Simulated Heatwaves Affect Development of Two Congeneric Gregarious Larval–Pupal Endoparasitoids
Lizhi Wang, Yanli Zhao, Zhihui Jiao, Baoping Li, Minghui Fei

TL;DR
Short heatwaves reduce the brood size and affect development in two parasitoid wasp species that target ladybirds, which are important for controlling crop pests.
Contribution
This study is the first to investigate the effects of heatwaves on fourth-trophic-level parasitoids.
Findings
Heatwaves reduced brood size in both Oomyzus scaposus and O. spiraculus.
Short heat exposure accelerated development in O. scaposus but reduced adult body mass with longer exposure.
Daily heatwaves of a few hours can negatively affect the performance of both parasitoid species.
Abstract
Climate change is causing more frequent and intense heatwaves, which can disrupt food webs by affecting insects at the top of food chains. This study examined how short-term heatwaves influence two species of parasitoids that attack the larvae and pupae of seven-spot ladybirds, which are important predators of crop pests. We simulated heatwaves of different lengths and measured the performance of the two parasitoids. We found that even brief heatwaves reduced the brood size in both wasp species. For Oomyzus scaposus, short heat exposure sped up development, but longer exposure led to smaller adults. O. spiraculus showed similar patterns, but these were not statistically significant. Overall, the study shows that even short daily heat events can influence the performance of both species. Understanding these effects is important for predicting how climate change may alter predator–prey…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
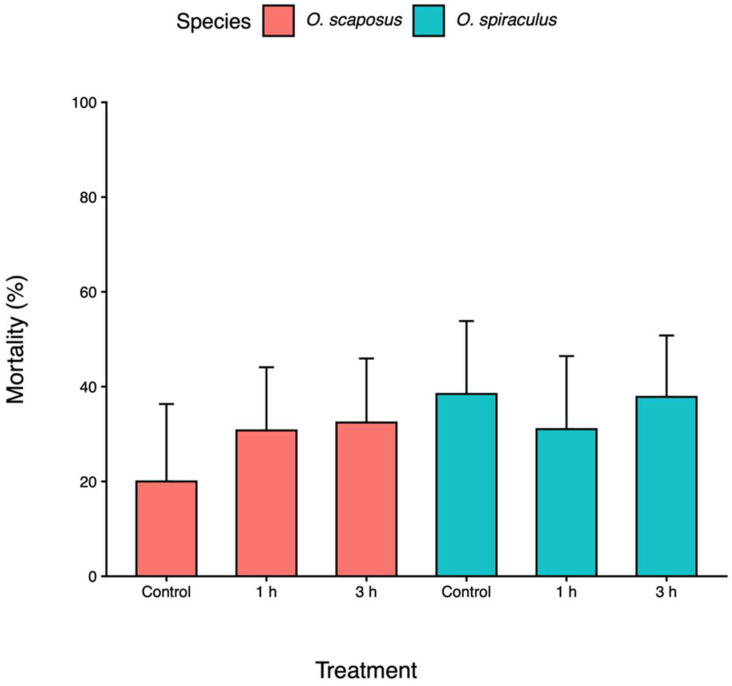 Figure 1
Figure 1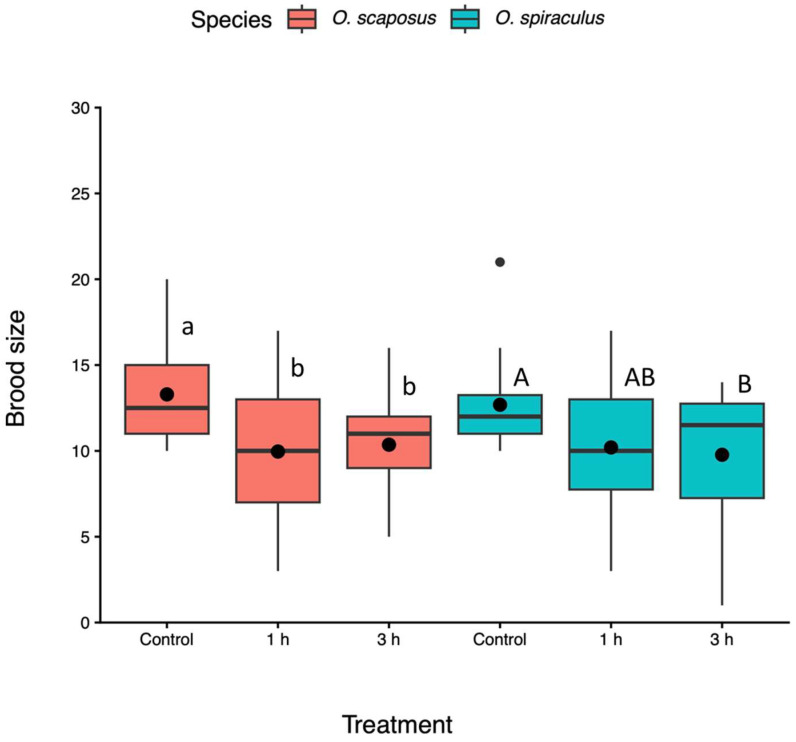 Figure 2
Figure 2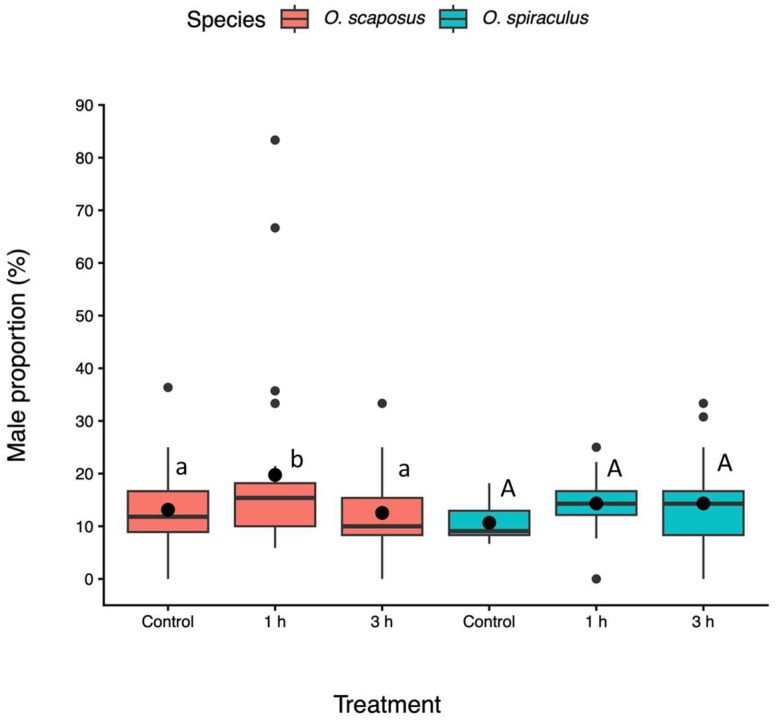 Figure 3
Figure 3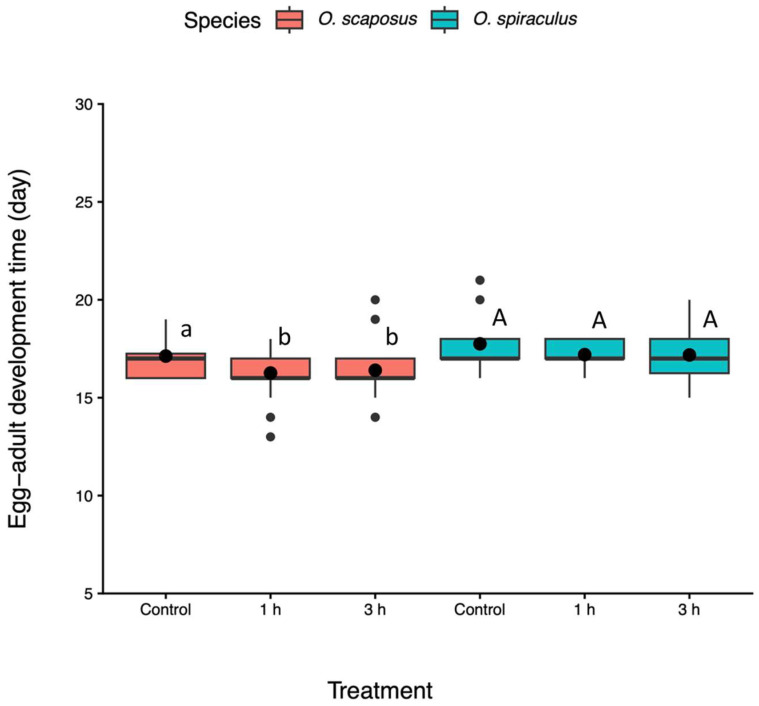 Figure 4
Figure 4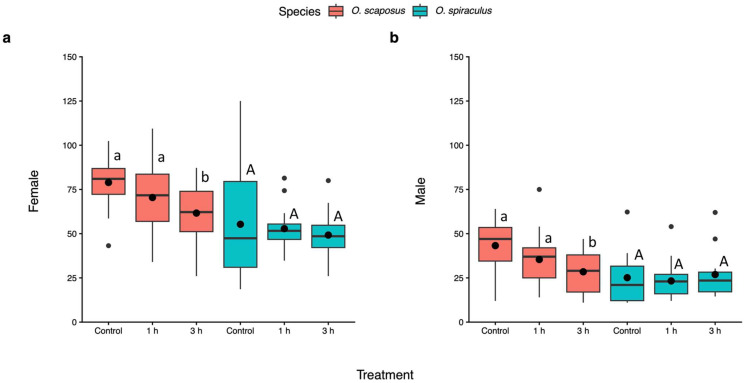 Figure 5
Figure 5- —Joint Research Program of State Key Laboratory of Agricultural and Forestry Biosecurity
- —The National Key Research and Development Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect-Plant Interactions and Control · Physiological and biochemical adaptations · Animal Ecology and Behavior Studies
1. Introduction
Climate change is altering thermal environments globally, resulting in an increase in the frequency and intensity of extreme temperature events [1], while also triggering ecological impacts across multiple scales [2,3]. During these events, daily maximum temperatures can surpass the upper thermal limits of local organisms. Even brief exposures of just a few hours to such high temperatures can be detrimental to ectotherms like insects, whose body temperature depends largely on environmental temperature [4,5,6].
The effect of extreme temperatures on insects can directly affect their physiology and behavior [7,8]. The responses of insect fitness to extreme temperature are likely to be both complex and variable, depending on insect life-history traits and the heat amplitude and heat frequency [9,10,11]. Although species can exhibit variable levels of thermal tolerance, exposure to extreme temperatures often results in a decrease in insect performance [5,12,13], such as growth and survival of immature stages [14], adult lifespan and reproductive success [15,16], and immunity and the accumulation of fat reserves [17].
Parasitoids are insects that deposit their eggs on or in a host organism (usually another insect or arthropod), which they use as a food source for their development, and eventually kill the host [18]. Parasitoid insects are vital top-down regulators of many insect herbivores, and there is a growing body of evidence that they have greater thermal sensitivity and lower thermal tolerance than their hosts [19,20]. Because parasitoids rely on complex physiological mechanisms to survive within their hosts, they are especially vulnerable to increasing temperatures, unpredictably variable temperatures, and stressful temperature events that could disrupt these processes [21]. The impact of extreme temperature is likely even more severe in parasitoids at higher trophic levels that depend on the capacity of those at lower trophic levels to adapt to these changes. However, compared with the parasitoids at the third trophic level, less is known about these effects on parasitoids at the fourth level at the terminal end of food chains [22]. Understanding the effects of climate variability on the fourth trophic level is of fundamental importance to understanding the ecological consequences of elevated temperatures.
The present study investigates the effects of experimental heatwave duration variability on the development of two congeneric gregarious larval–pupal endoparasitoids, Oomyzus scaposus Thomson and O. spiraculus Song, Fei & Cao (Hymenoptera, Eulophidae), of the seven-spot ladybird, Coccinella septempunctata L. (Coleoptera, Coccinellidae). Most ladybird beetles are attacked by primary parasitoids occupying the fourth trophic level [23], and these parasitoids have a potentially fundamental importance in mediating trophic cascades in natural communities. Several parasitoids of C. septempunctata have been previously described in the literature, but the biology, ecology, and developmental parameters of O. scaposus or O. spiraculus are thus far less studied [24]. Given that they are sister species and exhibit significantly overlapping traits, we hypothesize that both species respond to heatwave exposure similarly.
2. Materials and Methods
2.1. Insect Cultures
The prey for the seven-spot ladybirds, vetch aphids, Megoura viciae Mordvilko, originated from a long-standing laboratory colony maintained at the Jiangsu Academy of Agricultural Science. Seven-spot ladybirds, C. septempunctata, and their endoparasitoids, O. scaposus and O. spiraculus, were originally collected from vetch fields near Nanjing, east China. The ladybird colony was established from 30 adult seven-spot ladybirds randomly collected from the field and was maintained in insect rearing cages (40 × 40 × 40 cm) containing beans (Vicia faba L.) infested with vetch aphids as food resources. Colonies of O. scaposus and O. spiraculus were reared on host larvae. In total, 2 or 3 instar ladybird larvae were present at the end of a small brush applied to 2-to-3-day-old female wasps of each species in a plastic vial, and they were considered to be parasitized when the female wasp inserted her ovipositor into the host for at least 30 s. The parasitized larvae were then reared in a Petri dish (diameter of 5 cm) with vetch aphids that were replenished daily, and they were individually transferred into a glass tube after pupation. All insects were maintained in a climate room at a temperature of 22 ± 2 °C, a relative humidity of 50–70%, and a photoperiod of 16 L:8 D.
2.2. Experimental Design
Since in one of our previous thermal effects studies, we found that all parasitized seven-spot ladybirds (n = 33 for O. scaposus, n = 37 for O. spiraculus) had died at the pupal stage under an extremely high temperature of 33/28 °C; we dissected the pupae that have been found to be more than 90% parasitized but from which adult wasps could not successfully emerge. Therefore, we set a temperature of 33 °C to investigate the effect of variable heatwave duration on the performance of the two parasitoid wasps. We exposed the parasitized ladybird larvae to different heatwave (33 °C) durations using incubators (RGLC-P400-C3, Darth Cater, Hefei, China). The incubators were set with regular day–night temperatures of 22/20 °C, a relative humidity of 50–70%, and a photoperiod of 16:8 h L:D. In the middle of the daytime, the parasitoids were exposed to a 1 h or 3 h heatwave of 33 °C through their whole development period until adult wasps egressed. A group of parasitoid wasps that did not undergo any heat exposure served as the control.
A female wasp (3 days old) with a similar body size from each species was allowed to parasitize the host larvae once until leaving the host voluntarily. Each female wasp was used once. Each parasitized larva was transferred into a glass tube and put in incubators with a variable heatwave duration temperature with a relative humidity of 50–70%. The parasitized larvae were provided with vetch aphids until the adult wasps emerged. The development time from oviposition to adult eclosion, the number of emerging wasps (brood size), and the sex ratio (proportion of male adult parasitoids) were recorded. At eclosion, one male and five female wasps from each brood were dried in an oven at 60 °C for 72 h and weighed on a microbalance. The mortality rate (proportion of hosts that could not give rise to wasps) was also measured.
2.3. Statistical Analysis
Data were analyzed per parasitoid species and per sex if applicable (biomass). Data on mortality, brood sizes, sex ratio, adult body mass, and egg-to-adult development time were analyzed using general or generalized linear models depending on the distribution of the data. Mortality refers to individual trial failure (i.e., a host did not produce adult parasitoids). Brood sizes refer to the realized brood sizes, i.e., the number of wasps that successfully developed into adults. Adult body mass was the mean biomass of the parasitoids eclosed from the same host individual. All wasps from each host eclosed on the same day, resulting in a single data entry per host for development time. Sex ratio refers to the proportion of males per brood. Adult biomasses were log-transformed to meet assumptions of normality and homoscedasticity. In all statistical models, parasitoid species, temperature treatment, and their interactions served as explanatory variables. Mortality was analyzed using a GLM with a quasi-Binomial distribution and a logit link function. Brood size and development time were analyzed using a GLM with a quasi-Poisson distribution and a log link function to correct for overdispersion of the data. Sex ratio was analyzed using a quasi-Binomial distribution (correcting for overdispersion) and a logit link function. Post hoc Tukey HSD tests were performed to compare means when any of the model terms were significant. Statistics are provided when each of the main effect terms is entered last (Type III effects). All statistical analyses were undertaken with the R statistical software (version 4.4.3) [25].
3. Results
Mortality. Mortality was neither affected by parasitoid species (F_1, 248_ = 0.43, p = 0.25), heatwave duration (F_2, 248_ = 0.43, p = 0.65), nor their interaction (F_2, 248_ = 0.34, p = 0.71). In general, mortality ranged between 20 and 37% (Figure 1).
Brood sizes. The heatwave duration had a significant effect on parasitoid brood size (F_2, 167_ = 6.12, p < 0.01), and this effect was similar for the two parasitoid species (species: F_1, 155_ = 1.75, p = 0.18; heatwave–species interactions: F_2, 155_= 0.58, p = 0.56). The heatwave treatment negatively affected the brood size of O. scaposus (Tukey test: p < 0.05, Figure 2), and there were no significant differences between the 1 and 3 h treatments (Tukey test: p > 0.05). O. spiraculus had a similar pattern; the brood size under the 3 h treatment was smaller than that under the control treatment (Tukey test: p < 0.05, Figure 2), and there were no significant differences between the 1 and 3 h treatments (Tukey test: p > 0.05).
Sex ratio. The sex ratio of both species was highly female-biased (Figure 3). The sex ratio of parasitoid offspring was affected by the heatwave treatment depending on the parasitoid species (heatwave: F_2, 21_ = 6.76, p < 0.01; species: F_1, 19_ = 0.13, p = 0.72; heatwave–species interactions: F_2, 20_ = 3.86, p = 0.023). For O. scaposus, the male proportion initially increased and then decreased as the heatwave duration increased (Tukey test: p < 0.05, Figure 3). The proportion of male O. spiraculus increased with heatwave duration, but this was not statistically significant (Tukey test: p > 0.05, Figure 3).
Development time. All males and females of the two parasitoid species eclosed on the same day from a single host. Egg-to-adult development time varied depending on the temperature treatment (F_2, 12_ = 3.70, p = 0.027) and parasitoid species (F_1, 12_ = 4.71, p = 0.032), but it was not affected by their interactions (F_2, 11_ = 0.067, p = 0.94). The development of O. spiraculus was longer than that of O. scaposus (Tukey test: p < 0.01, Figure 4). Heatwave exposure for 1 h could accelerate the development of O. scaposus (Tukey test: p < 0.05, Figure 4), and there were no significant differences between the 1 and 3 h treatments (Tukey test: p > 0.05). O. spiraculus had a similar pattern, but this was not statistically significant (Tukey test: p > 0.05, Figure 4).
Adult body mass. Both female and male adult biomass were affected by the parasitoid species (female: F_1, 187_ = 22.40, p < 0.01; male: F_1, 183_ = 19.08, p = 0.01). The female adult body mass was also affected by the heatwave treatment (heatwave: F_2, 172_ = 7.03, p < 0.01; species–heatwave: F_2, 163_ = 1.70, p = 0.19). The heatwave treatment affected male adult body mass depending on the parasitoid species (heatwave: F_2, 176_ = 1.38, p = 0.26; parasitoid–heatwave interactions: F_2, 170_ = 4.67, p = 0.011). Both male and female wasps of O. scaposus were heavier than those of O. spiraculus (Tukey tests: p < 0.05, Figure 5). For O. scaposus, the body mass of both females and males was the lowest under the 3 h heatwave treatment (Tukey test: p < 0.05, Figure 5), whereas there were no significant differences between any other heatwave treatment comparisons. For O. spiraculus, the heatwave marginally negatively affected both female and male adult body mass, which was not statistically significant (Tukey test: p > 0.05, Figure 5).
4. Discussion
Our study provides clear evidence that transient, recurring heatwaves can significantly affect the performance of two congeneric endoparasitoids, O. scaposus and O. spiraculus. While mortality remained unaffected, the observed reductions in brood size for both species and changes in development time and adult body mass for O. scaposus suggest that heat stress can impose substantial sublethal fitness costs. These findings align with a growing body of literature demonstrating that the effects of extreme temperatures on insects are often more pronounced in sublethal performance traits than in direct survival [5,13].
The most consistent negative effect across both species was the reduction in brood size under heatwave conditions. This decline in the number of offspring reaching adulthood could be due to a number of underlying mechanisms, including a reduced egg hatch rate or increased mortality in developing larvae within the host under heat stress [6,14]. Heat stress reduces brood size, which directly influences population growth and stability. This is particularly critical for parasitoids at the fourth trophic level, like Oomyzus spp., which are already predicted to be more vulnerable to climate change due to their reliance on lower trophic levels and complex physiological interactions with their hosts [19,20]. Therefore, the heatwave effect on brood size, rather than mortality, may pose a greater threat to population persistence in the long term.
Local Mate Competition (LMC) theory is fundamental to understanding parasitoid ecology, predicting a female-biased sex ratio to minimize mating competition among male offspring—a pattern confirmed in both studied species. The highly female-biased sex ratio observed in both species is consistent with the expectations of LMC theory, which is common in gregarious parasitoids [26]. Even though the male proportion of O. scaposus significantly increased with the 1 h heatwave, it was still female-biased. One possible explanation for this result is differential thermal sensitivity between the sexes; however, the underlying mechanism requires further investigation. Furthermore, this observed increase in male proportion could contribute to a reduction in population growth potential, as fewer females are available for reproduction. Future studies should further investigate the ecological consequences of such sex ratio shifts under heat stress, including potential impacts on population dynamics. Insects are ectothermic animals, and their body temperature is easily influenced by the ambient environmental temperature. Previous studies have shown that high temperatures can accelerate the growth and development of insects [27,28]. In this study, we also found the accelerated development of O. scaposus even under a daily 1 h heatwave during its development. However, this accelerated development came at a cost, as evidenced by the significantly reduced adult body mass under the 3 h heatwave duration. O. spiraculus tended to have a similar pattern, but this was not statistically significant. This trade-off between development time and body size is a classic life-history constraint [29]. A smaller body size in parasitoids is often correlated with reduced fat reserves, shorter lifespan, and lower fecundity [30,31], which could undermine the apparent short-term benefits of faster development.
In conclusion, our results demonstrate that even short daily exposure to heatwaves can significantly impact the performance of two congeneric parasitoid wasps, the effects of which were more pronounced in sublethal performance traits than in direct survival. Given the crucial role of these parasitoids at the fourth trophic level in regulating ladybird populations and potentially mediating trophic cascades [24], a decline in their performance due to increasing heatwave events could disrupt the stability of this multi-trophic system. Consequently, future research should prioritize quantifying these sublethal effects across different trophic levels to better forecast the full ecological footprint of a warming climate.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lee H. Calvin K. Dasgupta D. Krinner G. Mukherji A. Thorne P. Trisos C. Romero J. Aldunce P. Barret K. IPCC 2023: Climate Change 2023: Synthesis Report, Summary for Policymakers Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change Core Writing Team Lee H. Romero J. IPCC Geneva, Switzerland 2023
- 2Kingsolver J.G. Buckley L.B. Evolution of plasticity and adaptive responses to climate change along climate gradients Proc. R. Soc. B 20172842017038610.1098/rspb.2017.0386 PMC 556379228814652 · doi ↗ · pubmed ↗
- 3Bailey L.D. van de Pol M. Tackling extremes: Challenges for ecological and evolutionary research on extreme climatic events J. Anim. Ecol.201685859610.1111/1365-2656.1245126433114 · doi ↗ · pubmed ↗
- 4Vasseur D.A. De Long J.P. Gilbert B. Greig H.S. Harley C.D.G. Mc Cann K.S. Savage V. Tunney T.D. O’Connor M.I. Increased temperature variation poses a greater risk to species than climate warming Proc. R. Soc. B 20142812013261210.1098/rspb.2013.2612 PMC 392406924478296 · doi ↗ · pubmed ↗
- 5Colinet H. Sinclair B.J. Vernon P. Renault D. Insects in fluctuating thermal environments Annu. Rev. Entomol.20156012314010.1146/annurev-ento-010814-02101725341105 · doi ↗ · pubmed ↗
- 6Ma C.-S. Ma G. Pincebourde S. Survive a warming climate: Insect responses to extreme high temperatures Annu. Rev. Entomol.20216616318410.1146/annurev-ento-041520-07445432870704 · doi ↗ · pubmed ↗
- 7Iltis C. Louapre P. Vogelweith F. Thiéry D. Moreau J. How to stand the heat? Post-stress nutrition and developmental stage determine insect response to a heat wave J. Insect Physiol.202113110421410.1016/j.jinsphys.2021.10421433662375 · doi ↗ · pubmed ↗
- 8González-Tokman D. Córdoba-Aguilar A. Dáttilo W. Lira-Noriega A. Sánchez-Guillén R.A. Villalobos F. Insect responses to heat: Physiological mechanisms, evolution and ecological implications in a warming world Biol. Rev.20209580282110.1111/brv.1258832035015 · doi ↗ · pubmed ↗