The Courtship Behavior and the Ultrastructure of Sex Pheromone Glands in the Hind Tibiae of Male Ghost Moth Endoclita davidi (Lepidoptera: Hepialidae)
Xingrui Huang, Shan Chen, Xing Li, Zihao Zhou, Qiong Zhou

TL;DR
This study explores the courtship behavior and structure of pheromone glands in male ghost moths to understand how they communicate during mating.
Contribution
The study identifies the hind tibiae as the site of pheromone glands in male E. davidi and describes their ultrastructure.
Findings
Courtship behavior in E. davidi occurs during the scotophase and involves wing vibrations and hairpencil unfolding.
Pheromone glands in the hind tibiae connect to scent scales via glandular ducts and microvilli.
Glandular cells contain mitochondria, endoplasmic reticulum, and lipid droplets, indicating active pheromone production.
Abstract
Endoclita davidi bores into and feeds on the stems and roots of Clerodendrum cyrtophyllum. The larvae, when parasitized by the fungus Ophiocordyceps xuefengensis, form valuable Xuefeng Cordyceps. To understand the mechanisms of E. davidi’s reproductive behavior, we studied its courtship behavior and sex pheromone glands. Infrared videos showed that the courtship behavior of E. davidi occurred during the scotophase, during which males vibrate their wings and unfold the hairpencils located on the hind tibiae (the third segment of the hind legs). The bottom of the pits in the bulging apical region of the hairpencils connects to the tibial glands; the epidermal cells of the hind tibiae develop into gland cells with dense microvilli. We confirmed that the male E. davidi’s pheromone glands are located in the hind tibiae and release pheromones via scent scales. Research on the ultrastructure…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
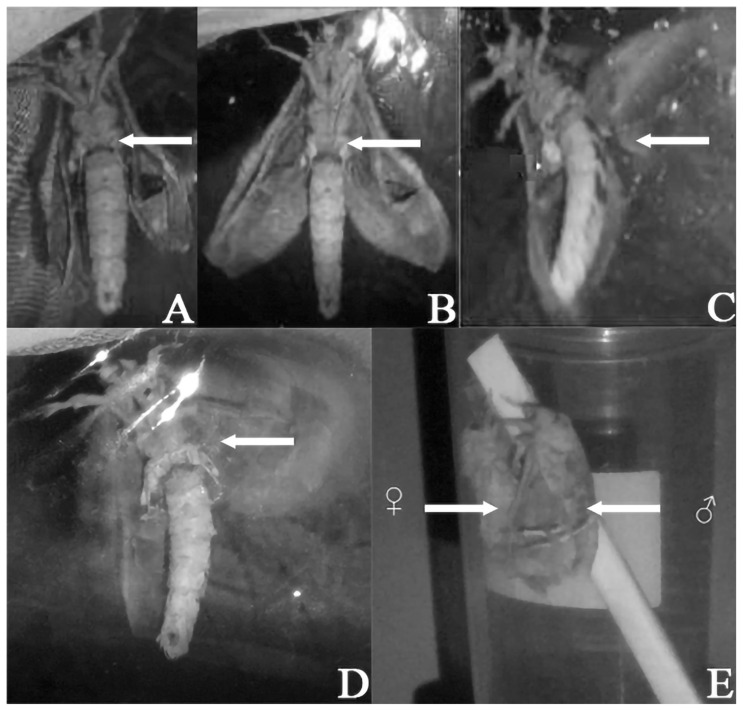 Figure 1
Figure 1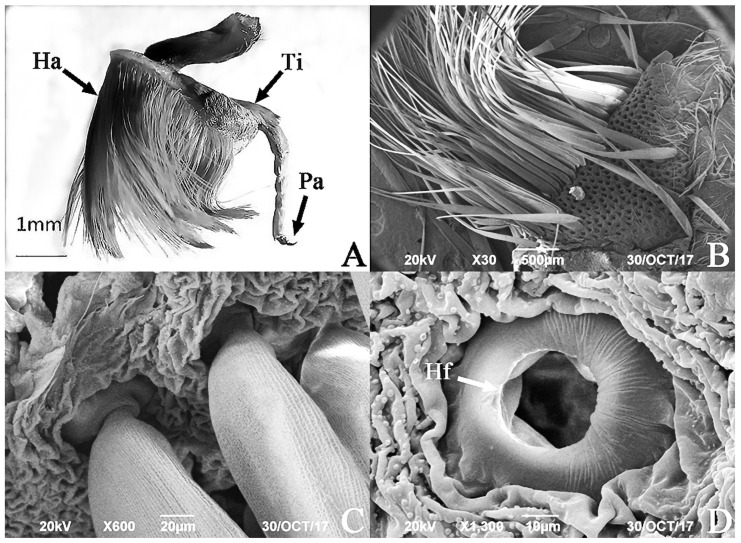 Figure 2
Figure 2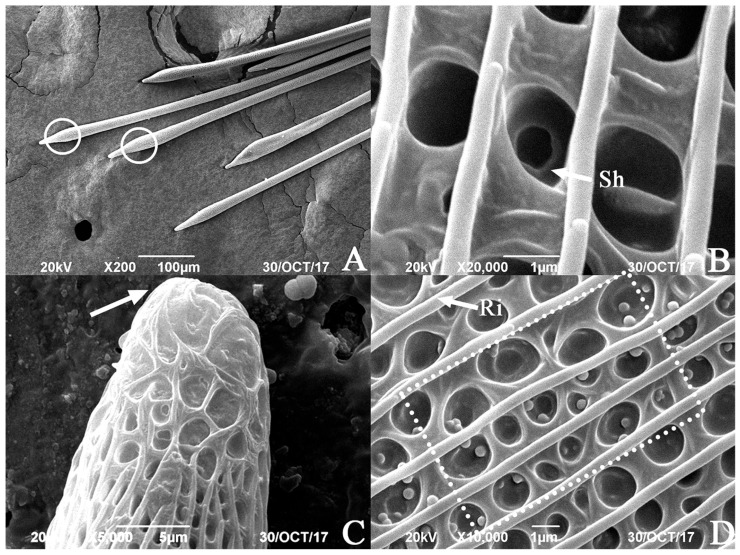 Figure 3
Figure 3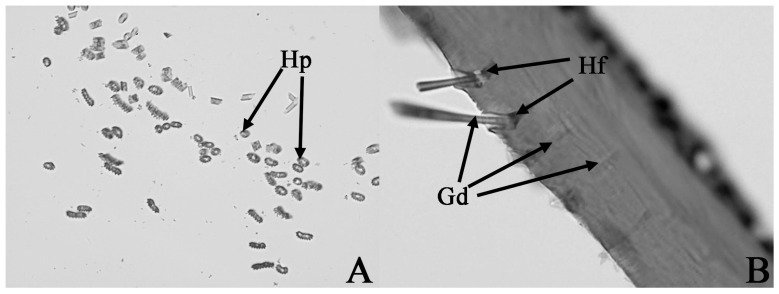 Figure 4
Figure 4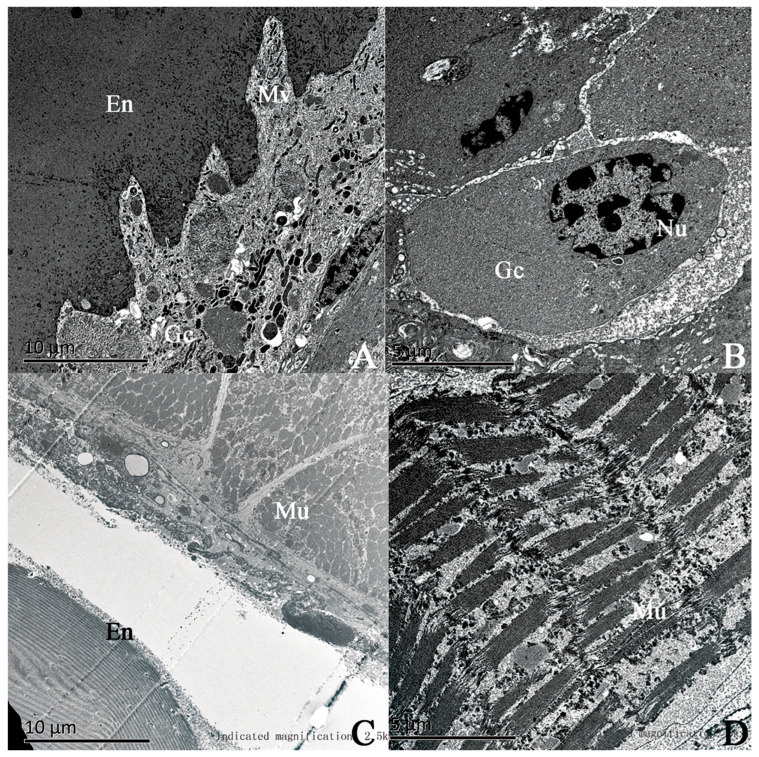 Figure 5
Figure 5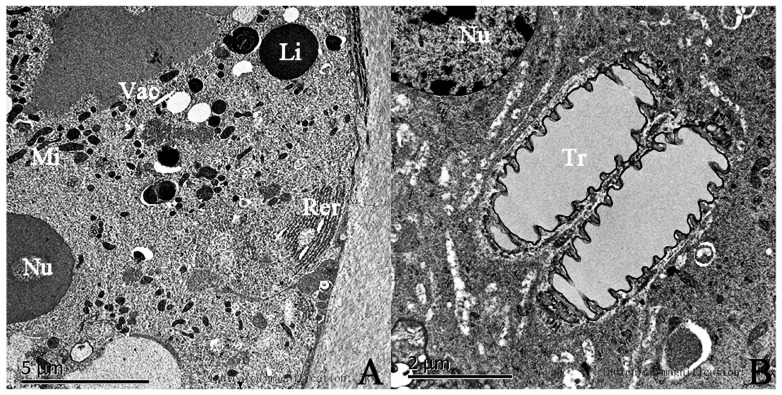 Figure 6
Figure 6- —the Postgraduate Joint Training Base project in Hunan Province
- —Changsha Natural Science Foundation Project
- —the Major Science and Technology Projects of the Hunan Provincial Department of Science and Technology
- —the Natural Science Foundation of Hunan Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect Pheromone Research and Control · Neurobiology and Insect Physiology Research · Lepidoptera: Biology and Taxonomy
1. Introduction
Endoclita davidi (Synonym Phassue giganodus), primarily a stem and root borer of Clerodendrum cyrtophyllum (Tubiflorae: Verbenaceae), is mainly distributed in Guangxi, Fujian, and Hunan Provinces in China [1,2,3]. The larvae of E. davidi are infected by Ophiocordyceps xuefengensis, an entomogenous fungus belonging to Ascomycota, leading to the formation of Xuefeng Cordyceps. This rare Chinese herbal medicine exhibits pharmacological activities analogous to those of Cordyceps sinensis [4,5,6]. Currently, research on E. davidi primarily focuses on the molecular identification of different insect forms [7], the characterization of antennal sensillum types [8], the nutritional composition analysis of larvae [9], and investigations into the reproductive behavioral rhythm of adults [3].
The courtship behavior in insects is a crucial aspect of the reproductive process, typically involving a sequence of complex visual, auditory, and chemical signal exchanges. Among these signals, sex pheromones serve as key chemical mediators in insect courtship [10]. Sex pheromones are chemical substances secreted by specialized glands on the body surface and released into the external environment to elicit courtship behavior among heterosexual conspecifics [11,12]. The location and characteristics of these secretory glands differ among insect species. Lepidopteran insects have been extensively studied with respect to sex pheromones, with research focusing on the morphological characteristics of sex pheromone glands in Noctuidae [13,14,15,16], Pyralidae [17,18,19], and Geometridae [20,21]. For moths, sex pheromone glands are primarily located in the following regions:
(1) The eversible bladder type, composed of specialized glandular epithelial cells in the intersegmental membrane between the eighth and ninth abdominal segments at the posterior end of the female abdomen. Representative species include Helicoverpa zea [14] and Trichoplusia ni [13] (Noctuidae), Conopomorpha sinensis (Gracillariidae) [22], Scopula subpunctaria (Geometridae) [23], and the female moths of Hepialus deainensis (Hepialidae) [24].
(2) The thoracic gland type, located on the dorsal or ventral thoracic surface. In Psychidae, three saddle-shaped glandular areas are present on the dorsal thorax. Additionally, the pheromone gland of female Clania variegata (Psychidae) is situated on the thoracic tergite [25], while in male Ceromitia chalcocapna (Adelidae), it is located on the posterior thoracic sternum [26].
(3) The specialized scale/pouch-like structure type, distributed on the wing margins, abdominal apex, or leg attachment sites. Representative examples include the hairpencils near the proximal margin of the costal fold on the forewings of male Tortricidae and Pyralidae moths [27], the wing-associated glands of male Ephestia elutella (Pyralidae) [28], the abdominal brushes at the abdominal apex of male Grapholita molesta (Tortricidae) [29], the tarsal segments of male Helicoverpa zea, Helicoverpa armigera, H. assulta, and Heliothis virescens (Noctuidae) [30], as well as the hind tibial brushes of male Hepialus humuli [31], Hepialus hecta [32], and Endoclita signifer (Hepialidae) [33]. However, the ultrastructural characteristics of the sex pheromone gland in E. davidi have not been reported to date. Building on our previous understanding of the reproductive behavioral rhythm of E. davidi [3], we further investigated the courtship behavior process of this moth and the ultrastructural features of the male sex pheromone gland. This study aims to provide a foundation for the accurate extraction, isolation, identification, and utilization of E. davidi sex pheromone, as well as to clarify the mechanism underlying its sexual chemical communication.
2. Materials and Methods
2.1. Insect Source and Rearing Conditions
The pupae and larvae of E. davidi were collected in September 2016, 2017, and 2019 from Dongkou County, Shaoyang City, Hunan Province (27°48′ N/110°30′ E, altitude 300–600 m), and then transported to the laboratory in the Life Science College of Hunan Normal University, China. The larvae were individually reared in tissue culture flasks (8.5 cm height; 6 cm diameter) with artificial feed (Table S1) within the Artificial Climate Chamber (RXZ-280D-LED; Ningbo Jiangnan Instrument Factory, Ningbo, China). The Chamber was maintained at 19–22 °C, 80 ± 2% RH with complete darkness. After pupa eclosion, healthy female and male moths with normal wing expansion were selected for testing.
2.2. Observation of Courtship Behavior
The female and male moths on the day of eclosion were collected and caged (30 cm × 30 cm × 30 cm) together with male and female in pairs, with 22 cages in total. The lab was kept at 25 ± 1 °C and 60 ± 5% relative humidity. The behavior of adults in cages was captured using an infrared camera (Rich HD-A210; Shenzhen Lenyin Technology Company, Shenzhen, China). The shooting time was determined to be 23:00 a.m.–03:00 a.m. the next day [3], according to the observation of the pre-experiment. After that, the courtship behavior procedure was recorded through the analysis of the video data.
2.3. The SEM Sample Preparation
For SEM (Scanning Electron Microscope), the hind tibiae were collected from 1-day-old male moths, and some of the scent scales above the tibiae were carefully removed. The samples were processed following the method described by Li et al. [8]: The prepared male hind tibiae were attached to sample stubs using conductive silver paste. These samples were then coated with gold using a vacuum sputtering device and observed under a JSM-6490LV SEM (JEOL Ltd., Tokyo, Japan) at an acceleration voltage of 20 kV.
2.4. Paraffin Histological Sections
The hind tibiae of freshly emerged male moths were fixed in Bouin’s solution (LMAI Bio; Shanghai, China) for 1 day and then rinsed repeatedly with phosphate-buffered saline (PBS, pH 7.2). The samples were dehydrated through a graded series of ethanol solutions (75%, 80%, 85%, 90%, 95%, and 100%) for 1–2 h each. The samples were then cleared in a mixture of absolute ethanol and xylene (1–2 h), followed by two clearings in pure xylene (0.5 h each). The samples were embedded in paraffin, sectioned longitudinally and transversely at a thickness of approximately 6 µm, stained with hematoxylin and eosin, and observed and photographed under an optical microscope (OLYMPUS BX51; Olympus Corporation, Tokyo, Japan).
2.5. TEM Sample Preparation
For TEM (Transmission Electron Microscopy), the hind tibiae and middle tibiae of 1-day-old male moths were fixed in 2.5% glutaraldehyde (prepared in PBS) for at least 2 h. They were rinsed three times with 0.1 M PBS for 10–15 min each (fixed with 1% osmic acid for 2 h, rinsed again three times with 0.1 M PBS, and dehydrated through a graded series of acetone solutions (50%, 70%, and 90%) for 10–15 min each, followed by dehydration in pure acetone for 15–20 min each). The samples were soaked in a mixture of acetone and embedding medium (1:1) at 37 °C for 12 h and then embedded in pure embedding medium for 10–12 h. They were oven-dried at 37 °C overnight and fixed at 60 °C for 12–24 h. Ultrathin sections (50–100 nm) were cut, stained with 3% uranyl acetate and lead nitrate, and observed and photographed under the HT7700 TEM (Hitachi, Tokyo, Japan).
3. Results
3.1. The Courtship Behavior of Endoclita davidi
Behavioral observations in the laboratory: During the daytime, male and female moths climb onto the top of the net cage using their forelegs and middle legs, remaining stationary with their wings covering both sides of their bodies. The female and male E. davidi moths in cages can engage in courtship and copulation on the evening of eclosion. According to the analysis of the video taken by the infrared camera, the courtship behavior of this moth is roughly shown as follows: The male moth that is hanging moves first, the antennae are raised, the two wings change from the state of being flat on both sides of the body during the day (Figure 1A) to the state of being immobile at an angle of about 45° with the body and begin to beat the wings (Figure 1B), and even twist the abdomen. After that, the female moth also changes from a static climbing state during the day to slightly unfolding her wings and slightly swinging her abdomen. Over time, the male moth’s wings beat at a higher frequency (Figure 1C) and gradually spread the smell brush of the hind tibiae, and the end of the abdomen begins to circle or swing left and right while climbing on the top of the cage and staying near the female moth. During this process, the female moth also begins to vibrate its wings, climb, and approach the male moth. Thus, both sides were in a state of excitement, flapping their wings with high frequency, and the abdomen twists strongly to try to make the ends of the abdomen touch each other (Figure 1D). When the abdominal ends of female and male moths are stably connected, mating behavior can be carried out (Figure 1E). A detailed description of the specific process of courtship behavior in E. davidi is provided in Table 1.
In the 22 cages observed, the male moths were active in most cases. There is only one special case: the female moth flaps its wings first, the end of its abdomen sprays out a liquid, and then the male moth flaps its wings and moves closer to the female moth.
3.2. SEM of the Hind Tibiae and Hairpencils of the Male E. davidi
The hind tibiae of the male E. davidi are slightly swollen into a pear shape, with a scent brush attached to its outer side (Figure 2A). The scent brush is composed of numerous golden yellow hair pencils. Hairpencils have a smooth surface, distributed on the entire outer side of the hind tibia. The basal half of each hairpencil is broader and more oblate (Figure 2B), with the widest point reaching 120.71 µm. The apical half is finer, with the thinnest diameter ranging from approximately 14.77 to 16.42 µm, averaging 15.53 ± 0.63 µm (n = 10).
The SEM revealed that the hairpencils are attached within specialized hair follicle fossae. The hair follicles are surrounded by a raised rim resembling a swimming ring. The outer diameter of the swimming ring averages 58.80 ± 5.16 µm (n = 10), and the inner diameter averages 29.37 ± 3.43 µm (n = 10). Each hair follicle houses a single hairpencil (Figure 2B,C), and the surface of the tibia between the follicles is wrinkled (Figure 2C). The hair follicle fossae of the hairpencils are connected to glandular ducts on the tibia (Figure 2D).
The proximal end of the hairpencil is slightly intumescent, approximately 60 μm from its base, with a width of 24.61 ± 4.52 μm (n = 10) (Figure 3A). The bottom of the concave pits at the intumescent part has a distinct small hole leading into the internal glandular duct of the hairpencil (Figure 3B). The end of the hairpencil is blunt and nonporous (Figure 3C).
The surface of the hairpencil is adorned with neatly arranged longitudinal ridges (Figure 3D), with an average ridge width of 0.43 ± 0.028 μm (n = 10) and an average inter-ridge width of 1.58 ± 0.29 μm (n = 10). Between these ridges, there are uniformly distributed circular concave pits of varying sizes: the larger ones are more abundant and arranged in sequence, with diameters ranging from 1.40 to 2.05 μm and an average of 1.66 ± 0.22 μm (n = 10). Additionally, these larger pits have a secondary depression in the center, with diameters ranging from 0.48 to 0.81 μm and an average of 0.64 ± 0.12 μm (n = 10). The smaller pits have diameters ranging from 0.59 to 0.95 μm and an average of 0.68 ± 0.062 μm (n = 10), with no central depression. On the surface of the hairpencil, some spherical particles are arranged within the concave pits located between three adjacent longitudinal ridges that run the entire length (Figure 3D).
3.3. The Tissue Structure of Sex Pheromone Glands in the Hind Tibia of the Male E. davidi
Tissue section observations of the hind tibiae of the male moths reveal that the hairpencils are a hollow tubular structure (Figure 4A), and there are glandular ducts beneath the base of the hair follicles. Below the glandular duct, it connects to gland cells in the epidermal layer through epidermal gland duct-connecting cells (Figure 4B).
3.4. The Ultrastructures of Sex Pheromone Glands in the Hind Tibia of the Male E. davidi
The results of TEM for the hind tibiae and middle tibiae of a male moth are shown in Figure 5. Comparing the ultrastructures of the two, it is observed that the epidermal cells underlying the tibial segment of the hind leg are specialized gland cells (Figure 5A,B). The microvilli at the tops of the gland cells are neatly and densely arranged and penetrate the inner epidermis, while there are numerous tiny pore canals distributed in the inner epidermis above the microvilli (Figure 5A). There are distinct intercellular spaces between the gland cells, and the nuclei are large and oval (Figure 5B). In contrast, the epidermal cells underlying the tibial segment of the middle leg are less developed, with developed striated muscle bundles on the inside (Figure 5C,D).
Furthermore, the sex pheromone gland cells in the tibial segment of the male E. davidi possess many rod-shaped mitochondria. Within the cytoplasm, there is a well-developed rough endoplasmic reticulum accompanied by a large number of free ribosomes surrounding it, as well as transparent vesicles of varying sizes. These vesicles are often closely surrounded by numerous light gray round lipid droplets with relatively smooth surfaces. Additionally, there are structures such as tracheae present (Figure 6B).
4. Discussion
The courtship behavior of nocturnally active moths is strictly restricted to the scotophase. Endoclita davidi initiates courtship only in dark environments while remaining in a quiescent suspended state during the photophase (daylight or under indoor lighting). This behavior aligns with the general behavioral paradigm in moths, referred to as “photophase inhibition and scotophase activation,” thereby reinforcing the evolutionary adaptiveness of the scotophase as the optimal timing for moth courtship. Specifically, the scotophase reduces visual interference and lowers predation risk [34,35,36]. Furthermore, male E. davidi initiate wing vibration within 5 min after the start of the scotophase. This aligns with observations in other moth species: male diamondback moths (Plutella xylostella) reach the peak of courtship behavior immediately after light extinction [37], and the peak mating period of Glyphodes caesalis occurs within 0–3.5 h of the scotophase. These consistent responses reflect the high sensitivity of moths to scotophase initiation signals.
Under natural circumstances, the high-frequency wing-flapping exhibited by male moths likely involves circular flight patterns, which help in the effective dispersal of sex pheromones released by the hairpencil to attract females [38]. Notably, the wing-vibrating behaviors of male and female moths demonstrate distinct differences, consistent with the findings of Endoclita excrescens [39] and Endoclita signifier [33] within the Hepialidae family. In their natural habitat, male moths engage in group flights and initiate circling prior to female moths. Additionally, the flight behaviors of both sexes vary significantly, with the commencement of flight being closely associated with light intensity.
The surface ultrastructure of hairpencils exhibits certain conserved similarities among male moths of different species. For instance, male Mythimna separata [40] and Endoclita vietnamensis [41] possess hairpencils with hollow bristle shafts, as well as surface pores and longitudinal ridges—a structural feature consistent with the hairpencil morphology of E. davidi. SEM observations of the hind tibiae of male E. davidi revealed that the surfaces of their hind tibial hairpencils are characterized by regular longitudinal ridges and concave pits, with a species-specific distinction that some of these pits contain granular structures. These ultrastructural traits are inferred to constitute the physical basis for the golden yellow coloration of E. davidi’s hind tibial hairpencils. Notably, male Hepialus humuli [42] have been reported to utilize a combination of visual signals (from their silvery wings) and chemical signals for female attraction. Building on this finding, we hypothesize that male E. davidi may similarly employ the reflective properties associated with their golden yellow hairpencils to enhance female attraction during the scotophase. Alternatively, female E. davidi might be dually attracted during mating behavior: by olfactory cues from sex pheromones emitted by the male’s hind tibial hairpencils, and by visual stimulation derived from the golden yellow coloration of these structures.
Additionally, histological sections and SEM observations revealed that the hairpencil on the hind tibiae of male E. davidi is hollow in structure, with glandular ducts passing through the epidermal layer into the tibiae just below the base of the hair follicles. Each hairpencil is slightly swollen near its distal end, and numerous concave pits arranged regularly have small holes communicating with the lumen of the scales. Further TEM studies showed that the ultrastructural organization of the epidermal cells on the hind tibiae of male moths exhibits typical characteristics of sex pheromone-secreting gland cells. These cells are predominantly oval-shaped and clustered together, featuring numerous microvilli, large oval-shaped nuclei, and abundant organelles such as mitochondria, endoplasmic reticulum, and lipid droplets distributed within the gland cells. This structure is similar to that described by Percy and Macdonald [43]. Furthermore, Yang [44] and Hu [23] studied the ultrastructural organization of the sex pheromone glands in Holcocerus vicarius and the ultrastructural changes in sex pheromone glands at different developmental stages in Scopula subpunctaria, respectively. They found that not only the endoplasmic reticulum but also a significant number of lipid droplets are present within the gland cells. They suggested that lipid droplets serve as the raw material reservoir for sex pheromone production. Therefore, it can be inferred that the organelles within the gland cells of the hind tibiae of E. davidi are the sites for synthesizing sex pheromone precursors and storing crude products. Additionally, by comparing the structural features of the bundled striated muscles without gland cells in the mid tibiae of male E. davidi, along with the calling behavior and the unique structural characteristics of the hind tibiae of male moths, it can be speculated that during the initiation of courtship, the sex pheromone components are formed in the gland cells, pass through the glandular ducts in the epidermal layer of the tibiae, enter the intra-scale glandular ducts via the base of the hair follicles, and are ultimately released through the small holes in the concave pits between the longitudinal ridges near the distal end of the hairpencil. Furthermore, the wrinkled epidermal structure between the bases of the hairpencil of the hind tibiae in E. davidi, which resembles the layered folds and the absence of bristle coverage on the sex pheromone glands located on the intersegmental membranes between the eighth and ninth abdominal segments in Conopomorpha sinensis [22] and Algedonia coclesalis [17], is a retractable organ. During courtship in male E. davidi, the folds open, the hind tibiae expand, and the epidermis unfolds, allowing the hind tibia brushes to disperse, facilitating the full release of sex pheromones.
It is known that the majority of Lepidoptera release sex pheromones from females, such as Heliothis virescens [45], Helicoverpa armigera [16], Helicoverpa zea [14] of the Noctuidae, Lambdina fiscellaria lugubrosa [20] of the Geometridae, Holomelina lamae [46] and Estigmene acrea [47] of the Arctiidae, as well as Deqin Hepialus [24], Korscheltellus gracilis [48], and Wiseana copularis [49] of the Hepialidae. Lycorea ceres was the inaugural Lepidoptera species for which male pheromones were recorded and analyzed [50]. Subsequent studies have identified male pheromones in other Lepidoptera species, such as Plodia interpunctella [27] and Ephestia elutella [28] of the Pyralidae, Choristoneura fumiferana [51] of the Tortricidae, Lycorea halia [52] of the Nymphalidae, and Hepialus hurnuli [31] of the Hepialidae. The biological roles of volatiles released by male Lepidopteran hairpencils vary among species. For example, volatiles from the hairpencils of male Grapholita molesta exert a close-range attractant effect on females [53], while those from male Pseudaletia separata are proposed to inhibit the courtship responses of conspecific males towards females [54]. Building on the observed calling behavior of E. davidi and the principle of structural–functional adaptation [55], it can be reasonably inferred that the hind tibial hairpencils of male E. davidi function to release sex pheromones, thereby facilitating female attraction.
5. Conclusions
This study conducted a comprehensive analysis of the courtship behavior and sex pheromone gland ultrastructure of male E. davidi. Our results demonstrate that the courtship behavior of E. davidi occurs in the scotophase, during which males evert the hairpencils on their hind tibiae to lure female moths. Additionally, the hairpencil surface has regular longitudinal ridges with multiple circular pits, and small holes (formed at the pit bases) on its slightly enlarged tip areas connect to internal glandular ducts, which further link to tibia glandular canals; the hind tibia epidermal layer contains numerous glandular canals and dense microvilli, while dermal cell-specialized glandular cells harbor mitochondria, rough endoplasmic reticulum, vesicles, and lipid droplets. This work confirms that male E. davidi’s sex pheromone glands release pheromones via hind tibia hairpencils and provides a theoretical basis for further understanding lepidopteran pheromone release and communication mechanisms.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhu H.F. Wang L.Y. On the stem-borers of Chinese Heplalids (Lepidoptera:Hepialidae)Acta Entomol. Sin.1985293301+35710.16380/j.kcxb.1985.03.010 · doi ↗
- 2Wen T.C. Zhu R.C. Kang J.C. Huang M.H. Tan D.B. Ariyawansha H. Hyde K.D. Liu H. Ophiocordyceps xuefengensis sp. nov. from Larvae of Phassus nodus (Hepialidae) in Hunan Province, Southern China Phytotaxa 20131234110.11646/phytotaxa.123.1.2 · doi ↗
- 3Li X. Chen S. Zhou Q. Reproductive Behavior Rhythm of Endoclita davidi (Lepidoptera: Hepialidae), an Host Insect of Ophiocordyceps xuefengensis J. Environ. Entomol.2021431273127910.3969/j.issn.1674-0858.2021.05.23 · doi ↗
- 4Fung S.Y. Cheong P.C.H. Tan N.H. Ng S.T. Tan C.S. Nutrient and Chemical Analysis of Fruiting Bodies of a Cultivar of the Chinese Caterpillar Mushroom, Ophiocordyceps sinensis (Ascomycetes)Int. J. Med. Mushrooms 20182045946910.1615/Int J Med Mushrooms.201802625229953361 · doi ↗ · pubmed ↗
- 5Lu Z.H. Chen S.J. Review on the practice and perspective of the industrial production of Ophiocordyceps sinensis J. Environ. Entomol.201638243010.3969/j.issn.1674-0858.2016.01.2 · doi ↗
- 6Wang Y. Zhang X.H. Zhang S.B. Wang X.L. Preliminary Study on Production and Antimicrobial Activity of Triterpenoidsfrom Xuefeng Congcao(Ophiocordyceps xuefengensis) by Liquid Cell Culture China TCM Sci. Technol.2020273437
- 7Chen S. Zhou Q. Li G. DNA barcoding of various developmental stages of Endoclita davidi (Lepidoptera: Hepialidae) based on mt DNA COI gene sequence Acta Entomol. Sin.20176068169010.16380/j.kcxb.2017.06.008 · doi ↗
- 8Li G. Zhou Q. Chen S. SEM observation of antennal sensilla of adult Endoclita davidi, one host of Ophiocordyceps xuefengensis J. Chin. Electron Microsc. Soc.201736637010.3969/j.issn.1000-6281.2017.01.011 · doi ↗