have been characterized in many major insect pest species; for instance, 77 P450-encoding genes have been identified in Chilo suppressalis, 58 in Bemisia tabaci MED, 103 in Tenebrio molitor, and 71 in Bactrocera dorsalis [27,28,29,30]. In this study, we identified 94 full-length CYP genes in P. striolata for the first time. These were classified into four clans: CYP2, CYP3, CYP4, and mitochondria. The genes were further subdivided into 23 families and 57 subfamilies. Considerable interspecific variations in CYP gene numbers were observed, consistent with the high evolutionary dynamism of P450-encoding genes in adaptation to xenobiotic metabolism. Particularly, genes in the CYP3 clan undergo frequent gene duplication and loss along with point mutations [31]. Gene duplication and nucleotide substitutions coupled with positive selection led to functional divergence, driving P450-encoding gene diversification. This study identified expansions in specific CYP subfamilies in P. striolata. Four distinct clades were identified within the CYP3 clan. For example, the CYP6TJ and CYP6TH subfamilies formed one clade, while the CYP6BQ, CYP6SW, CYP6TF, CYP6SH, CYP6TG, CYP6TK, and CYP6HD subfamilies constituted another clade. These expanded clades may significantly enhance metabolic resistance to specific insecticides or plant secondary compounds [28,31,32].
Despite the expansion and diversity of some P450 clades, many other CYP genes are evolutionarily conserved across insect species. The conserved genes include the Halloween genes (CYP302A1, CYP306A1, CYP307A2, CYP314A1, and CYP315A1) and the CYP18A1 gene. The conserved P450-encoding genes are usually essential for biosynthesis and inactivation of the molting hormone 20-hydroxyecdysone (20E) [33,34]. In our study, five Halloween genes (CYP302A1, CYP306A1, CYP307A2, CYP314A1, and CYP315A1) and the CYP18A1 gene were identified in P. striolata, suggesting their conserved roles in ecdysteroid regulation. In addition to these well-known conserved P450-encoding genes, three other genes (CYP305A1, CYP49A1, and CYP353A1) were also found highly conserved. Of these, CYP305A1 and CYP49A1 have been reported to play a role in insect resistance to chlorantraniliprole, possibly through enhanced detoxification [35].
Spatiotemporal expression profiling revealed that most P450-encoding genes in P. striolata, particularly those in the CYP3 and CYP4 clans, were highly expressed in adults. This adult-biased expression may reflect increased metabolic demands and direct exposure to insecticides, necessitating robust detoxification capacity. In contrast, earlier developmental stages (larvae and pupae) are more sheltered within the soil matrix, reducing exposure to xenobiotics [36]. Halloween genes, which regulate ecdysteroid synthesis, were predominantly expressed in egg, larval, or pupal stages—a pattern consistent with other insects. For example, in B. dorsalis, CYP302A1 is highly expressed in eggs and contributes to embryonic cuticle formation [37]. Tissue-specific expression also aligned with physiological roles: the midgut, a primary site for digestion and xenobiotic defense [38,39], showed specific expression of CYP49A1 in Spodoptera frugiperda, where its upregulation correlates with chlorantraniliprole resistance [35]. Three Halloween genes, including CYP314A1, CYP315A1, and CYP306A1, are highly expressed in the ovary, consistent with their role in adult ecdysteroid synthesis following the degeneration of the prothoracic gland [40].
Upregulation of detoxification genes in response to xenobiotics is a well-documented resistance mechanism [41,42,43,44,45]. For example, in Helicoverpa armigera, CYP9A14 and CYP6AE11 are induced by multiple insecticides and contribute to resistance [42]. Similarly, in Bradysia odoriphaga, lambda-cyhalothrin and imidacloprid induce CYP6QE1 and CYP6FV21 [46], and in Locusta migratoria, CYP6FE1, CYP6FF1, and CYP6FG1 are associated with deltamethrin detoxification and resistance [47]. In this study, exposure of P. striolata to pyridaben resulted in significant upregulation of six P450-encoding genes, five of which belong to the CYP3 clan. To date, most P450s known to metabolize pesticides and xenobiotics have been identified within the CYP6 and CYP9 families of the CYP3 clan. Mounting evidence indicates that CYP6 family genes play important roles in insecticide resistance. For instance, CYP6EM1 confers thiamethoxam resistance in B. tabaci [48], CYP6CX4 contributes to flupyradifurone resistance [49], and CYP6AQ83 enhances flonicamid tolerance in Solenopsis invicta [50]. Here, RNA interference targeting either CYP6TH1 or CYP6TH2 increased P. striolata susceptibility to pyridaben, suggesting their roles in pyridaben resistance. Although RNAi confirmed the involvement of CYP6TH1 and CYP6TH2 in pyridaben tolerance, they are likely not the sole contributors. Our data showed that other P450 genes, such as CYP6TG1, CYP6TJ2, and CYP6TJ3, were also upregulated upon exposure. Insecticide resistance is typically a complex trait involving functional redundancy or synergism among multiple enzymes. Therefore, the observed tolerance is likely the collective result of a broader P450 network. Molecular docking has been widely used to predict the binding affinity between detoxification enzymes and xenobiotics, especially in insecticide resistance studies [51,52,53]. Our docking results showed CYP6TH1 and CYP6TH2 have a strong binding affinity for pyridaben.
5. Conclusions
In summary, this study identified 94 full-length cytochrome P450-encoding genes in P. striolata. We characterized their spatiotemporal expression patterns and demonstrated that pyridaben upregulates CYP6TH1 and CYP6TH2. RNAi assays provide initial evidence that CYP6TH1 and CYP6TH2 contribute to pyridaben tolerance. Molecular docking suggests stable binding of pyridaben to both CYP6TH1 and CYP6TH2 proteins. These findings underscore the importance of CYP6 subfamilies in insecticide tolerance and mark the first investigation into the role of the CYP6TH subfamily in pyridaben tolerance, as well as the first mechanistic insights into pyridaben tolerance in P. striolata.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Li Z. Costamagna A.C. Beran F. You M. Biology, ecology, and management of flea beetles in Brassica Crops Annu. Rev. Entomol.20246919921710.1146/annurev-ento-033023-01575338270984 · doi ↗ · pubmed ↗
- 2Tengfei X. Nanda S. Fengliang J. Qingsheng L. Xia F. Control efficiency and mechanism of spinetoram seed-pelleting against the striped flea beetle Phyllotreta striolata Sci. Rep.202212952410.1038/s 41598-022-13325-835680974 PMC 9184531 · doi ↗ · pubmed ↗
- 3Li J. Qiu L. Wang H. Fu J. Chemicals used for controlling Phyllotreta striolata (F.)Fujian J. Agric. Sci.200711518
- 4Liang M. Huang D. Field Trials of Several Pesticides against Phyllotreta striolata J. Anhui Agric. Sci.201713149150
- 5Wei J.L. Li X.L. Li J.J. Wang X.M. Control effect of five pesticides against Rehamanniae flea beetle and their influence on natural enemies Hunan Agric. Sci.20083108110
- 6Zheng Y. Liu X. Jiang L. Qiao K. Zuo Y. Wang K. Toxicity of clothianidin and other insecticides to Phyllotreta striolata Chin. J. Pestic. Sci.20152230234
- 7Khalighi M. Dermauw W. Wybouw N. Bajda S. Osakabe M. Tirry L. Thomas V.L. Molecular analysis of cyenopyrafen resistance in the two-spotted spider mite Tetranychus urticae Pest Manag. Sci.20157210311210.1002/ps.407126118668 · doi ↗ · pubmed ↗
- 8Dong B. Hu J. Residue levels and risk assessment of acetamiprid–pyridaben mixtures in cabbage under various open field conditions Biomed. Chromatogr.202337 e 572810.1002/bmc.572837700621 · doi ↗ · pubmed ↗