Caste-Dependent Interspecific Tolerance Permits Alien Reproductives to Reproduce Within Host Colonies in Reticulitermes Termites Under Laboratory Conditions
Zhuang-Dong Bai, Ya-Nan Dong, David Sillam-Dussès, Rui-Wu Wang

TL;DR
This study shows that termite colonies can be infiltrated by foreign queens and kings, who are tolerated and allowed to reproduce, revealing a loophole in their defense system.
Contribution
The study demonstrates caste-dependent interspecific tolerance in termites, enabling alien reproductives to reproduce within host colonies.
Findings
Host termites killed intruder workers but tolerated intruder queens and kings.
Alien reproductives were fed and cared for by host workers, leading to successful reproduction.
Molecular testing confirmed the offspring were genetically from the intruding R. labralis pair.
Abstract
Termite colonies are known as “closed fortresses” that fiercely attack any intruders to protect their nest. However, it is unclear whether this defense is always perfect. In this study, we tested whether termites from one species (Reticulitermes labralis) could successfully infiltrate and live within the colony of a different species (Reticulitermes aculabialis). We introduced workers, queens, and kings of R. labralis into orphaned groups of R. aculabialis in the laboratory. We found that the host termites immediately killed the intruder workers, but surprisingly tolerated the intruder queens and kings. These surviving royal intruders were fed by the host workers and successfully produced their own babies. We used molecular testing to prove that the new babies were indeed the offspring of the intruders. This study shows that termite colony defense has a “loophole”: while they recognize…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2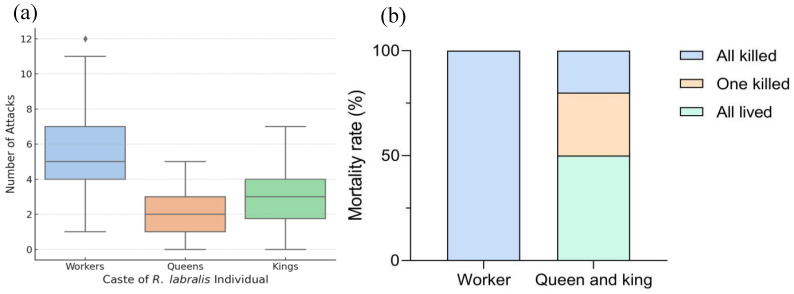 Figure 3
Figure 3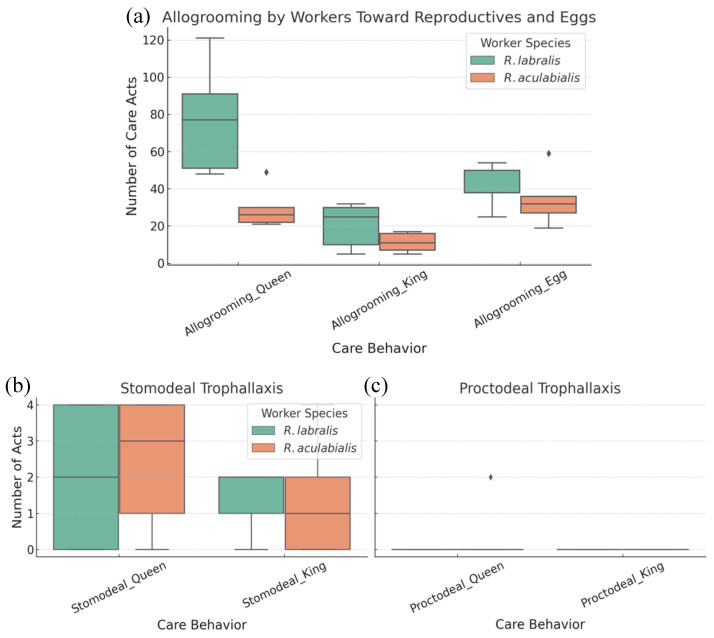 Figure 4
Figure 4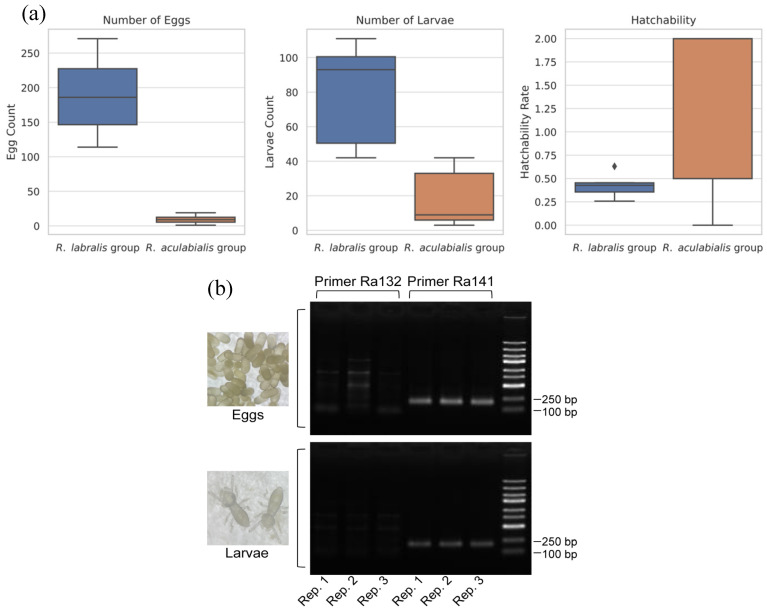 Figure 5
Figure 5- —China Postdoctoral Science Foundation
- —National Natural Science Foundation of China-Yunnan Joint Fund
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect and Arachnid Ecology and Behavior · Plant and animal studies · Fossil Insects in Amber
1. Introduction
Nestmate recognition constitutes the primary immune defense of eusocial insects, allowing colonies to protect their resources and brood from competitors and predators [1,2]. In termites, this recognition system is largely mediated by cuticular hydrocarbons (CHCs) and environmental cues, typically resulting in immediate and often lethal aggression toward any non-nestmate, especially those of different species [3,4]. Consequently, widely accepted dogma suggests that termite colonies are closed societies with strict species boundaries [5]. However, the absolute rigidity of these boundaries has been challenged by observations of colony fusion in lower termites and the existence of obligate inquilines in higher termites [6,7,8]. These phenomena raise a fundamental question: Is the termite recognition system sufficiently flexible to permit the infiltration of heterospecific individuals under specific conditions?
While social parasitism, where one species exploits the social care of another, is highly diverse in the Hymenoptera (ants, bees, and wasps) [9,10,11,12], it is virtually undescribed in termites. Theoretical explanations for this disparity often cite the hemimetabolous development of termites and the lifetime presence of the royal pair, which theoretically leaves little niche space for parasitic intruders [13,14]. Nevertheless, early reviews and laboratory assays have noted instances where heterospecific termites, particularly reproductives, survived initial encounters with alien colonies [3]. Despite these intriguing historical observations, it remains unknown whether such surviving “intruders” can physiologically integrate into the host colony, exploit the host’s labor for brood care, and successfully produce viable offspring. Determining this reproductive potential is crucial for understanding whether the behavioral precursors for social parasitism exist in termites.
A potential driver for such integration is phylogenetic proximity. “Emery’s rule” posits that social parasites are often closely related to their hosts, as shared ancestry implies similar chemical profiles and communication signals, facilitating deception or tolerance [15,16]. In termites, while colony-specific odors vary, certain “royal” recognition signals (e.g., queen pheromones) appear to be evolutionarily conserved across species [17,18]. This suggests a hypothesis of caste-dependent tolerance: while host workers may aggressively reject alien workers due to divergent colony odors, they might exhibit reduced aggression toward alien reproductives that bear conserved status signals.
To test the permeability of species boundaries and the hypothesis of caste-dependent tolerance, we examined interactions between two subterranean termites, Reticulitermes labralis and R. aculabialis (Figure 1). These species overlap in distribution in central China and share similar nesting ecology, yet they possess distinct morphological and developmental traits (Figure 2, Figures S1 and S2). We established a controlled laboratory model to simulate the intrusion of R. labralis individuals (workers, queens, and kings) into orphaned R. aculabialis groups. Unlike previous studies that primarily focused on behavioral assays or short-term survival, we monitored the long-term reproductive outcomes of these mixed associations. We utilized species-specific microsatellite markers to verify the parentage of the resulting brood. This study provides the confirmed evidence that termite reproductives can not only survive within a heterospecific colony, but also successfully exploit host workers to rear their own offspring, revealing a latent potential for parasitic-like strategies in the genus Reticulitermes.
2. Materials and Methods
2.1. Termite Collection and Species Identification
Field colonies of Reticulitermes aculabialis and R. labralis were collected from decaying logs at two distinct geographic locations in China: Purple Mountain, Nanjing (Jiangsu Province; 118°53′ E, 32°03′ N) and Guifeng Mountain, Xi’an (Shaanxi Province; 108°46′ E, 34°00′ N). A total of five independent colonies per species were collected to ensure genetic diversity. Species identity was confirmed using a dual approach: morphological examination of soldier castes (labrum shape) and mitochondrial cytochrome oxidase subunit II (COII) gene sequencing (Supplementary Methods and Table S1). While workers of both species are morphologically similar, they can be distinguished by significant differences in body size and head width (see Figure S1), which were used to identify individuals during behavioral observations. Stock colonies were maintained in the laboratory in plastic containers provided with moist pine wood at 25 °C and 70% relative humidity under a 14L:10D photoperiod before experimental setup.
2.2. Experimental Setup and Neotenic Differentiation
To establish standardized experimental units, we separated termites from stock logs. For each replicate, 200 individuals (192 workers and 8 soldiers) were housed in a 9-cm Petri dish lined with moist filter paper. This group size was chosen to mimic a functional social unit while allowing for precise observation. A total of 40 replicate groups were established for each species. Groups were maintained under the controlled conditions described above, with water and filter paper replenished weekly. Within one month of isolation, neotenic reproductives (secondary queens and kings) differentiated from nymphs or workers in the R. labralis groups. To standardize the reproductive status of ‘donor’ groups, we removed excess reproductives, leaving exactly one neotenic queen and one neotenic king per R. labralis dish. Conversely, for R. aculabialis groups designated as ‘recipients’ (hosts), any differentiated reproductives were strictly removed to create an orphaned, reproductive-less state.
2.3. Interspecific Introduction Assays
We designed an introduction assay to simulate the intrusion of alien reproductives into an orphaned colony. The experimental design consisted of three treatments: 1. Mixed Group (Experimental): One R. labralis worker, one queen, and one king were introduced into an orphaned R. aculabialis host group (n = 20). To minimize handling stress and physical injury, introduced individuals were gently transferred using soft entomological forceps into the center of the recipient dish. 2. Host Control (Negative Control): Orphaned R. aculabialis groups maintained without any introduction to monitor baseline survival and verify the absence of spontaneous differentiation (n = 20). 3. Conspecific Control (Positive Control): R. labralis colonies maintained with their own native neotenic pair to provide baseline data on reproductive output (n = 20).
2.4. Behavioral Observations and Survival Monitoring
Aggression Assays: Immediately following introduction, interactions were video-recorded using a Sony AX100E camera (Sony Corporation, Tokyo, Japan). Aggressive behavior was quantified by counting the cumulative number of biting attacks directed by host workers toward each introduced individual (worker, queen, or king) during the first 2 min of contact. Long-term Survival: Mortality of the introduced individuals was recorded at 3 h, 24 h, and daily for the first week. Once the groups stabilized (cessation of aggression), monitoring continued weekly until day 140.
Social Care Quantification: To assess social integration, we analyzed caregiving behaviors in stabilized Mixed Groups (n = 5) and Conspecific Controls (n = 5). Three 30-min video sessions were recorded for each group. We quantified the frequency of allogrooming (worker toward royal/egg) and trophallaxis (stomodeal and proctodeal; Figure S3).
2.5. Reproductive Assessment and Parentage Verification
The presence of eggs and larvae was monitored weekly starting from day 29 until day 140. Hatchability was calculated as the ratio of hatched larvae to the total number of eggs produced in each group. To rigorously verify that the brood produced in Mixed Groups originated from the introduced R. labralis parents and not from the host R. aculabialis workers (via parthenogenesis or undetected neotenics), we performed molecular parentage analysis. Genomic DNA was extracted from workers, eggs (n = 20) and larvae (n = 3) using a phenol-free extraction kit (Tiangen Biotech, Beijing, China). We screened six microsatellite loci (Rs 03, Rs 76, Rs 78, Ra 132, Ra 141, Ra 144; see Table S2) developed for Reticulitermes [19,20]. Among these, primers Ra132 and Ra141 were identified as diagnostic markers: Ra132 amplifies a ~250 bp fragment exclusively in R. aculabialis, while Ra 141 amplifies a ~250 bp fragment in both species (Figure S4). PCR amplification was conducted as follows: 94 °C for 5 min; followed by 30 cycles of 94 °C for 30 s, 52 °C for 30 s, and 72 °C for 30 s; and a final extension at 72 °C for 10 min. Products were visualized on 1% agarose gels to determine species origin.
2.6. Statistical Analysis
To account for potential non-independence among replicates originating from the same colony, we fitted linear mixed-effects models (LMMs) using the lme4 and lmerTest packages in R, with colony identity included as a random effect. Differences in aggression toward introduced R. labralis castes (worker, queen, king) were analyzed with LMMs including caste as a fixed effect. Model syntax: lmer (attack_count ~ caste + (1|colony_ID), data = data), while mortality was analyzed using Fisher’s exact tests where data distributions did not allow for parametric modeling. Frequencies of grooming and trophallaxis were compared between host (Mixed Group) and conspecific (Control) workers using LMMs, using species identity (host vs. conspecific workers) as a fixed factor and colony ID as a random factor. Egg/larval counts and hatchability were compared between Mixed Group and Control Group using similar mixed models, while hatchability rates were also analyzed using Fisher’s exact tests where data distributions did not allow for parametric modeling. All tests were two-tailed with a significance threshold of α = 0.05. All statistical analyses and data visualization were conducted in R version 4.3.2.
3. Results
3.1. Caste-Dependent Aggression and Survival of Introduced Termites
Upon introduction into orphaned R. aculabialis groups, host workers exhibited immediate and highly discriminatory aggression based on the caste of the R. labralis intruders. The frequency of biting attacks directed toward introduced workers was significantly higher than that directed toward introduced queens (LMM: z = 10.17, p < 0.001) or kings (z = 7.28, p < 0.001). This selective aggression resulted in distinct mortality outcomes: 100% of introduced R. labralis workers were killed within 24 h of introduction. In contrast, the alien reproductives exhibited significantly higher survival rates (Fisher’s exact test, p < 0.001; Figure 3). Specifically, in 50% of the experimental groups, both the alien queen and king survived the initial aggression and persisted for the entire 140-day observation period. In 30% of groups, one of the pair survived, while both were eliminated in only 20% of cases. These results indicate that, while species recognition cues trigger lethal aggression against foreign workers, these barriers are relaxed toward foreign reproductives. In addition, our observations found that the R. labralis workers died quickly after the attacks began, while it took more time for the reproductives to succumb (Table S3).
3.2. Behavioral Integration and Social Care Toward Alien Reproductives
Following the stabilization of the mixed groups (typically after one week), surviving R. labralis reproductives were behaviorally integrated into the R. aculabialis host groups. Host workers were observed performing essential caregiving behaviors, including allogrooming and trophallaxis (both stomodeal and proctodeal), toward the alien royals. The quality of care, however, varied by behavior type. The frequency of allogrooming performed by host workers toward alien queens (z = −4.85, p < 0.001) and kings (z = −3.87, p < 0.001) was significantly lower than that observed in conspecific control groups. Similarly, alien eggs received significantly less grooming from host workers compared to conspecific eggs (z = −3.51, p < 0.001). Crucially, however, nutritional integration appeared successful: the frequency of stomodeal and proctodeal trophallaxis provided by host workers to the alien queen and king did not differ significantly from that observed in conspecific R. labralis groups (p > 0.05 for all comparisons; Figure 4). This indicates that, while hygienic behavior (grooming) remains constrained by species barriers, the nutritional transfer system is permeable to support alien reproductives. Furthermore, host workers were observed actively relocating alien eggs and larvae to safety piles upon disturbance, indicating behavioral acceptance of the brood (e.g., by opening the lid of the Petri dish; Videos S1 and S2).
3.3. Reproductive Success and Parentage Verification of the Brood
Physiological integration culminated in successful reproduction. In mixed groups where the royal pair survived, egg laying commenced on day 29, with larvae first observed on day 114. By day 140, brood was present in experimental groups, confirming that alien reproductives could exploit host resources to produce offspring. Quantitatively, the reproductive output in mixed groups was lower than in optimized conspecific controls. The total number of eggs (z = −23.32, p < 0.001) and larvae (z = −5.45, p < 0.001) was significantly reduced compared to pure R. labralis colonies. However, the hatchability of eggs in mixed groups was slightly higher than in controls (z = 2.38, p = 0.017; Figure 5a), suggesting that the surviving brood received effective developmental care from host workers.
To rule out the possibility of parthenogenesis by host workers or the presence of undetected host neotenics, we determined the genetic origin of the brood using species-specific microsatellite markers. The diagnostic primer Ra 132 amplifies a ~250 bp fragment in R. aculabialis but yields no product in R. labralis, while primer Ra141 amplifies a fragment in both species (Figure S5). PCR analysis of eggs and larvae collected from the mixed experimental groups showed consistent amplification with the universal primer Ra 141 but, crucially, no amplification with the host-specific primer Ra 132 (Figure 5b). This molecular profile matches that of the introduced R. labralis parents and differs from the R. aculabialis host profile. These results definitively confirm that the brood reared by R. aculabialis workers were the biological offspring of the introduced R. labralis reproductives.
4. Discussion
Our results demonstrate that the species boundaries in Reticulitermes are not absolute, but are instead conditionally permeable. The striking finding is the sharp contrast in host aggression: while R. labralis workers were immediately recognized and eliminated by R. aculabialis hosts, alien queens and kings were frequently tolerated and integrated. This pattern suggests that colony recognition cues are caste-specific [21,22]. In termites, worker recognition relies heavily on cuticular hydrocarbons (CHCs), which are species- and colony-specific [1]. The lethal aggression toward alien workers confirms that R. aculabialis maintains a functional “reject-foreign” system. However, the acceptance of alien reproductives implies that they may lack these specific rejection cues or, more likely, possess “royal recognition signals” that override species identity. Queen pheromones are known to be evolutionarily conserved across termite species [17]. We hypothesize that R. labralis reproductives exploit this sensory bias, a “Royal Trojan Horse” mechanism [23], where conserved royal signals suppress host aggression, allowing them to bypass the guard of R. aculabialis workers. This phenomenon aligns with “Emery’s rule”, where close phylogenetic relatedness facilitates social integration through shared chemical communication channels [16].
Furthermore, the survival of alien reproductives in our experiments may not be attributed solely to the host workers’ recognition of conserved royal pheromones; it likely also depends on the reproductives’ behavioral passivity. Similar “peaceful infiltration” strategies are characteristic of inquiline ants, whose queens exhibit submissive postures and reduced mobility to minimize host aggression [24]. In termites, such behavioral quiescence of reproductives may suppress the host’s recognition, triggering cues or delaying aggressive responses. This aligns with the hypothesis that early-stage parasites evolve appeasement behaviors rather than overt mimicry, behaviors that allow temporary coexistence and reduce the cost of direct conflict [25,26]. Our observations further revealed that host workers provided trophallaxis and allogrooming to alien reproductives, albeit at reduced levels compared to conspecifics. These behaviors are key components of social immunity and recognition maintenance [27,28,29]. Their occurrence toward heterospecific reproductives suggests a degree of behavioral generalization, an inherent flexibility in the social circuitry of termite workers that may inadvertently facilitate parasitism.
A key criticism of laboratory introduction assays is their lack of ecological realism, particularly given that interspecific social parasitism has not yet been documented in termites in the wild. However, our findings offer a plausible mechanism for this apparent absence. The transient nature of the association observed in our study indicates that such interactions may be ephemeral. During swarming events, alates may occasionally encounter foreign colonies that experience royal turnover, although the frequency is low. Under such conditions, the use of conserved royal recognition signals, together with behavioral passivity, may facilitate their integration into the colonies. Because host workers are short-lived, such associations could, in principle, lead to a gradual demographic shift toward offspring produced by the invading reproductives. This process may help explain why mixed-species phases are difficult to detect in natural populations. Moreover, the value of this study lies not in documenting a common field phenomenon but in revealing the physiological and behavioral capacities of the termite social system. Our results show that biological barriers (such as recognition-mediated aggression) are not the primary factors preventing parasitism between these species. When these ecological barriers are breached, as simulated in our experiment, the social barriers (recognition) are surprisingly fragile [30]. This distinction between ecological constraints and physiological capacity is vital for understanding how social parasites might evolve if environmental shifts force species into closer contact [31].
The incipient parasitism observed in Reticulitermes, where foreign reproductives exploit a stressed or orphaned host colony, suggests a potential parallel with the hypothetical early stages of inquilinism evolution (social parasitism without worker caste) [32]. In ants, temporary social parasitism is often a stepping stone to obligate inquilinism [33,34,35]. Our findings suggest a similar evolutionary potential exists in termites. If a mutation or environmental change aligned the swarming schedules of these species, R. labralis queens landing near orphaned R. aculabialis foraging groups could theoretically infiltrate and parasitize them [36].
5. Conclusions
In conclusion, our results show that the biological barriers between two sympatric Reticulitermes species are conditionally permeable. We demonstrate a distinct pattern of caste-dependent interspecific tolerance: while the recognition system strictly eliminates alien workers, it fails to reject alien reproductives. This vulnerability is likely driven by a “Royal Trojan Horse” mechanism, where conserved royal pheromones and behavioral passivity allow intruders to bypass host aggression. Crucially, the successful production of hybrid-reared offspring confirms that the physiological capacity for social parasitism–like exploitation exists in termites, even if it is currently suppressed by ecological constraints in nature. These findings challenge the traditional view of termite colonies as closed societies and suggest that the evolutionary precursors for social parasitism, specifically, the exploitation of host labor by close relatives (Emery’s rule), are latent within the termite lineage.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Clément J.-L. Bagnères A.-G. Nestmate recognition in termites Pheromone Communication in Social Insects CRC Press Boca Raton, FL, USA 2019126155
- 2van Zweden J.S. d’Ettorre P. Nestmate recognition in social insects and the role of hydrocarbons Insect Hydrocarbons: Biology, Biochemistry and Chemical Ecology Cambridge University Press Cambridge, UK 2010 Volume 11222243
- 3Thorne B. Haverty M. Review of inter colony, intraspecific, and interspecific agonism in termites Sociobiology 199119115145
- 4Kaib M. Jmhasly P. Wilfert L. Durka W. Franke S. Francke W. Leuthold R.H. Brandl R. Cuticular hydrocarbons and aggression in the termite Macrotermes subhyalinus J. Chem. Ecol.20043036538510.1023/B:JOEC.0000017983.89279.c 515112730 · doi ↗ · pubmed ↗
- 5Eggleton P. An introduction to termites: Biology, taxonomy and functional morphology Biology of Termites: A Modern Synthesis Springer Berlin/Heidelberg, Germany 2010126
- 6Deheer C.J. Vargo E.L. Strong mitochondrial DNA similarity but low relatedness at microsatellite loci among families within fused colonies of the termite Reticulitermes flavipes Insectes Sociaux 20085519019910.1007/s 00040-008-0999-0 · doi ↗
- 7Cristaldo P.F. De Souza O. Krasulova J. JirošováA. KutalováK. Lima E.R. Šobotník J. Sillam-Dusses D. Mutual use of trail-following chemical cues by a termite host and its inquiline P Lo S ONE 20149 e 8531510.1371/journal.pone.008531524465533 PMC 3897442 · doi ↗ · pubmed ↗
- 8Perdereau E. Bagnères A.-G. Dupont S. Dedeine F. High occurrence of colony fusion in a European population of the American termite Reticulitermes flavipes Insectes Sociaux 20105739340210.1007/s 00040-010-0096-z · doi ↗