Plant-Derived Agents and Systemic Sclerosis: A Systematic Review of Therapeutic Potential and Molecular Mechanisms
Cristian-Mihai Ilie, Teodora-Cristiana Grădinaru, Cătălina Anamaria Boromiz, Marilena Gilca

TL;DR
This review explores plant-based compounds that may help treat systemic sclerosis by targeting key disease processes like fibrosis and inflammation.
Contribution
The study systematically identifies phytocompounds and plant extracts with potential therapeutic effects in systemic sclerosis and predicts new molecular targets.
Findings
24 phytocompounds and 5 plant extracts were found to modulate SSc pathogenic processes like TGF-β/Smad signaling.
93 new molecular targets were predicted, with 41 showing relevance to SSc pathogenesis.
Phytochemicals showed effects on endothelial to mesenchymal transition, oxidative stress, and inflammation.
Abstract
Systemic sclerosis (SSc) is a rare multisystemic autoimmune disease associated with progressive fibrosis, vasculopathy, and immune dysregulation. Despite advances in its management, the disease remains associated with substantial morbidity and mortality, with limited therapeutic options. This systematic review aimed to identify phytocompounds and medicinal plants that had demonstrated efficacy in SSc. A comprehensive literature search was performed in PubMed and ScienceDirect, yielding 7797 records, of which 32 studies met the inclusion criteria. A second search was performed using the SwissTargetPrediction tool to identify new putative molecular targets for these phytocompounds, whose relevance for SSc pathogenesis was verified by a third search in PubMed and ScienceDirect databases. Our search found 24 phytocompouds (e.g., halofunginone, crocetin, and tanshinone IIA) and 5 plant…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1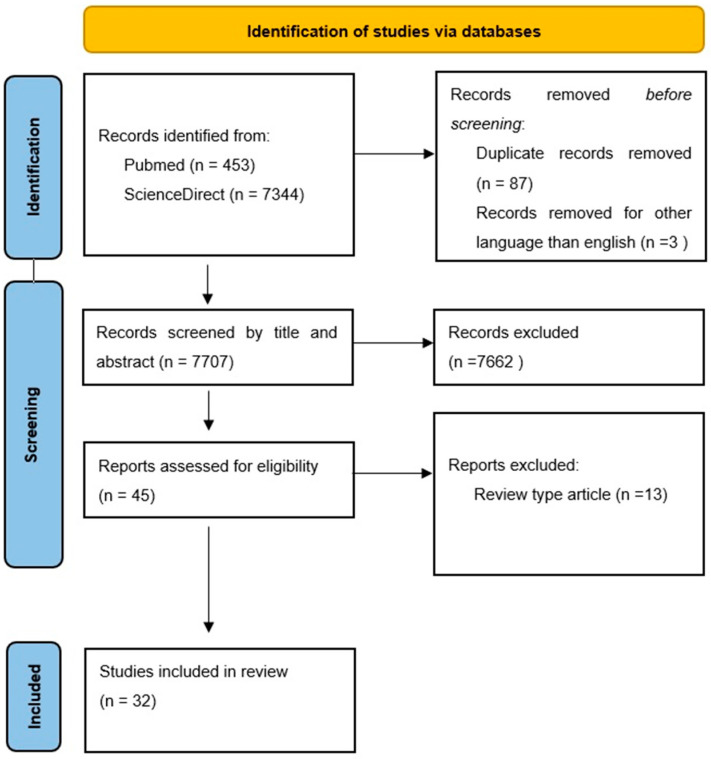 Figure 2
Figure 2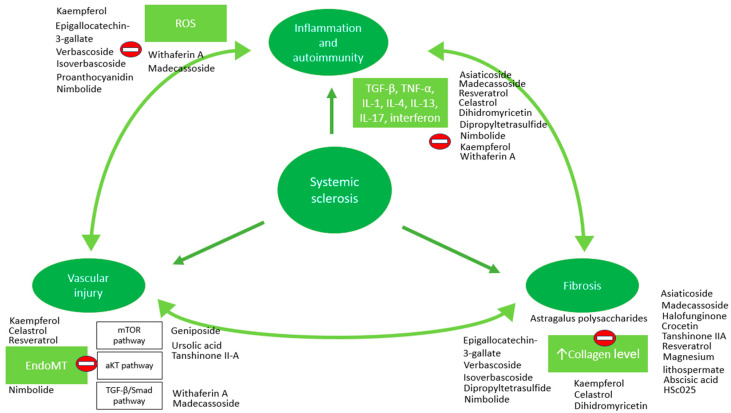 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSystemic Sclerosis and Related Diseases · Saffron Plant Research Studies · Medicinal Plants and Bioactive Compounds
1. Introduction
Systemic sclerosis (SSc) is a complex autoimmune connective tissue disorder, which is characterized by progressive fibrosis of skin and internal organs and microvascular damage [1,2].
The global incidence of SSc is estimated at 1.4–8.6 cases per 100,000 person-years, with a prevalence range from 17.6 to 18.9 per 100,000 individuals. Although mortality at younger ages has declined, the overall mortality still remains elevated compared to the general population. The leading causes of death in SSc patients are interstitial lung disease (ILD) and pulmonary arterial hypertension (PAH) [3].
SSc is a clinically heterogeneous disease characterized by a wide spectrum of manifestations, including cutaneous (progressive skin thickening with sclerodactyly, microstomia, and joint contractures leading to deformities), vascular (Raynaud phenomenon, cutaneous telangiectasia, and ischemic digital ulcers), and visceral (e.g., cardiac, respiratory, renal, and gastrointestinal) manifestations [4].
The pathogenesis of SSc is currently understood as a triad consisting of endothelial injury, immune system activation with the production of specific autoantibodies, and progressive fibrosis, although its exact mechanisms still remain unclear [5].
According to the updated 2023 European Alliance of Associations for Rheumatology (EULAR) treatment recommendations, the management of SSc is individualized and tailored to organ-specific involvement. In Raynaud phenomenon, first-line therapy consists of dihydropyridine-type calcium channel antagonists (most commonly Nifedipine), with escalation to phosphodiesterase-5 (PDE5) inhibitors or intravenous Iloprost in refractory cases. Digital ulcers are treated with PDE5 inhibitors, intravenous Iloprost, or with the endothelin receptor antagonist—Bosentan. PAH requires early initiation of combination therapy with PDE5 inhibitors and endothelin receptor antagonists, additional prostacyclin analogues (e.g., Selexipag) or Riociguat where indicated, with consideration of Epoprostenol for patients with severe PAH. Scleroderma renal crisis mandates the prompt initiation of angiotensin-converting enzyme inhibitors. Gastrointestinal manifestations are addressed with proton pump inhibitors for reflux, prokinetic agents for dysmotility, and cyclical antibiotics in the setting of small intestinal bacterial overgrowth. Cutaneous fibrosis may be addressed with immunosuppressive therapies such as Methotrexate, Mycophenolate mofetil, Rituximab, or Tocilizumab. Management of ILD involves immunosuppressive therapies such as Mycophenolate mofetil, Cyclophosphamide, Rituximab, or Tocilizumab, and antifibrotic treatment with Nintedanib. Musculoskeletal manifestations are generally addressed through treatment with Methotrexate [6].
Despite substantial progress in the management of SSc over the past few decades, the disease continues to be associated with considerable morbidity and mortality. Current management protocols have greatly improved survival in SSc-related ILD and PAH, but they are still limited regarding Raynaud phenomenon, digital ulcers, cardiac, gastrointestinal, or renal involvement. In addition, a subset of patients shows limited responsiveness to existing therapeutic options and continues to demonstrate disease progression despite the appropriate therapy [7].
A survey on herbal therapies conducted with patients with SSc reported that 9 of the 23 patients used a variety of phytotherapy products (essential oils, herbal tea, vegetable oils, or gemmotherapy). On a rating scale of 1–10, the perceived effectiveness of these treatments was an average of 7.2 [8]. In light of these findings, this research sought to evaluate the available data on the therapeutic potential of plant-derived agents in SSc and to summarize the putative molecular mechanisms underlying their biological activity.
2. Materials and Methods
The first step of our analysis consisted of a comprehensive search of the scientific literature, which was conducted in PubMed and ScienceDirect, encompassing publications from January 1975 through August 2025. Each study we found was carefully reviewed and assessed for eligibility following the PRISMA 2020 guidelines to ensure a transparent and systematic selection process [9]. Our review was focused on molecular mechanisms of plant-derived agents, and the results were synthesized using a narrative approach supported by tabular summaries; consequently, given these aspects and the absence of quantitative data synthesis or meta-analysis, the review protocol was not registered in the International Prospective Register of Systematic Reviews (PROSPERO).
The inclusion criteria for this review were established though a systemic search strategy combining various keywords including (“systemic sclerosis” OR “systemic scleroderma” OR “scleroderma” OR “sclerosis”) AND (“plants” OR “herb” OR “phytocompound” OR “phytochemicals”) NOT (“multiple sclerosis” OR “fungus” OR “lateral sclerosis” OR “tuberous sclerosis”). Additionally, a linguistic eligibility criterion was implemented to include only studies published in the English language. Search strings applied across PubMed and ScienceDirect are provided in the Supplementary Materials.
The exclusion criteria encompassed duplicate publication, studies addressing a natural compound but not related to SSc, research on SSc unrelated to natural compound and articles referring to herbal remedies from traditional medicine that had not been scientifically evaluated for SSc.
The main characteristics of the studies included in this systematic review are presented in Supplementary Table S1. An abbreviated version of the Standard Quality Assessment Criteria by Kmet et al. [10] was used to evaluate the methodological quality of all studies. Risk of bias was assessed according to study design, using the Systematic Review Centre for Laboratory animal Experimentation (SYRCLE) Risk of Bias tool [11] for in vivo studies and the Risk Of Bias In Non-randomized Studies-of Interventions (ROBINS-I) tool for human studies [12]. The assessments were performed by one reviewer and independently checked by a second reviewer to minimize bias. The results are summarized in Supplementary Tables S2–S4.
Following the systematic literature search, we extracted all evidence-based molecular targets involved in SSc pathogenesis and generated a synthesized table summarizing the reported interactions and activities of the identified phytocompounds on these targets.
Next, we performed a second search by using the SwissTargetPrediction web tool (http://www.swisstargetprediction.ch/, accessed on 15 October 2025) to identify new molecular targets of the bioactive phytocompounds that were found in our first search.
SwissTargetPrediction is a bioinformatic tool developed to predict ligand–target affinity for small molecules [13]; therefore, the phytochemicals with molecules that are too large, with more than 200 characters per SMILES (e.g., asiaticoside and madecassoside), or enzymes (e.g., bromelain) could not be submitted to the SwissTargetPrediction tool. Furthermore, no target predictions were produced by SwissTargetPrediction for the phytochemical extracts (e.g., proanthocyanidins and astragalus polysaccharides). The target predictions were sorted from high to low according to probability score. Subsequent analyses were limited to those targets with a SwissTargetPrediction probability score above 0.5. The greater the probability score, the more accurate the predicted target; therefore, a probability above 0.5 is commonly set as the threshold to filter more credible targets in network pharmacology studies [14,15,16,17].
We also excluded from this list any molecular targets for which the designation “by homology” appeared after the target name, in order to mi minimize potential false-positive predictions.
We listed all the putative molecular targets of the phytochemicals. Afterwards we have compared this list with the list of evidence-based targets from the first search.
Finally, a third search was conducted in PubMed and ScienceDirect to determine whether these newly identified putative molecular targets of the bioactive phytochemicals may play a demonstrable role in the pathogenesis of SSc, by using the phrase: [(name of the molecular target) AND (“systemic sclerosis” OR “systemic scleroderma”, “scleroderma” OR “sclerosis”) NOT “multiple sclerosis” NOT “fungus” NOT “lateral sclerosis” NOT “tuberous sclerosis”].
The workflow of the study, organized into three phases, is summarized in Figure 1.
3. Results
The literature search identified 453 studies on PubMed and 7344 studies on ScienceDirect. Titles and abstracts were screened to remove duplicates and irrelevant records. Duplicates, irrelevancy, or unreliability resulted in exclusion of 7765 articles. Lastly, 32 studies (original or research articles) were selected for the review and were included in the synthesized tables (Table 1, Table 2, Table 3, Table 4, Table 5 and Table 6). The study selection process is illustrated using a PRISMA 2020 flow diagram (Figure 2), and a complete PRISMA checklist is provided [9].
A structured synthesis of the principal phytocompounds evaluated, their molecular targets, and the corresponding levels of evidence, stratified by study design, is provided in Table 7, allowing a clear and systematic overview of the available data.
To integrate the findings of our literature analysis, we compiled a list of evidence-based molecular targets of the selected phytocompounds relevant to SSc pathogenesis and developed a summary table detailing the documented action of the corresponding bioactive phytochemicals, resulting in a comprehensive color-coded phytocompound-target map covering 68 molecular targets involved in SSc pathogenesis (Supplementary Table S5). In this map, green-marked entries correspond to evidence-based molecular targets (e.g., direct inhibition or binding), orange entries indicate an evidence-based effect on genetic expression (e.g., modulation of mRNA levels), and blue entries denote an evidence-based modification of molecular levels (e.g., plasma or tissular levels).
To further extend the analytical scope of these findings, all phytocompounds were subsequently analyzed using the SwissTargetPrediction web tool, resulting in the identification of an additional set of 93 predicted molecular targets (Supplementary Table S6). In Supplementary Table S6, only the first two columns generated by SwissTargetPrediction were retained: the molecular target name and the common name (linked to GeneCards) [13]. Notably, only one target from the initial evidence-based list derived from the primary literature search (MAPK p38α) was also predicted by the SwissTargetPrediction.
The predicted molecular targets identified using the Swiss Target Prediction analysis (which were not listed in the first phase, except for MAPK p38 alpha) were subjected to a third literature search. This search confirmed the relevance of 43 out of the 93 predicted targets to SSc pathogenesis (Table 8).
4. Discussion
In the context of autoimmunity, anti-centromere, anti-Th/To, and anti-topoisomerase I antibodies are recognized as classical biomarkers of SSc, reflecting disease-associated immune reactivity directed against nuclear components [105]. Upon exposure to pathological stimuli (TGF-β, TNF-α, IL-1, interferon, endothelin-1—ET-1, reactive oxygen species—ROS, hypoxia), endothelial cells undergo structural and functional changes, a process known as endothelial-to-mesenchymal transition (EndoMT) [106]. Through this process, endothelial cells adopt a myofibroblast phenotype, acquiring invasive properties, upregulating mesenchymal markers (α-smooth muscle active—α-SMA, CD44, N-cadherin, vimentin, smooth muscle 22—Sm22, and fibroblast-specific protein-1—FSP-1) and producing collagen [106]. Moreover, myofibroblasts could exacerbate the inflammation by upregulating adhesion molecules, thereby facilitating interactions with circulating immune cells, and by secreting growth factors and cytokines [107]. Fibrosis results from the excessive deposition of collagen and other extracellular matrix proteins by myofibroblasts, occurring in the skin, lungs, gastrointestinal tract, and other vital organs [108]. Subendothelial deposition of fibrous tissue promotes abnormal vascular remodeling, which increases capillary fragility and thereby sustains immune response activation [109]. Therefore, it can be stated that the three components of the pathogenic triad in SSc are in a constant and dynamic interrelationship (Figure 3).
There are several pathways involved in EndoMT, such as Smad-dependent and independent TGF-β pathways [110], mammalian target of rapamycin pathway (mTOR) [111], and protein kinase B pathway (aKT) [112].
During the EndoMT process, the cell loses endothelial cell biomarkers (VE-cadherin and CD31) [113] and upregulates the expression of mesenchymal cell biomarkers (α-SMA, N-cadherin, vimentin, FSP-1) [106]. Thus, as summarized in the results tables, the following phytocompounds have been reported to exert inhibitory effects on the EndoMT process: tanshinone II-A [21], resveratrol [22], geniposide [25], ursolic acid [37], nimbolide [41], withaferin A [42], kaempferol [31], celastrol [32], madecassoside [44], and 5-(tert-Butyl)-N-(1-hydroxy-2-methylpropan-2-yl)-1-(5-(trifluoromethyl)pyridin-2-yl)-1H-pyrazole-3-carboxamide [46]. Additionally, the ethanol extract of Capparis spinosa L., which contains flavonoids such as kaempferol, also induces inhibition of the EndoMT process [49]. Since no human studies have been reported for any of the aforementioned phytocompounds, further investigation of this mechanism in patients with SSc appears warranted.
Angiotensin II has been identified as a key inducer of EndoMT, contributing to fibrotic process progression [114]. Previous studies demonstrated that angiotensin II activate TGF-β signaling pathway, thereby promoting EndoMT and amplifying the inflammatory response through enhanced myofibroblast accumulation. This pathological cascade leads to excessive collagen synthesis and deposition in the skin [115]. Furthermore, co-expression of endothelial and mesenchymal markers in patients with interstitial lung disease (ILD) was observed, suggesting that EndoMT was involved in pathogenesis of ILD in SSc patients [116]. All these findings suggest that EndoMT has a role in the progression of fibrosis in SSc.
Across most studies, antifibrotic efficacy was assessed through the quantification of collagen. According to the data summarized in the tables, various phytocompounds have demonstrated antifibrotic activity in SSc models, including halofunginone [18], crocetin [19], magnesium lithospermate [23], dipropyltetrasulfide [24], tanshinone IIA [20,21], resveratrol [22], absicisic acid [28], HSc025 [30], epigallocatechin-3-gallate [33], curcumin [36], verbascoside [38], isoverbascoside [38], nimbolide [41], kaempferol [31], dihydromyricetin [34], astragalus polysaccharide [43], madecassoside [44], asiaticoside [45], and 5-(tert-Butyl)-N-(1-hydroxy-2-methylpropan-2-yl)-1-(5-(trifluoromethyl)pyridin-2-yl)-1H-pyrazole-3-carboxamide [46]. In addition, Capparis spinosa L., which contains flavonoids, such as Kaempferol, induces a similar effect [49].
pSmad2/3 is a key protein involved in TGFβ induced fibrosis [117]. Our search revealed that curcumin [36], verbascoside [38], isoverbascoside [38], dipropyltetrasulfide [24], nimbolide [41], withaferin A [42], and madecassoside [44] inhibit the phosphorylation, and therefore the activation of Smad2/3.
In the context of fibrosis, additional plant-derived agents have shown promising beneficial effects. Tanshinone II-A has already been traditionally used for treating patients with pulmonary [118] and liver fibrosis [119,120]. Results show that Tripterygium wilfordii Hook.f. confers pulmonary benefits evidenced by increases in mean FVC and FVC % of predicted, when it is used as a maintenance therapy. Considering that both in vivo and in vitro studies have demonstrated beneficial effects on pulmonary fibrosis for HSc025 [30], verbascoside [38], isoverbascoside [38], madecassoside [44], and asiaticoside [45], further investigation in patients with SSc and pulmonary involvement represents a novel and promising research direction.
Multiple phytocompounds (asiaticoside [45], resveratrol [22], celastrol [32], dihydromyricetin [34], dipropyltetrasulfide [24], nimbolide [41], withaferin A [42], kaempferol [31], madecassoside [44]) exerted anti-inflammatory effects in SSc models. One of the key players in the inflammation and fibrosis branches of the SSc pathogenesis triad, TNF-α, contributes to fibroblast activation and its expression has also been linked to the progression and severity of scleroderma [121].
In addition to the previously mentioned isolated phytocompounds, a few plant-derived whole or fractionated extracts were also tested, some proving various degrees of clinical efficacy even in human studies (e.g., caper bush extract [48,49], Lei Gong Teng [51]), while others not (e.g., St. John’s Wort [53]). Regarding St. John’s Wort, although this plant has not been demonstrated to improve the parameters of Raynaud’s phenomenon attacks [53], its content in flavonoids (kaempferol, luteolin) and tannins (proanthocyanidins) [52], which are already found to be bioactive in SSc, and its antioxidant potential [122], warrant further investigation in patients with SSc.
Evening primrose (Oenothera biennis L., family Onagraceae) seed oil is a rich source of gamma-linoleic acid [123]. Supplementation with Oenothera biennis L. oil in patients with SSc may alleviate pain in the hands and feet, promote digital ulcer healing, and improve telangiectasias and skin texture. The therapeutic effects are presumed to result from gamma-linoleic acid, which functions as a metabolic precursor of prostaglandin E1 [124]. Evidence regarding its efficacy in Raynaud phenomenon remains inconclusive [54,55].
Extract of Ginkgo biloba L. (family Ginkgoaceae), rich in flavonoids [125], has been demonstrated to attenuate endothelial-monocyte adhesion by reducing TNF-α-induced intracellular formation of reactive oxygen species (ROS) in human aortic endothelial cells [126]. Taking into account that the potent antioxidant capacity of flavonoids contributes to their therapeutic effect in oxidative stress-related dermatoses [127], it would be a reasonable assumption that this mechanism may also be involved in case of Ginkgo biloba L. use as an anti-SSC remedy. Elevated ROS contributes to tissue fibrosis by upregulating collagen type I and tissue inhibitor of metalloproteinase gene expressions and reducing the degradation of ECM [128]. Under these circumstances, future research investigating the effects of Ginkgo biloba L. on fibrotic processes in SSc appears warranted.
Centella asiatica (L.) Urb., a member of the Apiaceae family, contains a high amount of triterpene glycosides, primarily asiaticoside and madecassoside, which are the main bioactive compounds. Consequently, Centella asiatica extracts display notable antioxidant and anti-inflammatory properties, largely attributable to asiaticoside, asiatic acid, and madecassoside. These compounds mitigate oxidative stress by reducing malondialdehyde levels and enhancing glutathione levels [129].
Recent studies have demonstrated that the endocannabinoid system can modulate dysregulated mechanisms implicated in SSc pathogenesis, including fibrosis, inflammation, and vascular tone. Over 60 phytocannabinoids have been isolated from Cannabis sativa L. (family Cannabaceae). Considering the association of CB1 receptor ligands with significant psychiatric adverse effects, research efforts have increasingly focused on the development of synthetic CB2-selective cannabinoids (e.g., 5-(tert-Butyl)-N-(1-hydroxy-2-methylpropan-2-yl)-1-(5-(trifluoromethyl)pyridin-2-yl)-1H-pyrazole-3-carboxamide, WIN55212-2) [46]. A selective CB2 receptor agonist has been shown to exert antifibrotic effects by attenuating leukocyte infiltration in the skin and limiting fibrosis-associated tissue injury [130].
Ciplukan herb (Physalis angulata L., family Solanaceae) ethanol extract contains higher phenol concentration compared to methanol or hexane extracts [131]. These phenolic compounds exert antioxidant and immunomodulatory effects, protecting the lymphocytes from ROS-induced damage, and influencing the autoimmune process [132,133], while also promoting leukocyte proliferation [50].
Comparison of the two target lists—the evidence-based molecular targets identified through the literature search and the predicted targets generated using the SwissTargetPrediction tool—revealed that they are almost entirely disjunct, with the MAPK p38α representing the sole exception. This near-complete separation is likely attributable to our selection criteria, as only predicted targets with a probability greater than 0.5 were included in the analysis. Additional predicted targets with lower probabilities (less than 0.5) exist, some of which may correspond to targets already identified in the evidence-based list (e.g., MMP-1, IL-6, and MAPK ERK1/2).
Given that the pathogenesis of SSc remains only partially elucidated and highly complex, the specialized literature cannot definitively establish the role of certain molecules in its pathogenic mechanisms. In this context, Table 8 includes those molecular targets for which subsequent searches in PubMed and ScienceDirect identified studies that demonstrate or suggest a possible connection to the pathogenesis of SSc. Thus, Table 8 brings together several categories of research, such as studies that highlight the existence of a genetic susceptibility to SSc (tyrosine-protein kinase receptor FLT3 [102], P-glycoprotein 1 [93]); studies demonstrating the presence of autoantibodies such as anti-carbonic anhydrase I [68] and II [68,69], anti-DNA topoisomerase II alpha [77], or anti-estrogen receptor alpha [79]; research showing differential expression levels of certain molecules in SSc (apoptosis regulator Bcl-2 [60] and matrix metalloproteinase-12 [87]); studies indicating a potential therapeutic benefits of inhibitors targeting certain molecules (e.g., acetylcholinesterase [56,57 and aldolase reductase [59]); and studies reporting altered values of certain molecules (e.g., matrix metalloproteinase-12 [87] and matrix metalloproteinase-13 [88]).
The extent to which these targets contribute to the various branches of the pathogenic triad of SSc is more or less clear. For instance, aldose reductase, a phase I metabolizing enzyme, was recently suggested to be a potential major player in skin fibrosis [59]. An overactive aldose reductase, by catalyzing NADPH-dependent detoxification of various substrates, may lead to NADPH depletion, increased oxidative stress, which further triggers inflammatory signaling pathways and fibrosis [134]. Despite the fact that tissue activity of aldose reductase has not yet investigated in humans with SSc, its inhibition has proved a skin antifibrotic effect in vivo model. Similarly, MMP-12 and MMP-13 represent well-documented contributors to the fibrotic component of SSc pathogenesis. These enzymes, through their role in extracellular matrix remodeling, have been implicated in fibrotic tissue involvement [87,88]. Thus, it would be worthwhile to evaluate the anti-SSc therapeutic relevance of the phytochemicals that exert inhibitory activity on these targets.
With respect to the inflammatory branch of the pathogenic triad in SSc, Toll-like receptors TLR7 and TLR9 have been identified as key, but functionally opposing regulators of immune dysregulation [101]. In a murine model of SSc, deletion of TLR7 attenuated skin and lung fibrosis and was associated with reduced infiltration of pro-inflammatory and profibrotic immune cells and cytokines in the skin [101]. In contrast, deletion of TLR9 exacerbated skin and lung fibrosis and increased inflammatory cell infiltration, suggesting a protective role for TLR9 in this model [101]. These divergent effects are mediated by plasmacytoid dendritic cells and TLR-dependent induction of type I interferons, a central pathway in SSc pathogenesis [101]. Thus, phytochemicals displaying antagonist activity on TLR7 or/and/or agonist activity on TLR9 may be added to the list of anti-SSc candidate agents.
In SSc dermal fibroblasts, upregulation of NADPH oxidase 4 (NOX4) drives a sustained increase in ROS production, establishing an ROS-mediated positive feedback loop that promotes fibroblast activation, extracellular matrix synthesis, and DNA damage [90]. Oxidative stress driven by excessive ROS production has been implicated in endothelial damage and vascular remodeling, contributing significantly to the characteristic vasculopathy of SSc [135]. Thus, NOX4 appears to be involved in both the fibrotic and vascular damage pathways underlying SSc pathogenesis. Therefore, it would be reasonable to assume that the phytocompounds, which have an inhibitory interaction with NOX4, could exert some beneficial effects on both the vascular and fibrotic pathogenic branches of SSc.
5. Limitations of the Study
This review has several limitations that should be acknowledged. One limitation of this review is that its protocol was not prospectively registered in PROSPERO. Although a narrative synthesis with an emphasis on molecular mechanism was appropriate for addressing the research objectives, the absence of prior registration may reduce methodological transparency and increase the possibility of selective reporting. Efforts were made to mitigate this risk through a comprehensive search strategy and detailed reporting of the review methods.
The body of evidence synthesized in this review spans a broad and heterogeneous spectrum of experimental models and clinical contexts. Most available data derive from preclinical studies, including in vitro experiments and murine models of SSc, which primarily provide mechanistic insights into antifibrotic, anti-inflammatory, and antioxidant pathways relevant to SSc. In contrast, human studies mainly report clinical outcome improvements, such as changes in the MRSS, frequency and severity of Raynaud phenomenon, hand mobility, or respiratory function parameters, with limited mechanistic exploration. A major limitation of the available literature is therefore the predominance of preclinical data, with relatively few clinical studies. Accordingly, findings from preclinical and human studies were discussed separately to reflect differences in evidentiary depth; preclinical observations establish biological plausibility and mechanistic rationale, while the available human data represent an initial step toward clinical translation. Overall, the current evidence should be interpreted as hypothesis-generating rather than practice-changing, underscoring the need for well-designed clinical studies.
Although the experimental evidence summarized in this review highlights multiple molecular pathways through which plant-derived compounds may modulate fibrosis, inflammation, and endothelial dysfunction in SSc, several pharmacological and clinical aspects warrant careful consideration. The majority of the available data originates from in vitro and animal models, in which the concentrations used often exceed those achievable or tolerable in humans. Consequently, issues related to bioavailability, pharmacokinetics, and dosing feasibility represent major translational barriers. For example, curcumin, a polyphenol isolated from Curcuma longa L., has demonstrated antifibrotic effects in vitro, while its in vivo bioavailability remains low [35,136]. Consequently, oral doses of curcumin required to achieve therapeutically relevant serum concentrations are very high (above 8 g) and generally intolerable [137]. However, studies have shown that co-administration of curcumin with piperine (an inhibitor of hepatic glucuronidation involved in curcumin metabolism) can enhance curcumin bioavailability by up to twofold [138]. Furthermore, long-term safety data for most of the reviewed compounds remain scarce. While short-term experimental studies suggest favorable toxicity profiles, chronic administration, particularly in patients with multisystem involvement, requires cautious assessment.
Regarding SwissTargetPrediction results, the probability score does not illustrate the probability for a phytochemical of being bioactive, but only the probability to behave as a ligand for a protein [13]. SwissTargetPrediction also does not distinguish the functional nature of phytochemical—protein interaction (agonism or antagonism). While conservative, the exclusion of putative targets annotated “by homology” may inadvertently eliminate biologically meaningful interactions and thus constitutes an additional limitation. Furthermore, variability in target nomenclature may have led to the omission of certain molecular targets relevant to SSc pathogenesis during the third search.
6. Future Direction of Research
Evidence from both in vivo (SSc animal models) and in vitro (skin biopsies from SSc patients) studies indicates that nuclear factor-erythroid 2-related factor 2 (Nrf2) activation confers both antifibrotic and anti-inflammatory benefits [139]. Therefore, it would be a reasonable approach to further investigate whether certain phytochemicals that have already displayed in other disease models interfere with the pathogenic triad of SSc, and might also have potential therapeutic benefits in SSc.
Regarding the first branch of the triad (inflammation), sulforaphane, an isothiocyanate derived from Brassica oleracea var. italica Plenck (family Brassicaceae), exhibited potent antioxidant and anti-inflammatory activities. These effects are primarily mediated through activation of the Nrf2 pathway, leading to the upregulation of antioxidant gene transcription (NADPH quinone oxidoreductase-1 and glutathione S-transferase) and the suppression of proinflammatory gene (IL-6 and IL-1β) transcription. Additionally, sulforaphane modulated the NF-kB signaling pathway by preventing the phosphorylation of IkBα, thereby restricting NF-kB nuclear translocation and subsequently suppressing iNOS (inducible form of nitric oxide synthase) expression [137].
A wide spectrum of phytochemicals, whose role was not yet fully explored in conjunction with Nrf2-SSc relationship, have also shown antioxidant potential through activation of the Nrf2 signaling pathway (carnosic acid [140], withaferin A [140], luteolin [140], salidroside [141], naringenin [141], resveratrol [141], sesaminol [141], ellagic acid [141], ginsenoside Re [141], tanshinone I [141], curcumin [141], naringin [141], tetramethylpyrazine [141], withametelin [141], magnolol [141], piperine [141], myricetin [141], parthenolide [142], vitexin [143], aspalathin [143], morin [143], silibinin [143], daphnetin [143], epigallocatechin 3-gallate [143], and quercetin [143]). Among these, flavonoids (luteolin, naringenin, naringin, myricetin, quercetin, vitexin, aspalathin, and morin) and phenolic acids (ellagic acid, resveratrol, salidroside, daphnetin, and epigallocatechin-3-gallate) constitutes the largest subgroups, highlighting the central role of polyphenols in redox modulation that could partially be explained by their ability to activate Nrf2 and regulate the transcription of antioxidant and cytoprotective genes. This shared mechanism of action underscores the therapeutic potential of naturally occurring Nrf2 activators as modulators of the oxidative pathway implicated in the pathogenesis of SSc.
Regarding the second and third branches of the triad (vascular injury and fibrosis), TGF-β-dependent hyperactivation of signal transducer and activator of transcription 3 (STAT3) represents another key mechanism of fibroblast activation that promotes the progression of fibrosis in SSc [144]. Evidence from an in vivo murine model of SSc demonstrates that STAT3 inhibition suppresses TGF-β myofibroblast differentiation, reduces collagen production and profibrotic gene expression, and improves skin fibrosis [144]. Therefore, STAT3 represents a critical molecular checkpoint in fibrosis, and its pharmacological inhibition could serve as a viable treatment option for SSc. Several phytocompounds, which have been reported to inhibit STAT3, not yet studied in connection with SSc, might have certain benefits (curcumin [145], resveratrol [146], epigallocatechin-3-gallate [147], ginsenosides [148], quercetin [149], pterostilbene [150], piperine [151], embelin [152], andrographolide [153], baicalein [154], luteolin [155], delphinidin [156], piceatannol [157], parthenolide [158], magnolol [159], and indirubin [160]).
The efficacy of all these phytochemicals has to be evaluated in SSc animal models or in vitro experiments based on fibroblasts derived from SSc patients, in order to establish their potential therapeutic relevance in SSc.
Other future research directions might be to identify potential synergisms between these phytochemicals and standard conventional therapy and to analyze their putative utility as complementary therapy in SSc.
Their extensive structural optimization may also be potentially useful in designing new drugs with clinical therapeutic efficacy in SSc.
The safety and potential long-term effects of these phytochemicals or plant extracts remain to be established, as they have not been investigated in most cases.
7. Conclusions
According to our search, several phytocompounds (e.g., halofunginone and tanshinone IIA) and whole plant extracts (e.g., caper bush and ciplukan) proved beneficial influences on the pathogenesis of SSc, and in some cases (e.g., proanthocyanidin and bromelain) even some degree of clinical therapeutic efficacy.
In conclusion, given the current lack of effective therapeutic strategies for SSc and the growing inclination of patients toward natural treatment options [161], the use of phytocompounds or plant-derived extracts emerges as a potentially valuable complementary or alternative approach for this pathology.
Although both in vivo and in vitro findings show encouraging outcomes, indicating that phytocompounds may exert multi-target effects on molecular pathways driving fibrosis, inflammation, and oxidative stress involved in SSc pathogenesis, a substantial gap remains in the clinical evidence.
Robust clinical trials are required to evaluate the safety and therapeutic efficacy of the identified phytocompounds in patients with SSc, with careful consideration of personalized medicine strategies given the pronounced heterogeneity of the disease.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Denton C.P. Khanna D. Systemic sclerosis Lancet 20173901685169910.1016/S 0140-6736(17)30933-928413064 · doi ↗ · pubmed ↗
- 2Lemmers J.M.J. Velauthapillai A. van Herwaarden N. Vonk M.C. Change of the microvascularization in systemic sclerosis, a matter of air Best Pr. Res. Clin. Rheumatol.20213510168310.1016/j.berh.2021.10168333814313 · doi ↗ · pubmed ↗
- 3Good S.D. Lee J.Y. Johnson R.E. Volkmann E.R. A scoping review of the epidemiology of systemic sclerosis and its organ manifestations: 2018–2024 Curr. Opin. Rheumatol.20253710311210.1097/BOR.000000000000106339470126 PMC 11779589 · doi ↗ · pubmed ↗
- 4Jerjen R. Nikpour M. Krieg T. Denton C.P. Saracino A.M. Systemic sclerosis in adults. Part I: Clinical features and pathogenesis J. Am. Acad. Dermatol.20228793795410.1016/j.jaad.2021.10.06535131402 · doi ↗ · pubmed ↗
- 5Lepri G. Di Battista M. Codullo V. Bonomi F. Sulis A. Guiducci S. Della Rossa A. Systemic sclerosis: One year in review 2024 Clin. Exp. Rheumatol.2024421517152810.55563/clinexprheumatol/is 29he 39058484 · doi ↗ · pubmed ↗
- 6Del Galdo F. Lescoat A. Conaghan P.G. Bertoldo E. ČolićJ. Santiago T. Suliman Y.A. Matucci-Cerinic M. Gabrielli A. Distler O. EULAR recommendations for the treatment of systemic sclerosis: 2023 update Ann. Rheum. Dis.202584294010.1136/ard-2024-22643039874231 · doi ↗ · pubmed ↗
- 7Szűcs G. Szekanecz Z. Szamosi S. Can we define difficult-to-treat systemic sclerosis?Expert. Rev. Clin. Immunol.2024201065108110.1080/1744666 X.2024.235245038711393 · doi ↗ · pubmed ↗
- 8Blaise S. Garcin F. Seinturier C. Gilson M. Imbert B. Mercan A. Practical survey on the use of herbal medicine by patients with systemic sclerosis Vasc. Dis.20255010411010.1016/j.vasdi.2025.05.00240762126 · doi ↗ · pubmed ↗