Mapping Antennal Sensilla of Boxelder Bugs (Boisea trivittata) as the First Step in Understanding Overwintering Aggregation Behavior
Anika Sharma, Stephen A. Kells

TL;DR
This study maps the sensory structures on boxelder bug antennae to better understand their overwintering behavior and improve pest control.
Contribution
The first detailed mapping of antennal sensilla in boxelder bugs to study their aggregation behavior.
Findings
Boxelder bugs have 20 subtypes of antennal sensilla across seven categories, with distribution varying by segment.
Chemoreceptors for olfaction are concentrated on the distiflagellomere, while mechanoreceptors dominate closer to the head.
Two patches of sensilla coeloconica were found on the outer lateral pedicel and basiflagellomere.
Abstract
Insects use sensory structures called sensilla to sense the world around them, for example, to smell, detect the temperature and humidity, or position their limbs. Understanding the placement and distribution of these sensilla will enable further studies to advance prevention and control measures against insect pests. Boxelder bugs are free-living herbivores in the landscape; however, during the fall season, they exhibit behaviors to avoid adverse conditions, resulting in these insects entering buildings, such as residences, industrial structures, and commercial facilities. The majority of scent receptors on boxelder bugs are localized on their last antennal segment, with a few potential patches on the second and third segments. The majority of the receptors on segments closer to the head detect physical and mechanical changes. Knowledge of the locations of these sensory structures will…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
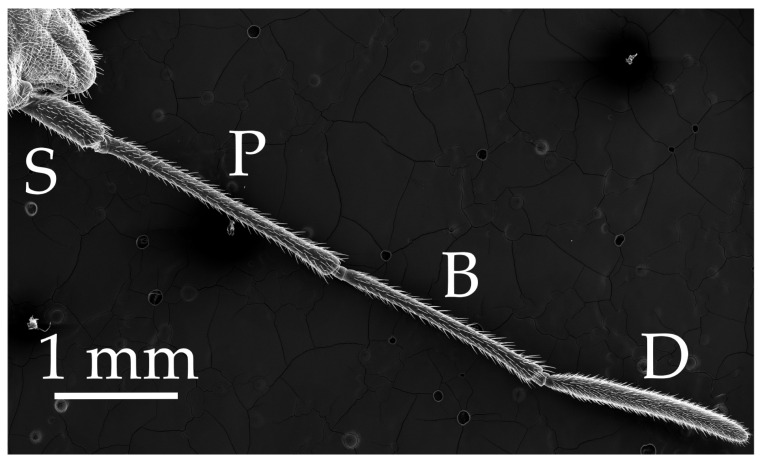 Figure 1
Figure 1 Figure 2
Figure 2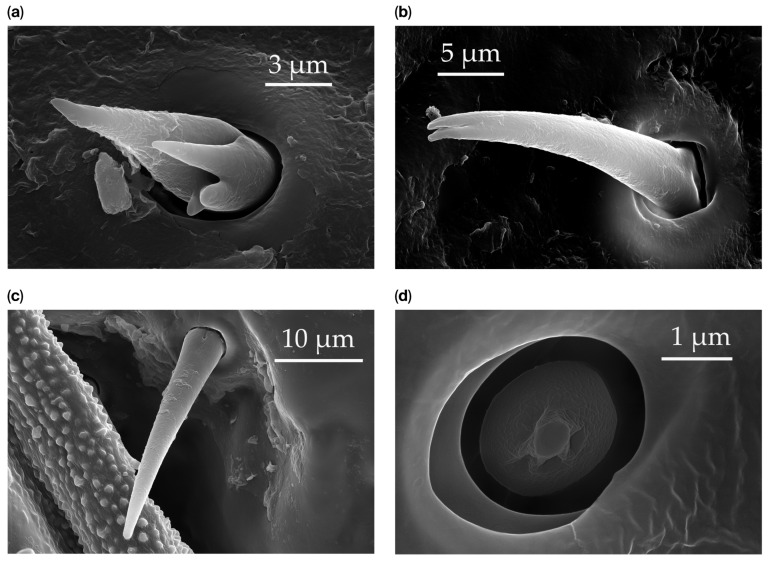 Figure 3
Figure 3 Figure 4
Figure 4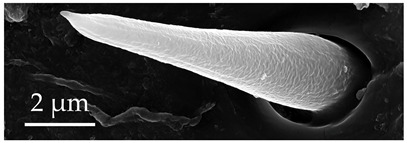 Figure 5
Figure 5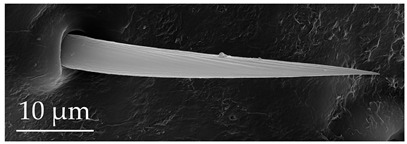 Figure 6
Figure 6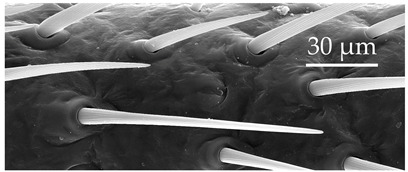 Figure 7
Figure 7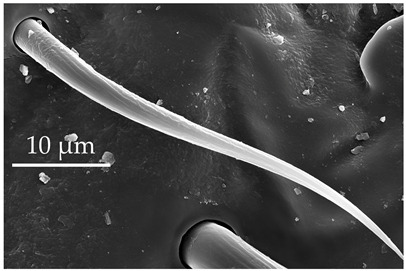 Figure 8
Figure 8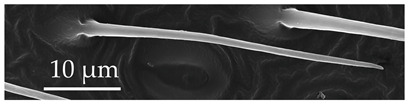 Figure 9
Figure 9 Figure 10
Figure 10 Figure 11
Figure 11 Figure 12
Figure 12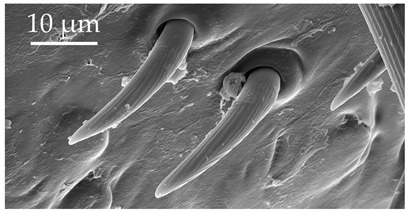 Figure 13
Figure 13 Figure 14
Figure 14 Figure 15
Figure 15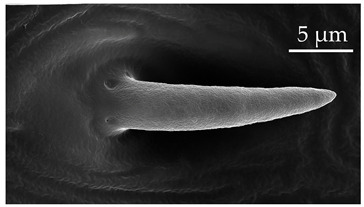 Figure 16
Figure 16 Figure 17
Figure 17 Figure 18
Figure 18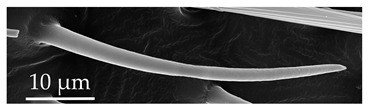 Figure 19
Figure 19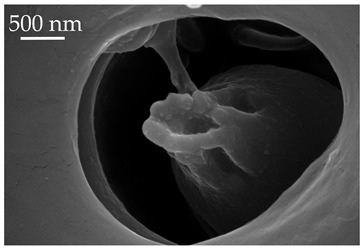 Figure 20
Figure 20 Figure 21
Figure 21 Figure 22
Figure 22 Figure 23
Figure 23 Figure 24
Figure 24- —U.S. Department of Agriculture’s National Institute of Food and Agriculture
- —Minnesota Pest Management Association
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurobiology and Insect Physiology Research · Hemiptera Insect Studies · Insect-Plant Interactions and Control
1. Introduction
The eastern boxelder bug, Boisea trivittata (Say) (Heteroptera: Rhopalidae), is native to North America and a specialist herbivore of the boxelder tree, Acer negundo L. (Aceraceae) [1], and other plants [2]. With increasing temperatures during springtime, adults emerge from overwintering harborages to initiate feeding and begin their reproductive season, which lasts through the summer [3]. In late summer or early fall, both nymphs and adults cease feeding and depart from host trees [4]. In late fall and in preparation for winter survival, adult B. trivittata aggregate in the natural cracks and crevices of rock piles, tree trunks, and similar sites with strong sun exposure. In anthropogenic structures, boxelder bugs mainly aggregate on the outside of south- or west-facing surfaces and move into buildings, including homes, commercial and industrial buildings, and other structures.
Boxelder bugs are generally not considered pests while in the landscape. However, their status as a pest changes when they start to aggregate in buildings, clustering under the exterior structural layers, such as exterior vinyl or wood siding, metal cladding, or brick veneer. Depending on the gaps provided in the exterior infrastructure, with repeated diurnal heating and cooling of building surfaces, boxelder bugs may subsequently move deeper into building structures. This subsequent movement results in boxelder bugs entering living spaces, mainly becoming an aesthetic pest for people adverse to insect activity in their homes.
When boxelder bugs enter operational areas of commercial or industrial sites, they can create serious economic damage. As economic pests, their aggregation behavior and subsequent movement may contaminate food or pharmaceutical products, or damage electrical and other equipment. Such damage from boxelder bugs is seldom reported publicly, but citations of other species with similar behaviors demonstrate the potential extent of damage. For example, Halyomorpha halys adults also aggregate inside anthropogenic structures to overwinter [5,6], and several car-carrier vessels from South Korea were temporarily refused entry into Australia because of the detection of overwintering H. halys in the vessels and cars [7]. This caused significant economic losses, as stink bugs delayed 10,000 vehicles following increased biosecurity checks. Understanding this aggregation behavior is the first step in predicting, preventing, or reducing the use of structures as overwintering sites and avoiding economic losses.
The aggregation of adults to the host tree is mediated by semiochemicals released by the boxelder tree [1]. Similarly, the aggregation of adults away from the host tree is believed to be mediated by pheromones [8]. Boxelder bug antennae consist of four segments: the scape, the pedicel, and two flagellomeres. The different sensilla present on all these segments play a significant role in habitat selection, aggregation behavior, host finding, courtship, and mating behaviors [9]. With their potential to cause damage from overwintering behaviors, it is necessary to investigate the antennae of these insects to understand possible locations for further electrophysiology recording used to support behavioral studies. Previous studies have demonstrated that insects detect semiochemicals through olfactory receptor neurons (ORNs) enclosed in olfactory organs [10,11]. For example, Cimex lectularius L. detects semiochemicals through ORNs in the olfactory sensilla on their antennae [12,13]. A study conducted by Olson et al. demonstrated that olfaction sensilla related to a patch on the distal pedicel mediated off-host aggregation behavior [14]. Such a specialty patch was not part of the regular areas of olfaction on the flagellomeres of this insect, and traditional electroantennography from whole antennae was very difficult with bed bugs.
To understand the placement and number of sensilla in preparation for further pheromone work, we performed a morphological study of boxelder bug antennae via scanning electron microscopy. The aim of the present study was to understand the morphology, location, and distribution of the antennal sensilla. This work is a prerequisite for future electrophysiological and behavioral studies of the antennal sensory system involved in intraspecific chemical communication, with the aim of eventually determining an effective way to control or reduce overwintering aggregations of boxelder bugs in human-made structures.
2. Materials and Methods
2.1. Insect Collection and Sample Preparation
Aggregating adult boxelder bugs were mass collected from boxelder trees (Acer negundo L.) and nearby buildings around the St. Paul Campus, University of Minnesota (Falcon Heights, MN, USA), and from a residence in Lino Lakes, MN from September to November over three years (2022–2024). The collected specimens were preserved in 70% ethanol (Decon Labs, Inc., King of Prussia, PA, USA) until required for morphological examination. Whole heads with intact antennae were excised from the preserved samples for scanning electron microscopy.
The excised heads of the adult boxelder bugs were transferred to vials containing 70% ethanol and sealed afterwards, with one sample/vial. The samples were sonicated for 2 min to remove foreign particulates, and then they were dehydrated over four days to stabilize soft tissues in a graded alcohol series of 70%, 80%, and 100% ethanol. An initial survey of the SEM micrograph images of the collected samples preliminarily confirmed specimen handling techniques, and the best images with a lack of damage were selected.
2.2. Scanning Electron Microscopy (SEM)
The excised heads of the boxelder bugs (n = 12), both males and females, were individually mounted either dorsally or ventrally on aluminum stubs (AMRAY 1000/1200, aluminum, 15 mm pin height; Electron Microscopy Sciences, Hatfield, PA, USA) with double-sided carbon conductive tape (PELCO Tabs™, Carbon Conductive Tabs, 25 mm dia) and sputter-coated with 60/40 gold–palladium with a 2–3 nanometer thickness (Cressington 108 Auto; Ted Pella, Inc., Redding, CA, USA). The initial SEM micrographs were obtained using a Hitachi SEM model S3500N (Hitachi High Technologies Corp., Tokyo, Japan), and the final high-resolution images were obtained with an accelerating voltage of 10 kV to 20 kV via a Thermo Fisher Apreo 2S SEM (Thermo Fisher, Waltham, MA, USA).
2.3. Sensilla Mapping and Enumeration
The identification of sensilla and examination of their features were based on Nowinska and Brozek [15,16], as well as a morphological comparison of the antennal sensilla of different species of Heteroptera characterized in studies conducted by Silva et al. [17], Gonzaga-Segura et al. [18], and Taszakowski et al. [19]. Micrographs of each antennal segment, including the scape, pedicel, basiflagellomere, and distiflagellomere, were taken to enumerate the different types of sensilla. This enumeration was carried out by counting the sensilla in 100 μm^2^ sections from the dorsal and ventral surfaces of each segment, which enabled the inclusion of sensilla on the lateral surfaces in these estimates. The type of sensilla, their placement, and their density were tabulated with a representative image.
3. Results
3.1. Gross Morphology of Antennae
Boxelder bug antennae consist of four antennomeres, including the scape (883.6 µm), pedicel (2.2 mm), basiflagellomere (1.8 mm), and distiflagellomere (1.7 mm; Figure 1). The scape is a short and thick segment. The pedicel is the longest segment, whereas the basiflagellomere and distiflagellomere are subequal in size. The density of sensilla increased from the first to the last segment, with the distiflagellomeres (fourth segment) having the greatest density of sensilla. Sexual dimorphism was not apparent in either antenna morphological or sensilla characteristics.
3.2. Sensilla Types and Arrangement
The principal system for describing the type of sensilla in B. trivittata is based on determining whether they have a porous or nonporous surface and the type of connection with the cuticle (i.e., a flexible or inflexible socket) (Figure 2). Then, the sensilla can be differentiated on the basis of size, shape, the presence or absence of pores, and the presence of a grooved or smooth surface. Seven types of sensilla, subdivided morphologically, were identified in the boxelder bug specimens: sensilla trichoidea (ST), sensilla chaetica (SCh), sensilla basiconica (SB), sensilla coeloconica (SCo), sensilla campaniformia (SCa), sensilla ampullacea (SA), and sensilla bell-mouthed (SBm).
Generally, flexible sensilla emerge from cuticular sockets, whereas inflexible sensilla appear contiguous with the cuticle (Figure 2). Aporous sensilla can have either flexible or inflexible sockets, whereas porous sensilla always arise from inflexible sockets. Sensilla trichoidea can be distinguished as flexible hair-like structures (Table 1). Sensilla chaetica are long thick hairs (Table 2). Sensilla basiconica are cone-shaped with a porous or aporous surface (Table 3). Sensilla coeloconica are short peg-like structures embedded in a pit (Table 4). Sensilla campaniformia are flat, oval structures on the surface of the cuticle (Table 5). Sensilla ampullacea consist of open pores on the cuticle surface, while sensilla bell-mouthed have a characteristic pocket on the surface (Table 5). Some modifications of sensilla occur, though the modified sensilla are not differentiated enough to be considered to belong to additional subclasses (Figure 3).
4. Discussion
Boxelder bugs have a distribution of sensilla consistent with biological function, and understanding this distribution will enable the identification of regions of focus for further electrophysiological studies. In C. lectularius, electroantennography related to the aggregation pheromone was best optimized when a patch was located on the outer lateral distal pedicel [14]. This patch on bed bugs was located after considerable work involving progressively removing structures and ultimately stopping the aggregation pheromone response. Further, the work indicated an arrestant rather than an attractant behavior in response to the aggregation pheromone [23]. This present survey of the sensilla on boxelder bugs provides the placement of key receptors along all four segments without having to assume that chemoreception is solely isolated to the distal two flagellomeres. Further, interferences in signals from unrelated receptors can be minimized during electroantennography through the placement of different electrodes closer to the sensilla of interest. Given the short period during which boxelder bugs exhibit seasonal aggregation and the potential for additional procedures to determine behavioral response, such knowledge is important for deciding the next steps in evaluating the aggregation pheromones used by boxelder bugs.
The general function of sensilla on boxelder bug antennae can be surmised based on their morphology and distribution. Porous sensilla are related to olfaction, whereas mechano-, thermo-, and hygro-receptors are often associated with aporous sensilla [19]. The type of cuticular attachment—a flexible or inflexible socket—serves as an important morphological indicator of sensillum function. Sensilla with inflexible sockets are generally associated with chemosensory roles, as neuro-dendrites are inserted inside the sensillar shaft. Mechanoreceptors generally have a flexible attachment with the cuticle, possessing a membrane that enables movement at the base. Visible porosity, or a lack thereof, provides indications of chemo- versus mechano-reception. Chemoreceptors can be multiporous olfactory sensilla or uniporous gustatory sensilla, whereas mechanoreceptors and thermo-hygroreceptors are attributed to aporous sensilla [24,25,26,27,28]. The most common mechanoreceptors are sensilla trichodea, sensilla chaetica, and sensilla campaniformia [24,28,29]. Chemoreceptors consist of one or more sensory neurons responsible for gustation and olfaction. Common chemoreceptors are sensilla basiconica, sensilla coeloconica, and sensilla trichodea [28,30]. In boxelder bugs, there are several exceptions to these general characteristics; specifically, some sensilla have flexible sockets and at least one pore (e.g., ST1 and SB1), and some have no pores but appear to be inflexible (ST5) (Table 1 and Table 3). Such exceptions are noted for future work. Sensilla are distributed across the antenna and can be summarized by antennal segment.
4.1. Scape
The scapes of boxelder bugs have numerous sensilla types, though at a lower density than other segments. The scape has sensilla trichoidea (ST1, ST2, ST3, and ST4), sensilla chaetica (SCh1, SCh2, SCh3, and SCh4), sensilla basiconica (SB1), sensilla ampulacea (SA), and sensilla campaniformia (SCa), which are aporous sensilla embedded in a flexible socket. In boxelder bugs, based on the morphological study, it can be stated that sensilla trichoidea, sensilla chaetica, sensilla campaniformia (SCa), and sensilla basiconica (SB1) are responsible for mechanoreception (Table 1, Table 2, Table 3 and Table 5). Sensilla trichoidea and chaetica are the dominant mechanoreceptors in B. trivittata and numerous other insect groups [14,15,18,31]. Sensilla trichoidea are considered the most common mechanoreceptors, and, depending on their location on the antennomere, they act as either exteroceptors or proprioceptors [32,33].
In B. trivittata, the proprioceptive sensilla (SB1), located within the joints at the proximal end of the scape and at the articulation between the scape and pedicel, function to detect the positional orientation of the antenna [18]. These sensilla are often recognized as short trichoidea, chaetica, or basiconica. An investigation of various heteropteran species, including those in the infra-orders Cimicomorpha, Gerromorpha, Nepomorpha, and Pentatomomorpha, characterized these sensilla as mechanoreceptive sensilla basiconica [15,34,35,36]. In Leptoglossus zonatus, small sensilla present between the scape and pedicel were identified as small, smooth sensilla trichoidea [18], which is in contrast to our finding of SB1 at these sites.
Sensilla campaniformia (SCa, Table 5) are strain sensors/detectors that monitor mechanical deformations of the cuticle and provide mechanical stimulation. These sensilla are predominantly localized near articulations [37]. In B. trivittata, a cluster of SCa was identified, exhibiting morphological features consistent with campaniform sensilla in Leptoglossus [19]. These sensilla are located proximally on the dorsal side of the scape and are also distributed sparsely across other antennomeres. Similarly, in Leptoglossus, a group of dome-shaped, aporous sensilla, named the A4 type, were documented on the ventral side of the scape by Taszakowski et al. [19], and these are morphologically analogous to mechanosensilla SCa. In addition to their presence on the antennae, SCa are extensively distributed on legs, including the trochanter, femur, tibia, and tarsal segments [38,39,40,41]. These sensilla occur as both isolated individual units and organized clusters, often positioned in regions subject to significant mechanical stress, such as near articulations or on cuticular surfaces that undergo frequent deformation during locomotion.
4.2. Pedicel
The pedicel of boxelder bugs has sensilla trichoidea (ST1, ST2, and ST3), sensilla chaetica (SCh1 and SCh2), sensilla basiconica (SB1), sensilla coeloconica (SCo2), sensilla ampulacea (SA), and sensilla campaniformia (SCa) (Table 1, Table 2, Table 3, Table 4 and Table 5). Sensilla trichoidea are the most numerous, with the other sensilla types scattered and interspersed along the segment. Sensilla coeloconica (SCo2) occur in a patch at the distal end of the outer lateral pedicel and basiflagellomere (Figure 4). Both sensilla coeloconica (SCo2) and sensilla ampulacea (SA) are assumed to be thermo-hygroreceptors. Their role is thought to support behaviors that prevent water loss and provide protection against temperature fluctuation. Kleineidam and Tautz [42] suggested two possibilities regarding the morphological structure of SCo and SA: both are embedded below the cuticle surface to either save space on the surface of the antennae or protect the sensory pegs against environmental extremes. However, with the placement of SCo2 in a latero-distal patch, there may be additional functions, given the placement of a similar patch on the pedicel of C. lectularius [14]. This patch is noted for further behavioral studies. The numbers of SCa gradually decrease in the distal segments.
4.3. Flagellomeres
Along the main segment, the basiflagellomere consists mainly of sensilla chaetica (SCh1) mechanoreceptors and sparsely interspersed sensilla campaniformia. Similar to the pedicel, there is a cluster of SCo2 on the outer lateral side, towards the terminus of this segment. The presence of these SCo2 should be investigated further to determine whether there are any differences between the patches on these two segments. The presence of numerous mechanoreceptors, and the incidental stimulation thereof, presents an additional risk of extra signal noise between the sites of olfaction and the placement of different and indifferent probes. Future studies will have to consider or accommodate these degraded signal-to-noise ratios.
The distiflagellomere of the boxelder bug shows specialization towards olfaction, as this terminal segment has a distinct diversity of sensilla with one or more pores, including SB2, SB3, SB3a, SB4, SB5, and SCo1 (Table 3 and Table 4). Chapman [20] stated that greater numbers of chemoreceptors increase the possibility of perception of the chemical environment. Still, there are mechanoreceptors on this last antennal segment likewise showing diversity in length and conformation, such as ST3, ST4, ST5, SCh1, SCh4, and SCa (Table 1, Table 2 and Table 5).
In this study, we classified porous sensilla into several forms: SB2, SB3, SB3a, SB4, and SB5. However, based on Steinbrecht [43] and Altner and Prillinger [44], the porous system and other morphological features differ in two (SB2-SB3a and SB4-SB5) distinguished groups. The first group consists of SB2, SB3, and SB3a, and they possess a non-grooved wall densely covered with clearly visible multiple pores along the surface. The other sensilla group (SB4 and SB5) has a porous wall that is grooved. The SB4 group is represented by deep longitudinal grooves and a single terminal pore. In B. trivittata, SB5 have shallow grooves consisting of linearly arranged pores. Sensilla basiconica in our study belong to both “single-walled wall pore” (SB2, SB3, SB3a, and SB5) and “double-walled wall pore” classes (SB4) [41,44].
Morphologically, chemosensitive sensilla basiconica are well represented in B. trivittata. Sensilla with olfactory functions are confined to the distiflagellum, suggesting that the last segment is a primary “olfactory segment.” Evidence of the terminal segment as a principal site for olfaction has also been reported in other heteropteran species. A study conducted by Andersen and Ball [45] reported more nervous tissue in the fourth segment of Oncopeltus fasciatus than in the remaining three segments. Oxycarenus laetus Kirby (Hemiptera) also reported a high sensilla density on the last segment compared to the other segments [46].
5. Conclusions
This survey was undertaken to map the various sensilla of boxelder bugs, and the identification of the distiflagellomere as the principal site of olfaction helps to focus our electrophysiology work on this site. The presence of two SCo2 patches on the latero-distal pedicel and basiflagellomere suggests that extra work should be undertaken to determine whether seasonal aggregation behavior would still occur after removing the distiflagellomere. The presence of many mechanical sensilla and the distance from the site of neuro-activity suggest that signal interference may occur, and focusing specifically on sites will reduce unnecessary noise. This work greatly enhances our ability to conduct further studies within the short period during which boxelder bugs display their seasonal aggregation behavior. By understanding the seasonal aggregation behavior of this insect, we can examine ways of preventing their occurrence in human-made structures when they go from being a free-living insect to a structural pest in residences, as well as at commercial and industrial sites.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Schwarz J. Gries R. Hillier K. Vickers N. Gries G. Phenology of semiochemical-mediated host foraging by the western boxelder bug, Boisea rubrolineata, an aposematic seed predator J. Chem. Ecol.200935587010.1007/s 10886-008-9575-319123035 · doi ↗ · pubmed ↗
- 2Aragon-Traverso J.H. Sanabria E.A. New record of the boxelder bug Boisea trivittata in Argentina suggests a rapid spread Rev. Peru. Biol.202532 e 2898310.15381/rpb.v 32i 1.28983 · doi ↗
- 3Smith R.C. Shepherd L. The life history and control of the boxelder bug in Kansas Trans. Kans. Acad. Sci.19374014315910.2307/3625403 · doi ↗
- 4Tinker M.E. The seasonal behavior and ecology of the boxelder bug (Leptocoris trivitatus) in Minnesota Ecology 19523340741410.2307/1932836 · doi ↗
- 5Hancock T.J. Lee D.H. Bergh J.C. Morrison W.R.III Leskey T.C. Presence of the invasive brown marmorated stink bug Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) on home exteriors during the autumn dispersal period: Results generated by citizen scientists Agric. Forest Entomol.2019219910810.1111/afe.12312 · doi ↗
- 6Inkley D.B. Characteristics of home invasion by the brown marmorated stink bug (Hemiptera: Pentatomidae)J. Entomol. Sci.20124712513010.18474/0749-8004-47.2.125 · doi ↗
- 7Dowling J. Stink bugs delay 10,000 Hyundai and Kia cars following increased biosecurity checks Drive 15December 2019 Available online: https://www.drive.com.au/news/stink-bugs-delay-10-000-hyundai-and-kia-cars-following-increased-biosecurity-checks/(accessed on 9 September 2025)
- 8Schowalter T.D. Overwintering aggregation of Boisea rubrolineatus (Heteroptera: Rhopalidae) in western Oregon Environ. Entomol.1986151055105610.1093/ee/15.5.1055 · doi ↗