Parasitic Recognition Behavior of Telenomus remus Nixon, an Important Egg Parasitoid of Spodoptera frugiperda (J. E. Smith)
Xiaolong Ma, Yujie Luo, Qiufen Zhao, Ruohan Zhang, Haiyan Lin, Jian Huang, Zhuhong Wang

TL;DR
This paper studies how Telenomus remus, a parasitic wasp, identifies and avoids re-parasitizing eggs of the fall armyworm, offering insights for biological control strategies.
Contribution
The study reveals how T. remus uses experience and environmental cues to avoid superparasitism, enhancing biological control applications.
Findings
T. remus performs a distinctive '8'-shaped marking to deter conspecifics from parasitizing the same host.
Experienced females can recognize marked eggs across a wide temperature range and time intervals.
Parasitism rates increase with host density, and searching time decreases.
Abstract
The fall armyworm, Spodoptera frugiperda, is a major pest of maize. Since its invasion of China in 2019, it has rapidly established itself and spread to most regions of the country. Its polyphagous feeding habits, high feeding capacity, rapid development cycle, robust reproductive potential, resistance to pesticides, and migratory tendencies pose significant challenges for its control. Utilizing natural enemies for biological control is therefore an essential strategy. The parasitoid, Telenomus remus, serves as an important egg parasitoid targeting various lepidopteran pests, and has demonstrated substantial efficacy against S. frugiperda. This study examined the host discrimination and marking behavior of T. remus, specifically assessing its ability to identify host eggs marked by itself or conspecific individuals under different temperature conditions and oviposition intervals. The…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1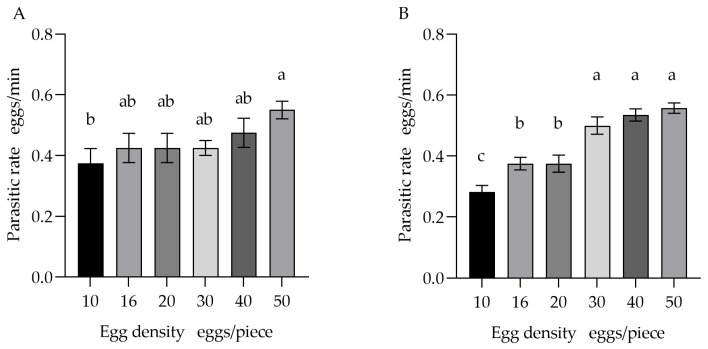 Figure 2
Figure 2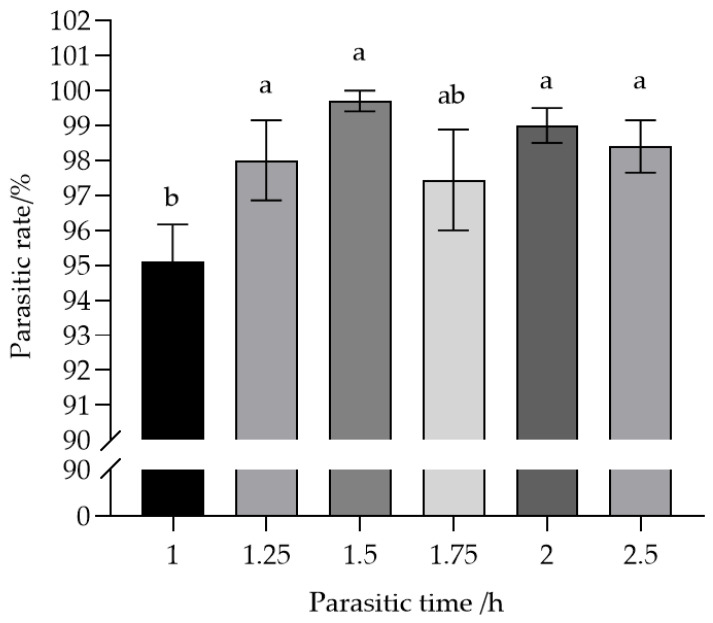 Figure 3
Figure 3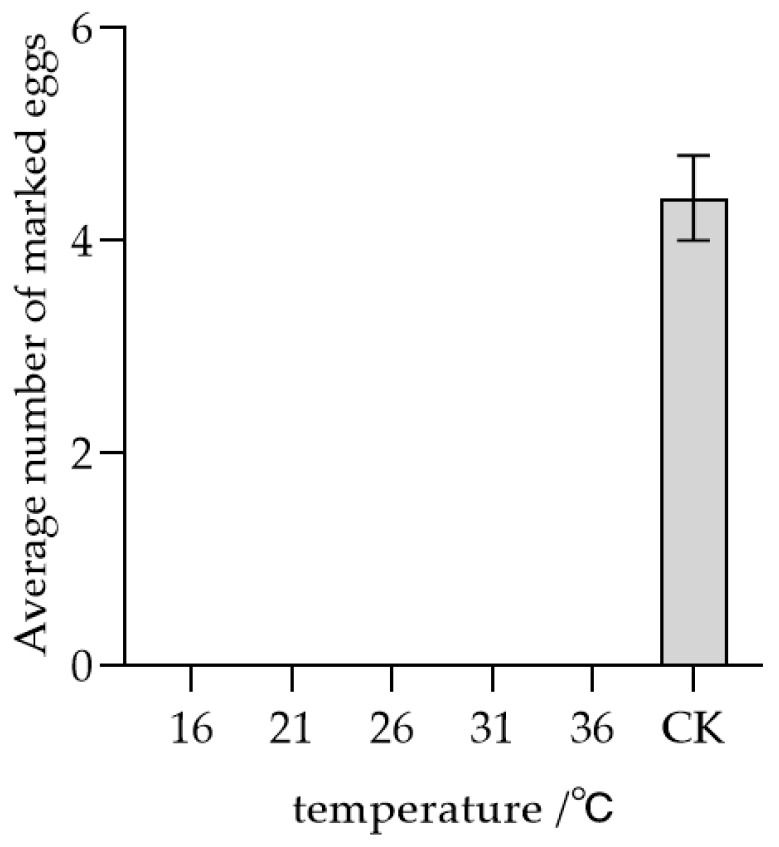 Figure 4
Figure 4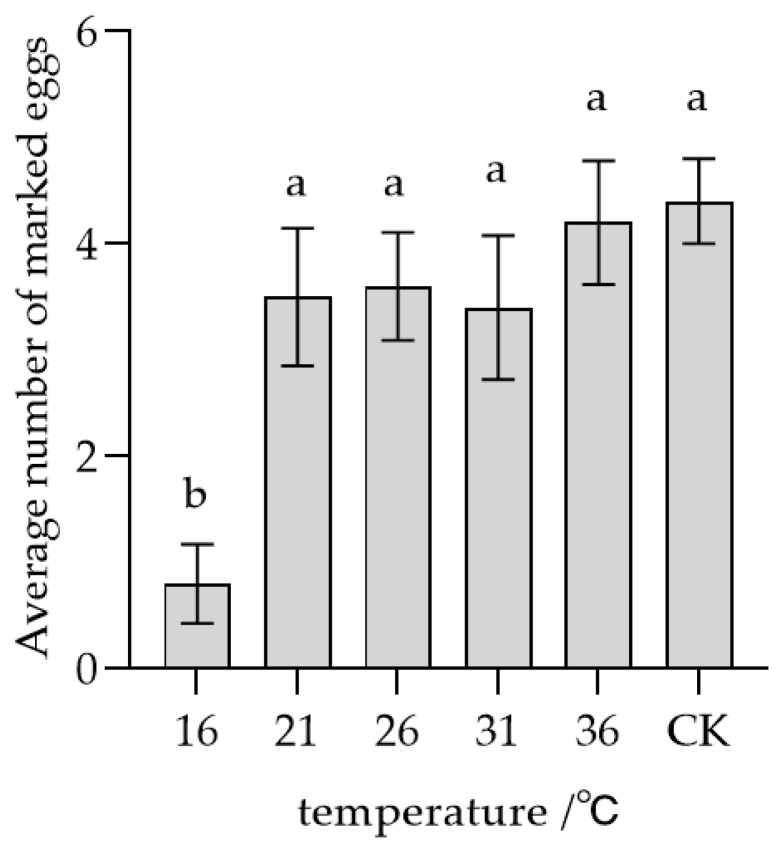 Figure 5
Figure 5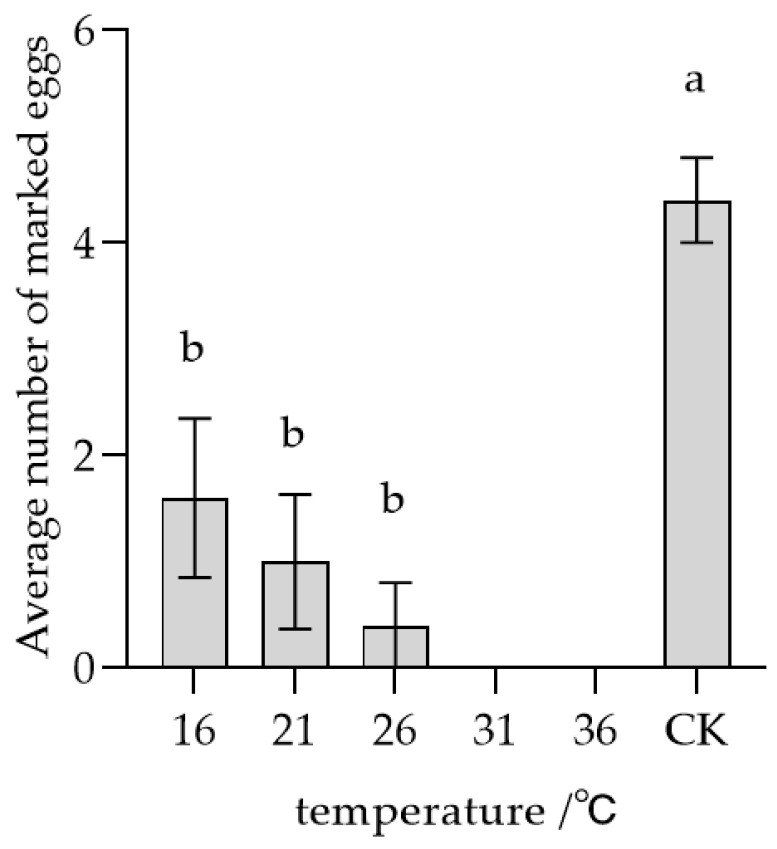 Figure 6
Figure 6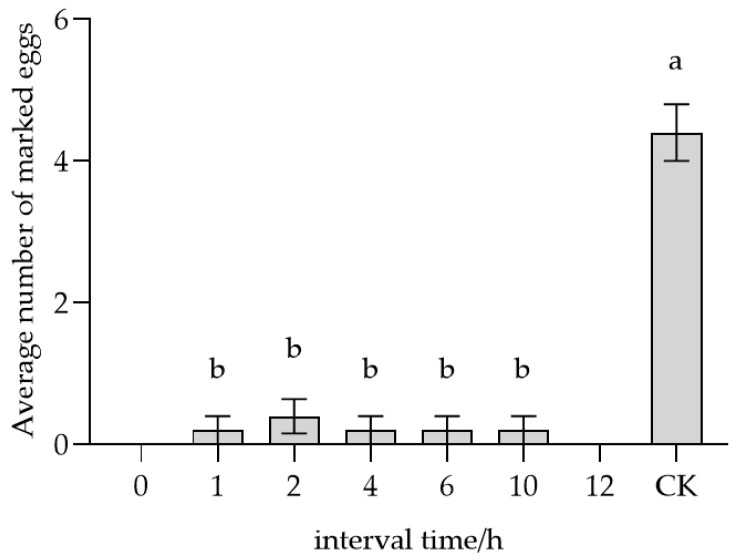 Figure 7
Figure 7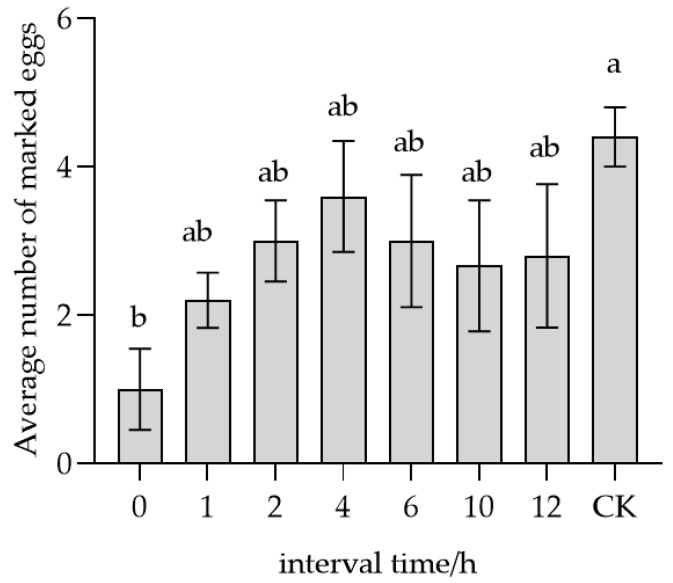 Figure 8
Figure 8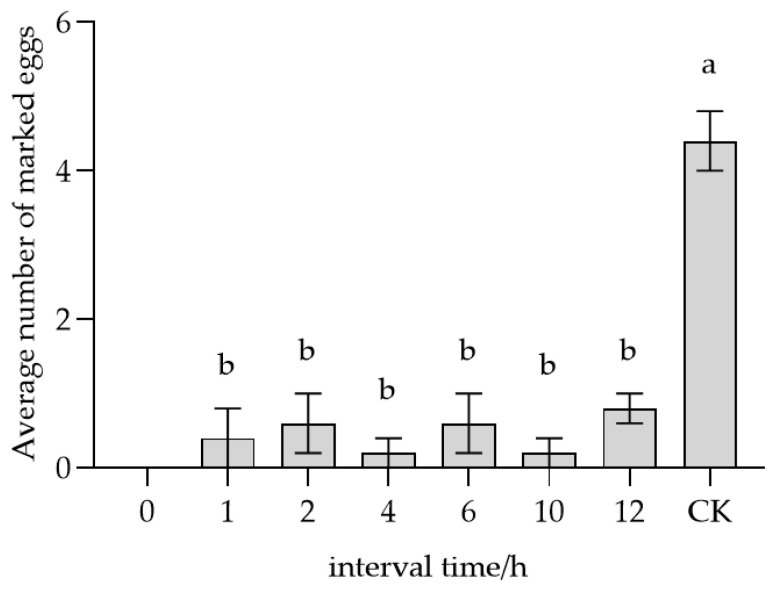 Figure 9
Figure 9- —National Key R&D Program of China
- —National Natural Science Foundation of China
- —Natural Science Foundation of Fujian Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect-Plant Interactions and Control · Insect Resistance and Genetics · Insect behavior and control techniques
1. Introduction
To ensure sufficient nutrition for their progeny, female parasitoids generally avoid ovipositing on hosts that have already been parasitized, thereby reducing competition among offspring for host resources [1]. This ability is known as host discrimination [2]. Host discrimination can be further delineated into intraspecific and interspecific recognition, with the former encompassing self-discrimination and conspecific discrimination [3]. Self-discrimination pertains to the female parasitoid’s ability to identify hosts they have previously parasitized, whereas conspecific discrimination involves distinguishing hosts parasitized by other females of the same species [3,4]. Parasitoids predominantly utilize sensory organs such as antennae and ovipositors to facilitate host discrimination. The sensilla located on the antennae are chiefly responsible for detecting external cues on the host surface, while those on the ovipositor assess the internal condition of the host [5].
A key mechanism underlying host discrimination is the use of marking pheromones. During oviposition, parasitoids frequently deposit marking substances on the host surface, which can be detected by subsequent females to avoid superparasitism [6,7,8]. More than 200 species of female wasps from Ichneumonoidea, Chalcidoidea, Proctotrupoidea, Bethyloidea, and Cynipoidea have been reported to use host markers to identify already parasitized hosts [9]. For example, females of Aprostocetus prolixus LaSalle & Huang apply a marking secretion to the oviposition slit after egg-laying, which deters subsequent parasitoids from parasitizing the same host [10]. These host-marking pheromones consist mainly of saturated and unsaturated hydrocarbons [11], and most parasitoids depend on them for recognizing parasitized hosts [11,12].
The efficiency of host recognition and superparasitism avoidance is influenced by multiple factors. These include the time elapsed since the host was last parasitized, the developmental stage of parasitoid eggs within the host, the quantity of eggs previously deposited, host density, the duration of activity of host-marking compounds, and the interval between oviposition events due to a shortage of suitable hosts [3,13,14,15,16]. Typically, upon detecting a marked or parasitized host, a parasitoid will abandon it and search for unparasitized ones, thereby enhancing biological control efficacy. However, the ability to recognize these marks declines over time [17,18,19]. There are two main mechanisms proposed to explain this phenomenon:
The first involves the chemical properties of the pheromone itself. One hypothesis suggests that the marking pheromone comprises two functional components: a short-lived, highly active component that primarily inhibits the marking female from re-ovipositing in the same host, and a longer-lasting, less active component that mainly deters conspecifics. Over time, the short-lived component degrades, reducing its self-recognition effect, while the more stable, long-lasting component continues to provide a degree of within-species recognition [20]. Environmental factors such as temperature can accelerate the volatilization and degradation of these pheromone molecules, shortening the effective recognition window [21]. For instance, the host-marking pheromone deposited by T. podisi Ashmead and T. euschisti Ashmead on stink bug eggs remains effective for recognition only within one hour after secretion [22]. Thus, factors such as temperature and time intervals may interfere with the parasitoid’s ability to distinguish between its own marks and those of others.
The second involves the cognitive processes of parasitoids—learning and memory. The parasitoids retain a transient memory of having parasitized a host, but this memory weakens and ultimately disappears over time [23]. In fact, the learning and memory behavior also plays a crucial regulatory role in host recognition. For example, Trichogramma ostriniae Pang et Chen exhibit both learning and memory capabilities and can acquire host-marking cues to enhance host-searching efficiency and modify its oviposition strategy [24]; Anastatus japonicus Ashmead shows significantly stronger attraction to volatiles emitted by litchi stink bugs after training with β-caryophyllene [25]; Asobara japonica Belokobylskij improves its recognition ability through repeated parasitism experiences [26]; Sclerodermus pupariae Yang et Yao demonstrates increased search efficiency following initial host contact, indicating that learning reinforces its host preference [27]. These species, T. ostriniae, A. japonicus, A. japonica, and S. pupariae are all advantageous for large-scale biological control. Through host recognition, continuous learning allows parasitoids to accumulate discriminatory experience, optimize foraging behavior, and improve host selection efficiency, thereby minimizing time wasted on already parasitized hosts.
Many parasitoids in the genera Telenomus and Trissolcus possess host recognition abilities. However, while most Trissolcus species employ a uniform marking method, Telenomus species exhibit diverse recognition-marking strategies (Table 1). In China, T. remus Nixon serves as a key parasitoid of Spodoptera frugiperda (J. E. Smith) and plays a significant role in its biological control. Nonetheless, as a monophagous parasitoid, superparasitism can still occur under mass-rearing conditions [3]. Comprehensive studies on the host discrimination behavior of T. remus remain scarce. Understanding this behavior is essential for improving its field deployment and enhancing its host selection efficiency under practical conditions.
Therefore, this study takes T. remus as the object, through observing its marking behavior on the surface of the host, dissecting the host to confirm the internal parasitic state, and comparing experiments under different temperature conditions and time intervals, in order to explore the following: (1) whether and how the marking of the host egg affects the recognition behavior of the parasitoid by volatilization or degradation; (2) a systematic evaluation of the self- and conspecific recognition behavior of T. remus toward marked host eggs. The findings are expected to provide scientific guidance for optimizing the field application strategy of T. remus.
2. Materials and Methods
2.1. Insect Sources
S. frugiperda larvae were initially collected from a maize field located in Shoushan, Fuzhou City, Fujian Province. A laboratory colony has since been established and maintained for over 10 generations under controlled conditions of 26 ± 1 °C, relative humidity of 70 ± 5%, a photoperiod of 16L:8D, and a light intensity of 1000 lux.
T. remus were also collected from the same site and have been continuously reared in the laboratory using S. frugiperda eggs as hosts. The rearing conditions were maintained at 26 ± 1 °C, relative humidity of 70 ± 10%, a photoperiod of 14L:10D and a light intensity of 1000 lux. Insects from both colonies were reared for multiple generations under these controlled conditions, and specimens were reserved for subsequent experiments.
2.2. Experimental Apparatus
The following instruments and equipment were used in this study: an artificial climate incubator (Model MRX-350B-LED with top-mounted LEDs, manufactured by Ningbo Prante Instrument Co., Ltd., Ningbo, China); a NikonNi microscope equipped with differential interference contrast (DIC) optics, coupled with a NikonDS-Ri2 camera and operated using NIS-Elements D v4.4 software (Nikon Corporation, Shanghai, China); a Nikon SMZ745 stereomicroscope (Nikon Corporation, Shanghai, China); an Haier DW-86L338 ultra-low temperature freezer (Haier Company, Fuzhou, China); handmade dissection needles; and multiple finger tubes (φ = 0.5 cm, h = 3 cm).
2.3. Test Methods
2.3.1. Observation of Parasitization Behavior in T. remus
The parasitization behavior of T. remus was observed by placing a newly emerged (≤3 h old), healthy female individual together with a fresh S. frugiperda egg mass (≤24 h old) inside a finger tube. A strip of filter paper soaked in a 20% honey solution was provided within the tube as a nutritional source. The parasitic behavior was continuously monitored for a duration of 30 min using a stereomicroscope, and photographic documentation was conducted to record the observed behaviors. This procedure was replicated five times.
2.3.2. Parasitism Efficiency of an Individual T. remus Female on Host Eggs at Different Densities
Fresh egg masses of S. frugiperda (≤24 h old) were prepared at densities of 10, 16, 20, 30, 40, and 50 eggs per mass. Newly emerged, healthy female T. remus individuals were collected for the study. Each experimental unit consisted of a single female introduced into a finger tube containing one host egg mass of a designated density. The number of parasitized eggs, identified by characteristic marking behavior, was recorded at two time intervals, 0–10 min and 0–30 min, to determine the parasitism rate. The experiment was conducted at room temperature and replicated five times.
2.3.3. Parasitism Rate of an Individual T. remus Female in Relation to Host Exposure Time
Based on prior findings, a host egg density of 30 eggs per mass was selected as optimal for parasitization and observation. Fresh S. frugiperda egg masses (30 eggs/mass, ≤24 h old) were exposed to single, newly emerged, healthy females of T. remus in finger tubes. These tubes were maintained in an artificial climate chamber set at 26 ± 1 °C, 70 ± 5% relative humidity, with a 14L:10D photoperiod, and a light intensity of 1000 lux. The females were allowed to parasitize the host eggs for designated exposure periods of 1, 1.25, 1.5, 1.75, 2, 2.25, or 2.5 h, after which they were removed. Parasitism was confirmed if, within three days, the host eggs remained plump, turned progressively black, and failed to hatch S. frugiperda larvae. The parasitism rate was calculated and recorded accordingly. The experiment was replicated 10 times for each exposure duration.
2.3.4. Dissection-Based Examination of Host Discrimination by T. remus Toward Previously Parasitized Host Eggs
Fresh S. frugiperda egg masses (30 eggs/mass, ≤24 h old)were parasitized by T. remus females one hour prior to the experiment. A single newly emerged, healthy female was then introduced with one pre-parasitized egg mass into a finger tube at room temperature (26 ± 1 °C). Using a stereomicroscope, the female’s behavior was monitored in real time. Eggs upon which T. remus exhibited either oviposition-marking or probing behavior were specifically identified and marked. Following the completion of these behaviors, the marked eggs were incubated for one day before being dissected. The number of first-instar T. remus larvae present within each dissected host egg was observed and recorded. The experiment was conducted in five replicates, each utilizing a set of 30 dissected eggs.
2.3.5. Self-Discrimination of T. remus Towards Marked Host Eggs Stored at Different Temperatures
Fresh S. frugiperda egg masses (30 eggs/mass) were exposed to individual, newly emerged T. remus females in finger tubes and allowed to parasitize completely under controlled conditions (26 ± 1 °C, 70 ± 5%RH, 14L:10D photoperiod, 1000 lux). Following parasitization, the T. remus individuals and their corresponding parasitized egg masses were separated. The egg masses were then stored for one hour at one of five designated temperatures (16, 21, 26, 31 or 36 °C) within climate chambers. Subsequently, each original T. remus was re-introduced to its respective egg mass in a finger tube. The re-marking behavior exhibited by T. remus towards the previously parasitized eggs was observed under a stereomicroscope at room temperature for a duration of 10 min, during which the average number of re-marked eggs was recorded. Five replicates were performed per temperature.
2.3.6. Conspecific Discrimination of T. remus Towards Marked Host Eggs Stored at Different Temperatures
Following the same procedure for parasitization and temperature treatment as in Section 2.3.5, the stored egg masses were then exposed to one of two types of naive conspecific females in finger tubes: (1) newly emerged females with no prior oviposition experience; (2) newly emerged females with prior oviposition experience. The re-marking behavior towards the parasitized eggs was observed for a duration of 10 min under a stereomicroscope at room temperature, and the average number of re-marked eggs was recorded and calculated. Each female type and temperature condition was tested in five replicates.
2.3.7. Self-Discrimination of T. remus Towards Marked Host Eggs After Different Time Intervals
Fresh egg masses laid by the same S. frugiperda female on the same day were divided into smaller units of 25 eggs each. Each unit (small egg mass) was placed into a finger tube with a single newly emerged T. remus female (1:1 ratio) and allowed to be fully parasitized. Following parasitization, the female wasp and the egg mass were separated. The parasitized eggs were then incubated in a climate chamber (26 ± 1 °C, 70 ± 5% RH, 14L:10D photoperiod, 1000 lux) for different time intervals (0, 1, 2, 4, 6, 10 or 12 h). Subsequently, each original wasp was then re-introduced to its respective time-interval egg mass. The re-marking behavior was observed for a period of 10 min at room temperature using a stereomicroscope, during which the average number of re-marked eggs was recorded and calculated. Five replicates were performed for each time interval.
2.3.8. Conspecific Discrimination of T. remus Towards Marked Host Eggs After Different Time Intervals
Using the same parasitization and time-interval storage procedure as in Section 2.3.7, the stored egg masses were subsequently exposed to one of two types of naive conspecific females: (1) newly emerged females with no prior oviposition experience; (2) newly emerged females with prior oviposition experience. The re-marking behavior of T. remus on parasitized S. frugiperda egg masses was observed using a stereomicroscope at room temperature for a period of 10 min. The number of eggs that were re-marked was recorded and the parasitism rate was calculated. This experiment was replicated five times for each combination of female type and storage time interval.
2.3.9. Data Analysis
Statistical analyses were conducted using SPSS 27.0. One-way analysis of Variance (ANOVA) was performed, and multiple comparison tests, including Duncan’s multiple range test, Tukey’s honestly significant difference (HSD) test, Games–Howell’s post hoc test, Dunnett’s test, and Dunnett’s T3 post hoc test, were applied according to the data characteristics (e.g., variance homogeneity and comparison purpose) to assess significant differences in the following datasets: parasitism efficiency of a single female on host eggs at different densities within 0.5 h, maximum parasitism rate across different exposure times, re-marking behavior on host eggs, and host discrimination (both self and conspecific) under different temperatures and time intervals. Data recording was performed using Microsoft Excel 19. (Microsoft Office, Redmond, WA, USA) and GraphPad Prism 9.5 was used for graph plotting (GraphPad Software, Inc., San Diego, CA, USA).
3. Results
3.1. Parasitization Behavior of T. remus
The oviposition behavior of T. remus encompasses a series of discrete actions, including searching, probing, oviposition, marking, and ovipositor grooming (Figure 1). During the searching phase, the female employs her antennae to continuously tap the surroundings of the host egg mass. Upon locating a host egg mass, she immediately climbs onto it and repeatedly taps each egg with her antennae for assessment. Eggs deemed unsuitable are abandoned in favor of further inspection. When an egg is considered appropriate, the female circles it or pauses to identify an optimal oviposition site using her ovipositor (Figure 1A). In the oviposition stage, if the egg proves unsuitable, the female retracts her ovipositor within a few seconds and resumes searching for other suitable eggs. Conversely, if the egg is suitable, she supports her body with her hind legs, inserts her ovipositor into the host egg, and deposits an egg internally (Figure 1B). Following oviposition, the ovipositor is withdrawn, and the female performs a distinctive “8”-shaped movement with the ovipositor tip on the egg surface to mark the parasitized egg, subsequently grooming the ovipositor (Figure 1C). This characteristic “8” marking serves as a reliable indicator of successful parasitization (Figure 1D). Additionally, it was observed that S. frugiperda egg masses are frequently covered by scales. The female detects interstitial gaps between these scales using her antennae to access the eggs, and may actively remove or displace portions of the scale layer to reach unparasitized eggs prior to probing and ovipositing.
Moreover, T. remus demonstrates the ability to recognize parasitized eggs during the searching phase. Upon encountering an egg that has been previously parasitized or marked, the female promptly avoids or bypasses it without extended inspection. When antennal tapping alone does not provide sufficient information to ascertain whether the egg has been parasitized or marked, she engages in ovipositor probing to assess the internal state of the egg. If the internal conditions are deemed unsuitable, such as the presence of prior parasitism, she retracts her ovipositor and departs. Conversely, if the egg is suitable, she proceeds to oviposit and subsequently deposits an oviposition mark on the egg’s surface.
3.2. Parasitism Efficiency of an Individual T. remus Female on Host Eggs at Different Densities
As shown in Figure 2, the parasitism rate of a single T. remus female gradually increased with host egg density and eventually plateaued. Within 10 min, parasitism efficiency was significantly higher (F = 2.035, p < 0.05) for the 50-egg masses compared to the 10-egg masses. No other significant differences (F = 2.035, p > 0.05) were detected among the other density groups (16, 20, 30, 40 eggs) relative to these two extremes within this period.
Within 30 min, parasitism efficiency at the lowest density (10 eggs) was significantly lower than at densities of 16 and 20 eggs (F = 22.575, p < 0.05), and markedly lower than at densities of 30, 40, and 50 eggs (p < 0.01). Furthermore, the rates for the 16- and 20-egg masses were also significantly lower than those for the 30-, 40-, and 50-egg masses (F = 22.575, p < 0.05). These results indicate that as host density increases, the time required by a single female for host searching significantly decreases, eventually stabilizing at higher densities.
3.3. Parasitism Rate of an Individual T. remus Female in Relation to Host Exposure Time
As shown in Figure 3, the parasitism rate achieved by a single T. remus female increased gradually as the exposure time extended up to 1.5 h, then slightly decreased and eventually stabilized. Except for the comparison with 1.75 h (F = 2.93, p > 0.05), the parasitism rate at 1 h showed significant differences (F = 2.93, p < 0.05) from those at 1.25 h, 1.5 h, 2 h, and 2.5 h. The highest parasitism rate (99.7%) was observed at 1.5 h of exposure, while the lowest rate (95.0%) occurred at 1 h. These results indicate that a parasitism duration of 1.5 h is optimal for a single female T. remus to achieve the highest parasitism rate on a 30-egg mass of S. frugiperda.
3.4. Parasitism Recognition of T. remus Towards Parasitized Host Eggs
T. remus exhibits marking behavior following oviposition on S. frugiperda egg masses, coupled with the capacity to recognize these marks. In the present study, the term “initial marking” denotes the mark deposited immediately after oviposition on fresh, unparasitized eggs. “Re-marking” refers to the marking applied subsequent to a second oviposition event on eggs that have already been parasitized. The term “re-probing” describes the behavior wherein the ovipositor is reinserted into a parasitized egg and then withdrawn without leaving a mark.
Oviposition was consistently accompanied by egg surface marking (Figure 1C,D). Dissection analyses revealed that eggs with initial marks contained no more than one larva, with an initial parasitism rate of 85.4%. Among eggs with re-marks, the distribution was as follows: 23 eggs contained two larvae, 5.8 eggs contained one larva, and 1.4 eggs contained no larvae. When parasitism rates were calculated based on the number of eggs containing two larvae, the rate was 76.6%. Statistical analysis indicated no significant difference (F = 1.417, p > 0.05) between the parasitism rates associated with initial marking and re-marking. Furthermore, no host egg was found to contain more than two larvae, indicating that each marking event corresponds to the deposition of a single egg within the host (Table 2).
Additionally, re-probing behavior was observed. Dissection of the eggs subjected to this behavior revealed that no egg contained two larvae. Among these eggs, 23.8 eggs contained one larva and 6.2 eggs contained none, with a resulting parasitism rate of 79.2%. Statistical analysis revealed no significant differences (F = 1.417, p > 0.05) in parasitism rates among initially marked, re-marked, and re-probed eggs. These findings indicate that re-probing does not result in oviposition and represents an additional form of host discrimination during oviposition.
3.5. Host Recognition by T. remus of Self-Marked Eggs Stored at Different Temperatures
As illustrated in Figure 4, the storage temperature of host eggs significantly influenced the parasitism recognition behavior exhibited by female T. remus. At temperatures of 16, 21, 26, 31, and 36 °C, the mean number of eggs marked during secondary parasitism recognition on treated host-marked eggs was 0, whereas in the CK group (first oviposition marking at 26 °C within 10 min), the mean number of eggs marked was 4.4. These results indicate that self-marked eggs stored between 16 °C and 36 °C consistently elicited host recognition behavior, effectively preventing re-parasitism by T. remus.
3.6. Host Recognition by Conspecific T. remus Females of Marked Eggs Stored at Different Temperatures
The average number of eggs marked by conspecific T. remus females with no prior oviposition experience increased as the storage temperature of the eggs rose (Figure 5). Specifically, the mean quantities of marked eggs after one hour of storage at 16, 21, 26, 31, and 36 °C were 0.8, 3.5, 3.6, 3.4, and 4.2 eggs, respectively. Statistical analysis revealed a significant difference (F = 6.001, p < 0.05) between eggs stored at 16 °C and those stored at temperatures ranging from 21 to 36 °C, as well as the control group (CK), which consisted of eggs marked at 26 °C within 10 min. However, no significant differences (F = 6.001, p > 0.05) were detected among eggs stored at temperatures ranging from 21 to 36 °C or the control. These results indicate that only eggs stored at 16 °C for one hour were effectively recognized by conspecific unmated females, suggesting that the marking pheromone exhibited reduced volatilization at this lower temperature, thereby facilitating recognition.
Conversely, the mean quantity of eggs marked by conspecific females with prior oviposition experience exhibited a decline as temperature increased (Figure 6). The average numbers of marked eggs stored at 16, 21, 26, 31, and 36 °C were 1.6, 1.0, 0.4, 0, and 0 grains, respectively. Each treatment group showed a statistically significant difference (F = 13.081, p < 0.05) compared to the control (CK), although no significant differences were detected among the temperature treatments themselves. These findings imply that experienced females were able to recognize marked eggs across all storage temperatures, with recognition intensity increasing with temperature, indicating that their response may be influenced by pheromone concentration.
3.7. Host Recognition of Self-Marked Eggs by T. remus over Different Time Intervals
T. remus females exhibited the ability to recognize their own marked eggs over various time periods (Figure 7). The average number of eggs remarked at 0, 1, 2, 4, 6, 10, and 12 h were 0, 0.2, 0.4, 0.2, 0.2, 0.2, and 0 grains, respectively. Statistical analysis revealed that all time intervals from 1 to 10 h showed significant differences (F = 47.398, p < 0.05) compared to the (CK),which had 4.4 eggs marked at 26 °C within 10 min. These findings indicate that self-marked eggs were consistently recognized by T. remus females over the examined time.
3.8. Host Recognition by Conspecific T. remus on Marked Eggs over Different Time Intervals
As illustrated in Figure 8, the average number of eggs marked by conspecific unmated females initially increased and then stabilized over time. At 0 h, the mean number of marked eggs (1 egg) was significantly lower (F = 2.146, p < 0.05) compared to the CK (4.4 eggs). The mean values recorded at 1, 2, 4, 6, 10, and 12 h were 2.2, 3.0, 3.6, 3.0, 2.7, and 2.8 eggs, respectively, with no statistically significant differences observed among these intervals. These findings indicate that the recognition ability of conspecific unmated females declined over time, likely due to natural degradation of the host-marking pheromone.
In contrast, experienced conspecific females consistently recognized marked eggs across all examined time intervals (Figure 9). Each time points showed a statistically significant difference (F = 21.774, p < 0.05) compared to the CK. The average quantities of marked eggs detected at 0, 1, 2, 4, 6, 10, and 12 h were 0, 0.4, 0.6, 0.2, 0.6, 0.2, and 0.8 grains, respectively, with no significant differences among intervals. These findings suggest that experienced females retained the ability to recognize marked eggs for up to 12 h, potentially attributable to learning experience associated with prior oviposition behavior.
4. Discussion
The parasitic behavior exhibited by numerous parasitoid wasps encompasses several stages, including host location, host examination, oviposition probing, and host-marking [40,41,42]. Among these behaviors, host-marking serves as an effective behavior to prevent superparasitism. Comparative analysis between the genera Telenomus and Trissolcus confirms that host recognition is widespread in both groups. Most Trissolcus species employ a uniform marking behavior: they drag the ovipositor in a characteristic “8”-shaped pattern over the egg surface to deposit the marking pheromone. Notably, T. remus in this study also employs this same “8”-shaped pattern marking behavior. Within the genus Telenomus, species such as T. busseolae Gahan and T. isis Polaszek, also perform similar abdominal twisting to trace an “8” pattern on the host egg [43], similar to T. remus. In contrast, some Telenomus species exhibit different marking motions, including continuous rubbing along a curved trajectory, lateral sweeping, or drawing irregular circular patterns around the oviposition site. Despite exhibiting greater diversity in marking behaviors than Trissolcus, most Telenomus species nonetheless share the key characteristic of host recognition with the latter. Recognition of these marking behaviors allows parasitoids to avoid intra- and interspecific competition, thereby reducing egg wastage and improving oviposition efficiency. This in turn stimulates parasitoids to search for more unparasitized hosts, which is highly advantageous for biological control programs.
Host density and the duration of parasitization are established factors influencing parasitism rates [44,45]. Research indicates that under conditions of host egg scarcity, T. remus exhibits protective behavior towards parasitized hosts, while prolonged parasitization can lead to superparasitism [3]. To evaluate how these factors affect T. remus, the present study confirms that both are critical in determining its parasitism rate. Short-term oviposition behavior (observed over 10 to 30 intervals) reflects the search efficiency of T. remus. Search time decreased with increasing host density until reaching a plateau, demonstrating a density-dependent “convenience effect”. These findings aligns with Morales-Sánchez et al., who reported that the parasitism rate of T. remus in response to host density follows Holling’s Type II functional response model [46]. A similar pattern has been documented in Meteorus pulchricornis (Wesmael), whose parasitism rate decreased with increasing host density below a threshold of about 8 hosts per plant, and stabilized near 50% beyond that threshold, indicating an upper limit to the parasitoid’s parasitic capacity [47]. Longer parasitization durations (e.g., 1.5 h) reflect the total parasitic capacity of T. remus. At a host density of 30 eggs per egg mass, a 1.5 h exposure achieved the maximum parasitism rate. Shorter exposure times were insufficient to achieve full parasitic potential, while excessively prolonged durations potentially diminished efficiency due to oviposition fatigue and excessive probing behavior. Consequently, a parasitization duration of 1.5 h was identified as optimal and was employed as the standard condition in subsequent experimental procedures.
In natural environments, many parasitoid species possess the ability to discriminate parasitized hosts [11], thereby avoiding oviposition in hosts that have already been parasitized and consequently minimizing intraspecific competition among their offspring [48,49,50]. Research indicates that T. remus exhibits a pronounced host discrimination ability using sensory organs such as antennae and ovipositor to detect parasitized hosts and prevent superparasitism. Behavioral studies corroborate that females typically lay only one egg per host and avoid re-parasitizing the same egg, aligning with observations reported by Xue et al. [3]. Despite this strong discriminatory behavior, the presence of multiple larvae within a single host egg has been observed both in natural and laboratory conditions [3]. However, usually only one wasp larva successfully reaches maturity [51]. This phenomenon may result from intraspecific larval competition or occasional failures in recognizing host-marking pheromones; further research is required to elucidate the underlying mechanisms.
Factors such as prior oviposition experience [52], temperature [53], and time intervals [43,54] can significantly influence parasitoid discrimination behavior. The present study demonstrated that experienced females of T. remus could recognize marked host eggs across a temperature range (16–36 °C) in both self- and conspecific recognition assays, whereas inexperienced females only discriminated hosts marked at 16 °C. These results suggest that lower temperatures may better preserve host-marking substances (e.g., pheromones or cuticular compounds), while higher temperatures (21–36 °C) likely accelerate their degradation or volatilization. Additionally, experienced females may also detect fainter chemical signals, possibly due to sensitized sensory perception. Moreover, experienced females maintained effective recognition of parasitized hosts for up to 12 h, whereas naïve females only recognized eggs marked at 0 h. This implies that experienced T. remus may rely on internal memory (e.g., long-term memory) to compensate for fading external chemical signals. Supporting evidence from Wang et al. [24] reported that T. ostriniae Pang et Chen with one or two oviposition experiences required less time for host searching and oviposition, and rarely needed to probe host eggs with their ovipositors for recognition, in contrast to naive wasps and control groups. Similarly, Trichogramma evanescens Westwood, initially exhibited no innate response to benzyl cyanide, an odor associated with mated Pieris brassicae (Lin.), but after a single oviposition experience, individuals learned to associate this cue with successful parasitism and formed a long-term memory [55]. These findings further support that oviposition experience prolongs the persistence of host discrimination memory. This behavioral difference may reflect an evolutionary strategy whereby experienced females optimize resource utilization and avoid superparasitism through accumulated experience, thereby enhancing offspring survival. Conversely, inexperienced females may rely predominantly on immediate signals to minimize energy expenditure.
Thus, oviposition experience emerges as a primary determinant influencing host discrimination in T. remus females, a characteristic similarly observed in other parasitoid wasps. For instance, oviposition experience significantly enhances the discrimination ability of Trichogramma chilonis Ishii [52]. Individuals with prior oviposition experience demonstrate enhanced discriminatory performance, suggesting that accumulated experience facilitates environmental adaptation and strengthens long-term recognition memory—a crucial advantage for parasitoids characterized by brief lifespans and limited reproductive opportunities [55]. These findings are consistent with the documented roles of learning and memory functions in parasitoids as reported by Lewis and Takasu [56] and Wang et al. [24]. Through experience-enhanced discrimination, T. remus optimizes resource allocation, reduces intraspecific offspring competition, and ultimately increases reproductive success. This behavioral adjustment represents a strategic mechanism to maximize offspring survival.
From the standpoint of biological control, these results offer a foundation for enhancing the deployment strategies of T. remus. Subsequent investigations may delve deeper into molecular mechanisms that underpin this discriminatory capacity, including associated gene expression patterns or neural changes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Prokopy R.J. Epideictic pheromones that influence spacing patterns of phytophagous insects Semiochemicals: Their Role in Pest Control John Wiley & Sons Hoboken, NJ, USA 1981181213
- 2Salt G. Competition among insect parasitoids. Mechanisms in biological competition Proceedings of the Symposium of the Society for Experimental Biology Birmingham, UK 1 December 196196119
- 3Xue T. Pan J. LIUL. Xu T. Huang J. He Y. Wang Z. Intraspecific host discrimination and superparasitism in Telenomus remus (Nixon), an egg parasitoid of Spodoptera frugiperda (J. E. Smith)Chin. J. Biol. Control 20244011110.16409/j.cnki.2095-039x.2023.01.026 · doi ↗
- 4Hardy I.C.W. Briffa M. Introduction to Animal Contests Cambridge University Press Cambridge, UK 20131410.1017/CBO 9781139051248.003 · doi ↗
- 5Zhu X. Liu Q. Xu C.Q. Guo K. Xu R. Qiao H.L. Chen J. Advances in research on sensilla structure andfunction in parasitic wasps Chin. J. Appl. Entomol.20215882284510.7679/j.issn.2095-1353.2021.081 · doi ↗
- 6Eisenstein E. Reep R. Behavioral and cellular studies of learning and memory in insects Compr. Insect Physiol. Biochem. Pharmacol.19859513547
- 7Nufio C.R. Papaj D.R. Host marking behavior in phytophagous insect and parasitoids Entomol. Exp. Appl.20019927329310.1046/j.1570-7458.2001.00827.x · doi ↗
- 8Huang Y.P. Wang S.F. Tang D.W. Ren L.Z. Host discrimination and marking pheromone of Telenomus dendrolimusi J. Cent. South Univ. For. Technol.19931310310810.14067/j.cnki.1673-923x.1993.02.001 · doi ↗