Jasmonic Acid Signals Involved in Valsa Canker Resistance Caused by C2H2-Type Transcription Factor PbeSTOP2 in Pyrus betulifolia
Longgang Zuo, Zhihong Liu, E Sun, Yuan Lu, Minrui Cai, Hongqiang Yu, Junying Zhao, Cunwu Zuo

TL;DR
This paper identifies a transcription factor, PbeSTOP2, that helps pear plants resist Valsa canker by boosting jasmonic acid signaling.
Contribution
The study reveals a new C2H2-type transcription factor, PbeSTOP2, involved in Valsa canker resistance through jasmonic acid signaling.
Findings
PbeSTOP2 expression is induced by Valsa canker pathogens and their metabolites.
Overexpression of PbeSTOP2 increases resistance to Valsa canker in pear cells.
PbeSTOP2 activates jasmonic acid and immunity-related genes, boosting disease resistance.
Abstract
Valsa canker, a destructive necrotrophic disease caused by Valsa species (V. mali and V. pyri), threatens global apple and pear production. Resistance breeding has gained much attention because of its environmental friendliness and effectiveness, making it extremely critical to obtain key disease resistance genes. In this study, we identified that the expression of PbeSTOP2, a C2H2-type transcription factor in ‘Duli-G03’ (Pyrus betulifolia, rootstock of pear), was induced in response to signals from Vp and Vp metabolites (VpM). Transient expression in fruits and stable overexpression in suspension cells demonstrated that PbeSTOP2 enhances resistance to Vp. In overexpressing cells, genes associated with jasmonic acid (JA) and pattern-triggered immunity (PTI) signaling were upregulated, and endogenous JA and auxin (IAA) levels increased. These results revealed that JA signaling was…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
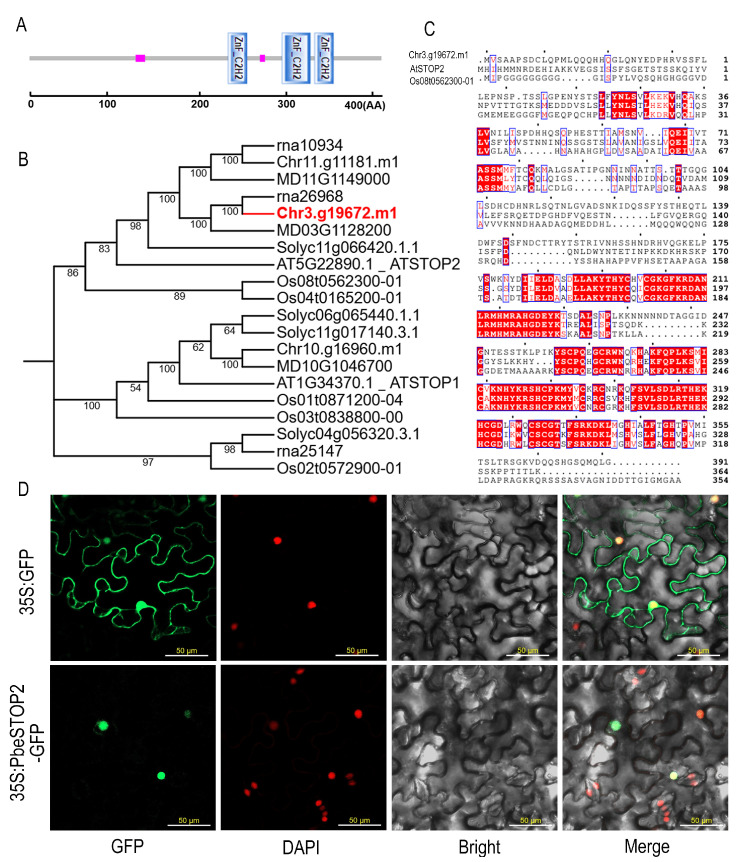 Figure 1
Figure 1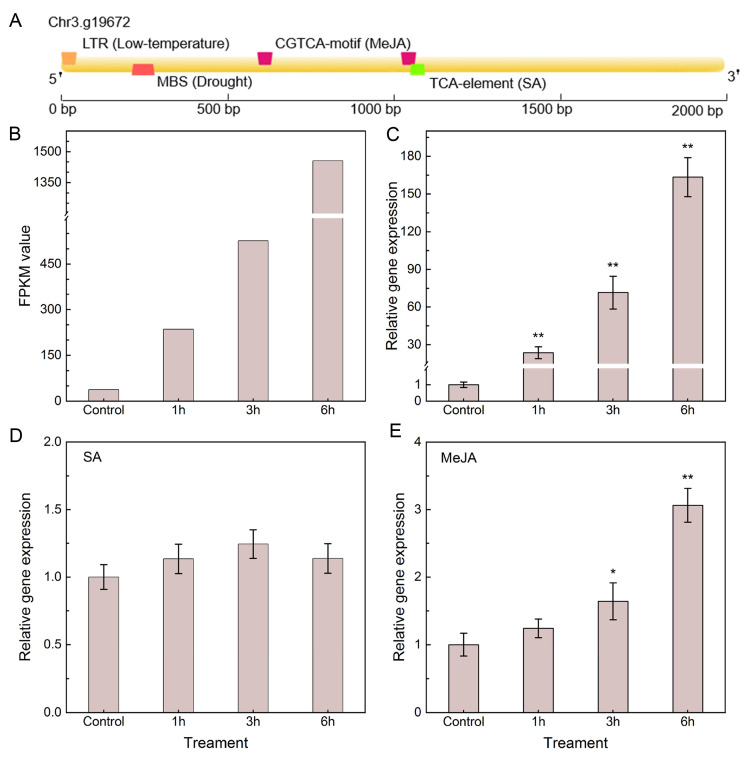 Figure 2
Figure 2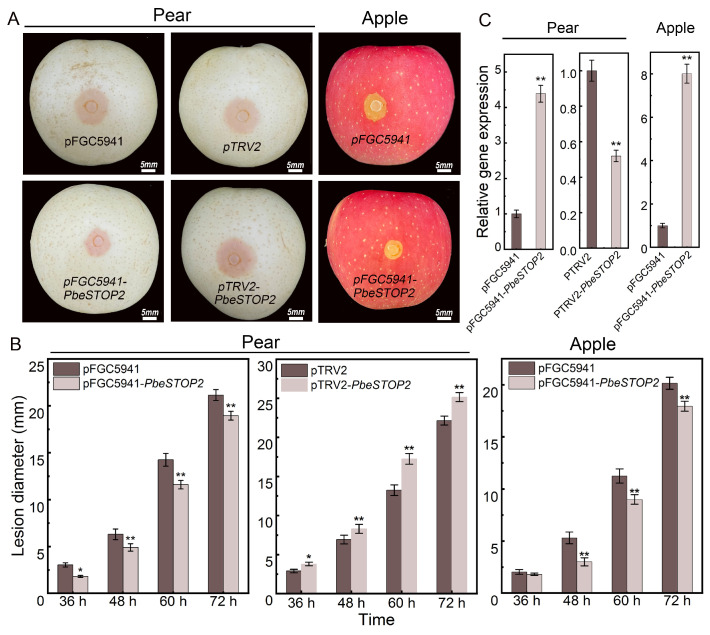 Figure 3
Figure 3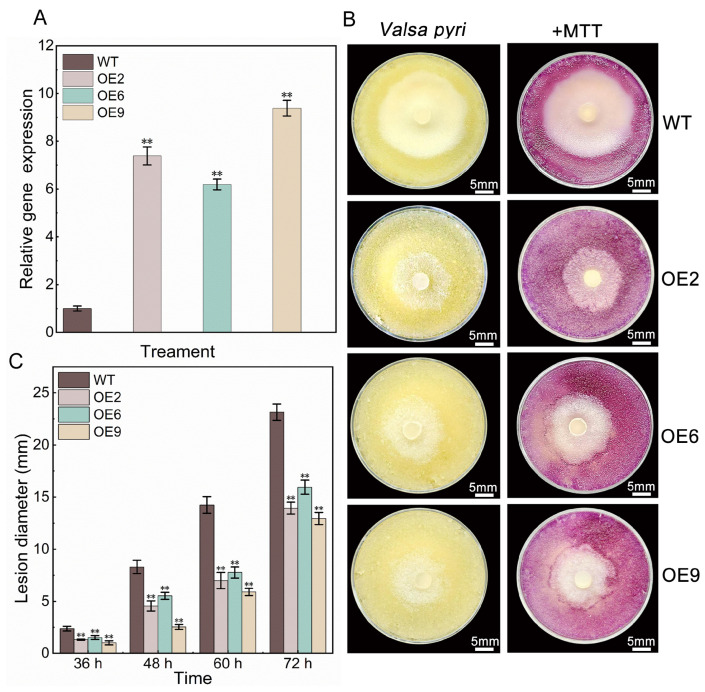 Figure 4
Figure 4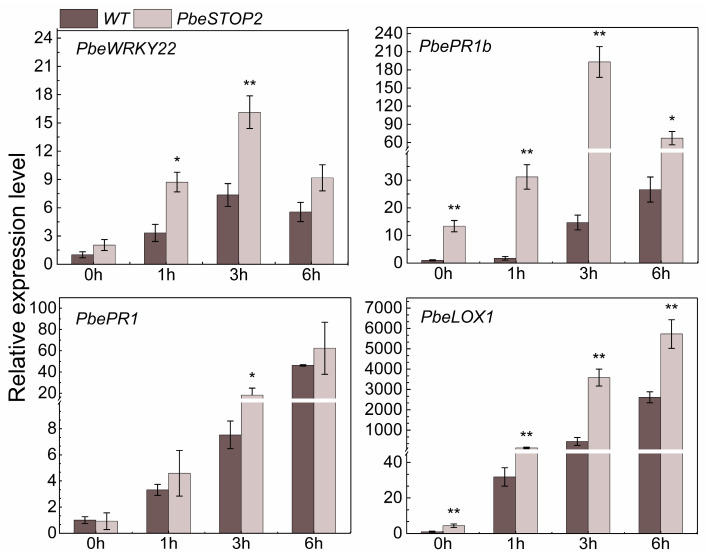 Figure 5
Figure 5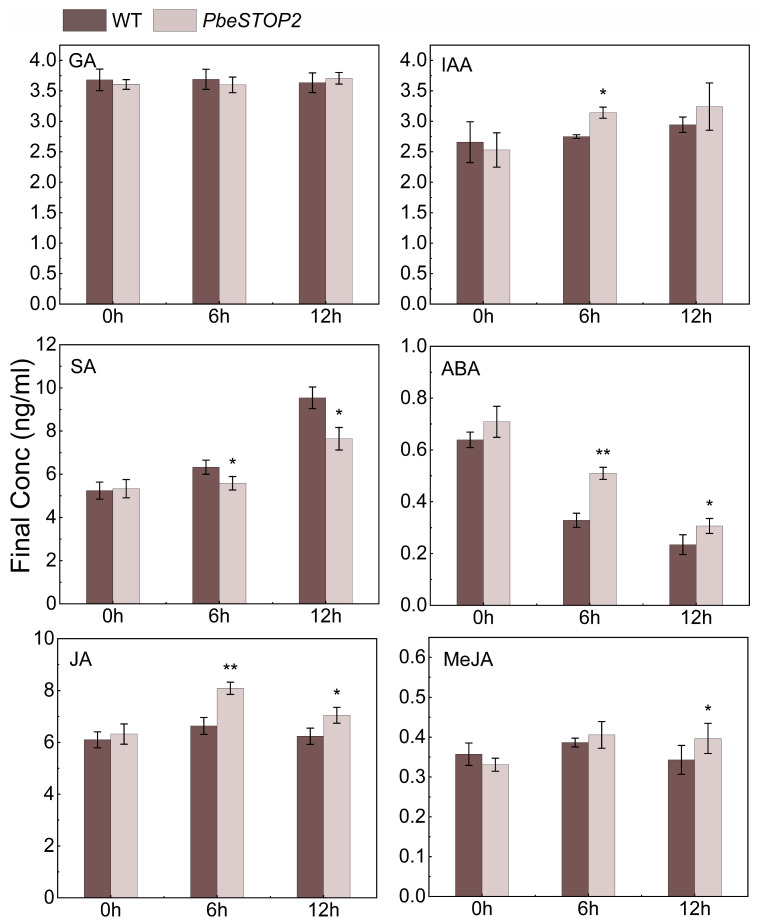 Figure 6
Figure 6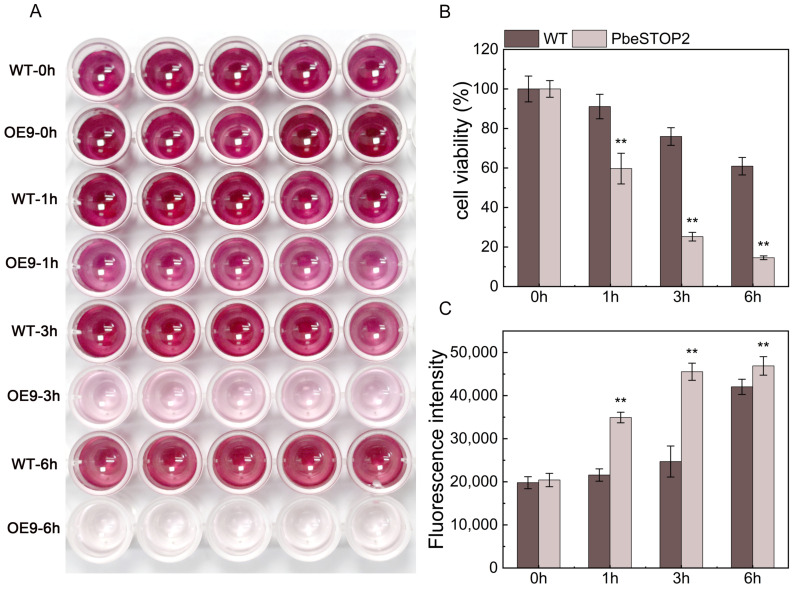 Figure 7
Figure 7- —Forestry and Grassland Science and Technology 430 Innovation Project of Gansu Province
- —The Science and Technology Major 431 Project of Gansu Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect-Plant Interactions and Control · Plant-Microbe Interactions and Immunity · Plant Virus Research Studies
1. Introduction
Valsa canker, a destructive disease caused by necrotrophic fungi (Valsa mali and V. pyri), poses a significant threat to apple and pear production [1]. The pathogen mainly attacks the bough and lateral branches via wounds, ultimately reducing orchard yield and fruit quality. Although chemical fungicides are widely used for control, their application is constrained by potential environmental and health risks. Resistance breeding, an environmentally friendly and effective practice, is commonly approved but has been restricted due to a prolonged breeding cycle [2]. Considering the multiple advantages of advanced molecular breeding, it is urgent to identify key genes regulating Valsa canker resistance from highly resistant germplasm.
During the long-term progress of struggling against pathogenic microbes, plants have evolved a natural two-tiered immune system, defined as horizontal resistance and vertical resistance [3]. First, pattern recognition receptors (PRRs) on the plant cell membrane could sense the signals from pathogens that produce harmful signaling molecules with special conserved motifs and trigger horizontal resistance, known as PRR-triggered immunity (PTI) [4]. Certain pathogens can inject virulence factors into plant cells to repress the activation of PTI for successful infection. After breaching the cell-surface defenses controlled by PRRs, these effector proteins can be specifically recognized by intracellular nucleotide-binding, leucine-rich repeat receptors (NLRs). This recognition in turn activates a stronger, vertical resistance response known as effector-triggered immunity (ETI) [4]. Subsequently, the local signals are transmitted throughout the plant and activate systematic acquired resistance (SAR). A series of signals, such as callus deposition, stomatal closure, hypersensitive response (HR), accumulation of reactive oxygen species (ROS), salicylic acid (SA) and jasmonic acid (JA), and transcription factors (TFs), take the crucial roles on initiation and signal transduction of immune responses [5,6]. Notably, the JA pathway is pivotal across multiple immune stages. Pathogen infection can disrupt the synthesis, metabolism, and signal transduction of JA within the host. The increase in JA levels activates downstream defense genes, thereby enhancing resistance to necrotic pathogens. For instance, exogenous application of methyl jasmonate (MeJA) can significantly enhance resistance against Sclerotinia sclerotiorum in both oilseed rape (Brassica napus) and soybean (Glycine max).
TFs are protein molecules that mediate intracellular signals and stress responses via regulating gene transcription [7,8]. Numerous TF families, such as zinc finger protein (ZFP), basic–leucine zipper (bZIP), myeloblastosis (MYB), N-acetylcysteine (NAC), high-mobility group (HMG), and APETALA2/ethylene-responsive element binding factor (AP2/ERF), have been shown to play vital and unique roles in plant specificity regulation [9]. Among these, ZFPs are the most widely distributed proteins in eukaryotic genomes, which can be classified into C2H2, C8, C6, C3HC4, C2HC, LIM, C4, C3H, and C4HC3 based on the number and position of cysteine (Cys) and histidine (His) residues [10,11]. C2H2 contains a zinc finger structure consisting of approximately 30 amino acids, with a conserved sequence of X2-Cys-X(2-4)-Cys-X12-His-X(3-5)-His (X represents any amino acid). The first plant-specific C2H2 member, ZPT2-1 (EPF1), was isolated in Petunia hybrida [12]. After that, a large number of C2H2-type ZFPs were successfully identified from various organisms, such as Arabidopsis thaliana [10], Oryza sative [13], Triticum [14], Solanum tuberosum [15], Solanum lycopersicum [16], and Nicotiana tabacum [17], with the member size recorded as 176, 189, 122, 79, 118 and 211, respectively. In-depth investigations revealed the crucial roles of numerous C2H2-type ZFPs in plant immunity. For instance, overexpression of the pepper C2H2-type ZFP gene CAZFP1 in transgenic Arabidopsis plants can enhance resistance to infection by Pseudomonas syringae [18]. In tomatoes, SlZF3 regulates plant height by directly inhibiting genes in the gibberellin biosynthesis pathway [19]. In addition, the C2H2-type ZFP transcription factor MdZAT10 in apples can significantly accelerate leaf senescence by promoting the expression of aging-related genes [20].
Duli is a rootstock belonging to Pyrus betulifolia that is widely planted in northern China as a rootstock due to its superior characteristics, such as wide adaptability and good graft compatibility. It has been proven to have high resistance against the attack of various pathogenic microbes, including the pathogen of Valsa canker [21]. ‘Duli-G03’ is a solitary plant identified by our research team during field investigations, which exhibits exceptional resistance to Valsa canker. Multiple previous investigations have been focused on the resistance mechanism of Duli against V. pyri infection. For instance, the PbeKCS1/10-1, PbeMPK4-1, MdCN19, and MdCN11 genes are determined as crucial signals that are involved in the responses of ‘Duli-G03’ (P. pyrifolia) against V. pyri [22,23]. In addition, based on the gene family analysis and functional validation, receptor-like proteins PbeRP23 and PbeRP27 are screened, which are key regulators for the Valsa canker resistance of ‘Duli-G03’ [24]. Unfortunately, whether C2H2 members also take vital roles in Valsa canker resistance has not been elucidated.
In a preliminary analysis, the expression of PbeSTOP2 was significantly activated when ‘Duli-G03’ suspension cells were challenged with signals of Valsa canker. In this study, by means of transient expression and stable transformation, we determined that PbeSTOP2 positively conferred Valsa canker resistance of ‘Duli-G03’ cells and apple and pear fruits. Moreover, the mechanistic investigations revealed that JA signals are involved in the resistance launching of immune response derived from PbeSTOP2.
2. Materials and Methods
2.1. Plant Materials and Pathogen Isolation
Calli was induced from leaves of pear ‘Duli-G03’ (Pyrus betulifolia) tissue-cultured plantlets. Vigorous callus was then subcultured in liquid MS medium and shaken at 130 rpm in darkness at 25 °C. After a week, cells were collected by filtration through a 40-mesh sieve. These suspension cells were maintained in MS medium under controlled conditions (25 °C, 130 rpm). The pathogenic fungus Valsa pyri strain Vp-001 was cultivated on potato dextrose agar (PDA) in complete darkness at 25 °C [25,26]. For Valsa pyri metabolites (VpM) extraction, static cultures of Vp-001 were grown in potato dextrose broth (PDB) at 25 °C without light. After 72 h of incubation, the culture supernatant was collected via centrifugation (8000× g, 10 min) and designated as undiluted VpM (100% concentration).
2.2. Bioinformatic Analysis of PbeSTOP2
The coding sequence (CDS) and corresponding protein sequence of PbeSTOP2 (Chr3.g19672.m1) were acquired from the publicly available Genome Database for Rosaceae (GDR: https://www.rosaceae.org/species/malus/all, accessed on 8 May 2025). Domain architecture prediction for the protein encoded by PbeSTOP2 was carried out with the web-based program SMART (http://smart.emblheidelberg.de/, accessed on 8 May 2025), using a significance threshold of E-value ≤ 1 × 10^−5^. Physicochemical characteristics and subcellular localization were evaluated, respectively, through ExPASy ProtParam (https://web.expasy.org/protparam/, accessed on 9 May 2025) and WoLF PSORT (https://www.genscript.com/wolf-psort.html, accessed on 9 May 2025). Candidate gene sequences from Arabidopsis thaliana, Oryza sativa, and Solanum lycopersicum were sourced from the Arabidopsis Information Resource (TAIR, https://www.arabidopsis.org/, accessed on 10 May 2025) and the National Center for Biotechnology Information (NCBI, https://www.ncbi.nlm.nih.gov/, accessed on 10 May 2025). Multiple sequence alignment was executed in Clustalx 2.0, and resultant alignments were subsequently analyzed with MEGA 8.0 (http://www.megasoftware.net, accessed on 11 May 2025) for evolutionary inference. A phylogenetic tree was generated via the neighbor-joining approach under the following settings: P-distance substitution model, pairwise deletion treatment, and 1000 bootstrap replications. The 2000 bp promoter region upstream of the PbeSTOP2 initiation site was extracted from the same database, and cis-regulatory elements within this sequence were predicted using Plant CARE (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 11 May 2025).
2.3. Subcellular Localization of PbeSTOP2
The full-length coding region of PbeSTOP2 was ligated into the pCAMBIA1300-GFP vector. Following sequence verification, the fusion construct 35S:PbeSTOP2-GFP was introduced into Agrobacterium tumefaciens strain GV3101 [27]. 35S:PbeSTOP2-GFP was activated, resuspended, and pressure-infiltrated into the leaves of Nicotiana benthamiana, with empty vector 35S: GFP as the control. After 48 h dark incubation, infiltrated leaf tissues were co-stained with 4′,6-diamidino-2-phenylindole (DAPI, 1 μg/mL) for nuclear visualization. Subcellular localization analysis was conducted on a laser scanning confocal microscope (FV1200, Olympus Corporation, Tokyo, Japan.).
2.4. Exogenous Hormone Treatment
For exogenous hormonal treatments, methyl jasmonate (MeJA) or SA at a concentration of 50 μM was added to ‘Duli-G03’ cell suspensions and incubated at 25 °C in the darkness. SA was purchased from Shanghai Yuan Ye Biotechnology Co. (Shanghai, China), and MeJA was purchased from Sigma-Aldrich ((St. Louis, USA). Following collection of cells at designated intervals (0, 1, 3, 6 h post-treatment), total RNA was isolated for subsequent qRT-PCR analysis performed in accordance with Zhao et al. [21].
2.5. Transient Expression of PbeSTOP2 in Fruits
The full-length coding sequence of PbeSTOP2 was cloned into the pFGC5941 vector via homologous recombination, and the recombinant plasmid PFGC5941-PbeSTOP2 was obtained. The genome-specific fragment PbeSTOP2 (318 bp) was introduced into the pTRV2 vector. All constructs were transformed into Agrobacterium strain GV3101. Positive colonies of GV3101 harboring PFGC5941-PbeSTOP2, pTRV2-PbeSTOP2 and empty vectors were cultured in 15 mL LB liquid medium containing 50 mg/L kanamycin and 50 mg/L gentamicin, shaking (180 rpm) at 28 °C to OD_600_ value of 1.0, followed by centrifugation at 5000 rpm for 8 min. For functional validation, transient overexpression and Virus-Induced Gene Silencing (VIGS) assays were conducted on ‘Huangguan’ pear and ‘Yanfu 6’ apple fruits via Agrobacterium-mediated infiltration [22]. Agrobacterium containing an empty vector was injected as the control. Three days post-infiltration, Vp was inoculated at the infiltration sites. Samples were incubated in darkness at 25 °C, and lesion diameters were measured every 12 h using digital vernier calipers.
2.6. Stable Overexpression and Functional Verification of PbeSTOP2
Suspension cells derived from ‘Duli-G03’ were pre-cultured in a constant-temperature oscillator at 25 °C for a duration of 3 days. Agrobacterium tumefaciens strain GV3101 carrying the 35 S: PbeSTOP2 recombinant plasmid was cultured until the bacterial culture reached an OD_600_ of 1.0, followed by resuspension in 2-(4-morpholino)ethanesulfonic acid (MES) medium. Afterwards, 2 mL of the prepared Agrobacterium suspension was mixed with 20 mL of the pre-cultured ‘Duli-G03’ suspension cells, and the mixture was incubated for 20 min. After removing residual Agrobacterium by centrifugation, treated cells were incubated for 3 days. Residual bacteria were eliminated by antibiotic treatment, and cells were plated on MS solid medium for 20-day dark incubation at 25 °C [22]. Overexpression lines were selected based on PCR and Quantitative real-time PCR (qRT-PCR) validation. The transgenic and wild-type (WT) cells were maintained separately in MS liquid medium (25 °C, 130 rpm), then 2 mL were transferred to MS solid medium. After 3 days of incubation in darkness at 25 °C, each plate was inoculated with a 5 mm plug of Vp strain. Lesion diameters were measured every 12 h using digital vernier calipers [22]. PbeSTOP2-overexpressing cells were infected with VpM at a concentration of 20%. For the control, the WT cells were treated under the same conditions described above. All samples were harvested at 0, 1, 3, and 6 h post-treatment, and cell viability was assessed via the methyl thiazolyl tetrazolium (MTT) assay [24]. For ROS detection, suspension cells of the PbeSTOP2-overexpressing cells and the WT were treated with 20% VpM solution. Samples were collected at 0, 1, 3, and 6 h and incubated with the ROS-specific fluorescent dye H2DCFDA (10 µmol L^−1^) for 10 min at 37 °C in the dark, followed by quantification in labeled cells using a Spark microplate reader (Tecan Group Ltd., Männedorf, Switzerland). The excitation and emission wavelengths were 488 nm and 525 nm, respectively.
2.7. Gene Expression Assays
The expressional patterns were detected by extracting data from released RNA-seq data and using a Quantitative real-time PCR (qRT-PCR) assay. For the RNA-seq data, ‘Duli-G03’ suspension cells treated with VpM for 1, 3, and 6 h were utilized [21]. Primers for both target were designed with the online tool Primer 3.0 plus (https://www.primer3plus.com/index.html, accessed on 15 May 2025) (Table 1). The reference gene TUB (β-Tubulin, Chr3.g20215) in Pyrus spp. was identified based on the method by Pessina et al. [28]. Total RNA was extracted from suspension cells of ‘Duli-G03’ using the Tiandz RNA extraction kit (Catalog No. 71203-50, Tiangen Biotech, Beijing, China) following the manufacturer’s protocol. The extracted RNA was treated with DNase I (1 U/μg RNA) at 37 °C for 30 min to eliminate genomic DNA contamination. First-strand cDNA was then synthesized from 1 μg of the DNase-treated RNA using PrimeScript™ Reverse Transcriptase (Takara Bio Inc., Kusatsu, Japan) and oligo(dT)18 primers in a 20-μL reaction volume. The reverse transcription was performed at 42 °C for 30 min, with subsequent enzyme inactivation at 85 °C for 5 s. qPCR was performed with SYBR^®^ Green Premix (Bio-Rad) on a CFX96 Touch™ system. Program: 95 °C for 30 s; 40 cycles of [95 °C for 5 s, 60 °C for 30 s, 72 °C for 30 s]; melting curve from 65 °C to 95 °C at 0.5 °C/5 s. Efficiency was determined via a standard curve from 10-fold cDNA dilutions, calculated as E = (10^−1/slope^ − 1) × 100%. All primer efficiencies were 90–110% (R^2^ > 0.99). Expression was normalized to TUB and calculated using the 2^−ΔΔCT^ − method [29]. Statistical analysis was performed in Microsoft Excel 2010. All experiments included at least three biological and technical replicates.
2.8. Endogenous Hormone Assays
Liquid chromatography-tandem mass spectrometry (LC-MS) for the determination of endogenous cellular hormones [30]. First, cell samples (1 g) were collected, ground in liquid nitrogen, and placed in 15 mL centrifuge tubes. Then, 10 mL of pre-cooled methanol-formic acid solution (99:1) was added, sonicated at 22 °C for 2 min, and stored at 4 °C for 12 h. Next, the extract was centrifuged at 10,000 r/min for 10 min, 1.0 mL of supernatant was aspirated, and H_2_O was added to the 10 mL of solution. Then, the ODS C18 solid phase extraction column was used for the adsorption of the upper samples, and the column was washed with 6 mL of 10% methanol solution in two batches. Finally, it was eluted with methanol-formic acid solution and fixed to 1.0 mL, and the sample was collected after filtration with a 0.22 μm microporous filter membrane. The instrument LC-MS coupled instrument (Agilent 1290-6460, Agilent Technologies, Santa Clara, CA, USA) was used for sample detection. LC conditions: Detection wavelength was set to 290 nm, column temperature maintained at 35 °C, and flow rate fixed at 0.3 mL/min. Mobile phase consisted of B (methanol): A (0.1% formic acid in water) with gradient elution programmed as follows: 0–1 min from 10% to 40% B, 1–2.5 min from 40% to 45% B, 2.5–5 min from 45% to 80% B, 5–5.1 min from 80% to 10% B, and 5.1–7.1 min held at 10% B; injection volume was 5 μL. MS conditions: Electrospray ionization source (ESI) was operated in negative ion mode with multiple reaction monitoring (MRM). Parameters included ion source temperature at 350 °C, drying gas flow rate at 11 L/min, nebulizer pressure at 35 psi, capillary voltage at 4 kV, and collision gas of high-purity nitrogen.
2.9. Statistical Analysis
The basic data were statistically analyzed in Microsoft Excel 2010 and visualized with Origin 2022. Differences between means were determined using Student’s t-test (** p* < 0.05; *** p* < 0.01).
3. Results
3.1. PbeSTOP2 Encoded a Typical C2H2-Type Zinc Finger Protein
Bioinformatics analysis of Chr3.g19672 was performed to obtain its related information. Domain prediction results demonstrated that the target gene encodes a protein that possesses three typical C2H2-type zinc fingers at 232 to 254, 295 to 328, and 333 to 355, respectively (Figure 1A). BLAST(NCBI BLAST+ 2.14.0, accessed on 11 May 2025) analysis indicated that the target protein shares high similarity with Sensitive To Proton Rhizo Toxicity (AtSTOP2) in Arabidopsis, leading to its designation as PbeSTOP2. Phylogenetic reconstruction revealed that PbeSTOP2 exhibits closer evolutionary relationships with homologs from Rosaceae species compared to those from other plant families (Figure 1B). The results of the microcolinearity analyses indicate that PbeSTOP2 is a direct ortholog of the Arabidopsis thaliana AtSTOP2 and rice Os08t0562300-01 (Figure S1). Based on sequence alignment, PbeSTOP2 shared 41.35% and 41.61% sequence identity with AtSTOP2 and Os08t0562300-01, respectively (Figure 1C). Additionally, transient expression in Nicotiana benthamiana leaves confirmed the nuclear localization of PbeSTOP2 (Figure 1D). Connectively, the above results indicated that PbeSTOP2 was a typical C2H2-type zinc finger protein and may possess unique functions in the nucleus.
3.2. PbeSTOP2 Responded to Vp and Related Defense Signals
We predicted the cis-element on the promoter region of PbeSTOP2 to investigate its potential function. Multiple stress- and signaling-related cis-elements were identified, including those responsive to MeJA, SA, and various abiotic stresses (Figure 2A). We extracted RNA-seq data of PbeSTOP2 after treating ‘Duli-G03’ suspension cells with VpM. Compared with the control, the expression of PbeSTOP2 was significantly up-regulated in ‘Duli-G03’ suspension cells during VpM infiltration (Figure 2B). This expression pattern was further validated by qRT-PCR analyses (Figure 2C). We further monitored the response pattern of PbeSTOP2 to exogenous SA and MeJA signaling in ‘Duli-G03’. The results showed that the expression of PbeSTOP2 was not changed with exogenous SA exposure, but was robustly activated under MeJA treatment (Figure 2D,E). In brief, the above results indicate that PbeSTOP2 positively responded to VpM and exogenous MeJA exposure.
3.3. PbeSTOP2 Positively Regulates Valsa Canker Resistance of Apple and Pear Fruits
To investigate the role of the PbeSTOP2 gene in Valsa canker resistance, we performed overexpression and virus-induced gene silencing (VIGS) of the target gene via A. tumefaciens-mediated transformation in fruits of ‘Huangguan’ pear and ‘Yanfu 6’ apple, respectively. Examination of lesions showed that transiently transformed fruits overexpressing PbeSTOP2 had significantly decreased lesion diameters compared to controls. In addition, VIGS experiments showed that lesion diameters were significantly increased in PbeSTOP2 silenced fruit compared to those in the control (Figure 3A,B). Overexpression and silencing of the genes at the infection site was confirmed by qRT-PCR (Figure 3C). These findings indicate that PbeSTOP2 acts as a positive regulator of resistance against Vp.
3.4. PbeSTOP2 Overexpression Enhanced Vp Resistance of ‘Duli-G03’ Suspension Cells
To further validate the regulatory role of PbeSTOP2 on Valsa canker, the target gene was overexpressed in suspension cells of ‘Duli-G03’. Three independent overexpression lines (PbeSTOP2-OE2/OE6/OE9) exhibited transcript levels that were 8-, 6-, and 10-fold higher, respectively, than those in WT cells (Figure 4A). Pathogen inhibition assays revealed markedly suppressed mycelial growth of Vp on all overexpression lines compared to the WT cells (Figure 4B). These observations were further corroborated by MTT-based viability assays. Quantitative detections recorded the colony diameter from 13 to 16 mm on overexpressing cell lines, whereas the diameter was 23 mm on the WT cells (Figure 4C). These findings demonstrate that elevated expression of PbeSTOP2 significantly enhances resistance to Valsa canker in ‘Duli-G03’ cells.
3.5. PbeSTOP2 Initiated Immune Responses
To investigate the signaling pathway activated by PbeSTOP2, we analyzed the expression of four defense-related marker genes during ‘Duli-G03’ suspension cells response to VpM signals (Figure 5). Compared with the WT cells, the expression of the PTI-related gene PbeWRKY22 [31] was significantly up-regulated at 1 and 3 h. For the SA-related gene, PbePR1 [31], the higher expression level was only observed from the overexpressing cells at 3 h compared with that from the WT cells. Strikingly, compared with the WT cells, prompt ascending of the JA-related genes PbePR1b and PbeLOX1 [32] was observed from the overexpressing cells at all four time points. Therefore, the above data suggest that JA and other defense-related signals are involved in regulating the defense response of PbeSTOP2.
3.6. PbeSTOP2 Induced the Accumulation of Defense-Related Phytohormones
To elucidate the function of phytohormones in the Valsa canker resistance mediated by PbeSTOP2, we examined the content of multiple hormones in overexpression and WT cells (Figure 6). For the Gibberellin (GA) levels, there was no obvious differentiation between the overexpression and WT cells. In addition, the increase in auxin (IAA) levels was observed in the overexpressing cells only at 6 h compared with the control. The lower levels of SA were discovered from the overexpressing cells than from the WT cells at 6 h and 12 h, whereas higher levels of abscisic acid (ABA) were detected. Notably, compared to WT cells, only JA levels were significantly elevated in overexpression cells at 6 h, whereas both JA and MeJA showed marked increases by 12 h. Thus, these data revealed the crucial roles of JA signals on PbeSTOP2-regulated Valsa canker resistance.
3.7. PbeSTOP2 Enhanced VpM Sensitivity and ROS Bursting of ‘Duli-G03’ Suspension Cells
Cell viability and ROS accumulation were also investigated to clarify the potential mechanisms of PbeSTOP2-conferred Valsa canker resistance. The cell viability of PbeSTOP2 was significantly decreased compared with the WT cells at 1, 3, and 6 h of VpM exposure (Figure 7A,B). In contrast, ROS quantitative analysis showed that the relative fluorescence ratio in PbeSTOP2-OE9 was significantly higher than in the WT cells at all time points (Figure 7C). Thus, these data also imply that cell viability and ROS accumulation are involved in PbeSTOP2-mediated Valsa canker resistance.
4. Discussion
As a result of evolution, plants have developed a series of stress-responsive protective mechanisms by regulating intracellular gene ex patterns pression, and an increasing number of TFs have been found to play key roles in responding to stress. In this study, we found that PbeSTOP2 was a typical C2H2-type TF, and the transcription of PbeSTOP2 was robustly induced by VpM and MeJA. Importantly, overexpression of PbeSTOP2 significantly enhanced the Valsa canker resistance of apple and pear fruits and ‘Duli-G03’ cells, and activation of the JA-related pathway was critical to the enhanced resistance.
In organisms, TFs directly or indirectly recognize and bind to the promoter region of target genes, activate the RNA polymerase activity of the dynamic transcription complex, and thus accomplish the regulation of gene expression [12,33]. Upon recognition of stress-associated cis-elements, transcription factors can respond to stress on C2H2-type gene regulation [34]. For instance, LOS2 (a bi-functional enolase) represses STZ/ZAT10 expression by binding to its promoter [35]. The cold stress C-repeat binding factor (CBF3) positively regulates gene ZAT10 expression through a DRE cis-element [36]. The distribution of multiple stress-related cis-elements in the PbeSTOP2 promoter region suggests that the regulation and expression of the target gene may be synergistically regulated by different hormones. In our study, we confirmed that the expression level of PbeSTOP2 was strongly induced by exogenous JA. These results implied that the resistance of PbeSTOP2 to Valsa canker is likely to depend on the activation of JA-related pathways.
Plants sensing pathogen signals stimulate their own PTI response, immediately followed by the initiation of nonspecific basal defense responses, manifested by the activation of the mitogen-activated protein kinase (MAPK) cascade, rapid deposition of callose-rich cell wall enforcements, bursts of ROS, etc. [37]. ROS, as part of the cellular signaling network, can be integrated with other signaling pathways such as ABA, SA, and JA to defend against pathogen infestation. It has been shown that ROS plays a major role in both PTI and ETI responses [38,39]. ROS production by the NADPH oxidase RBOHD is a critical early signal linking PRR and NLR-mediated immunity, and the receptor-like cytoplasmic kinase BIK1 is integral to this process [40]. In Arabidopsis, D36E (avrRpt2)-treated co-receptor mutants showed differential expression of genes regulated by BIK1, including many PTI immune pathway marker genes, such as WRKY22/29 and FRK1 [39]. However, when the concentration of ROS is too high, it can also lead to programmed cell death (PCD) and act as a layer 1 defense during a pathogen attack [41]. Signaling molecules that promote death through autophagy during pathogen-induced hypersensitivity response programmed cell death (HR-PCD) restrict cell death to the infected tissue site, effectively controlling the spread of HR to healthy tissues [42,43]. Following the silencing of Beclin1, a homolog of the yeast autophagy-related gene ATG6/Beclin1, HR-PCD triggered by tobacco mosaic virus (TMV) infection in tobacco was uncontrolled and spread to uninfected tissues [44]. In this study, a significant ROS burst was observed in PbeSTOP2-overexpressing cells after VpM infection compared with the WT cells, and the PTI-related gene PbeWRKY22 was up-regulated. Nevertheless, PbeSTOP2-overexpressing cells infected by VpM exhibited lower cell viability than the WT cells. We suggest that ROS production during PbeSTOP2-mediated immunization may act as a signaling molecule to initiate downstream molecular cascade reactions, while the burst of ROS caused a decrease in PbeSTOP2-overexpressing cell viability.
The response of plants to pathogen attacks is the result of a series of changes at the cellular level, which are mediated by hormone signals, such as SA, JA, ET, IAA, ABA, etc. [45]. Studies have shown that there is an overlap between different hormone signaling pathways, resulting in tightly regulated plant resistance mediated by these signaling pathways [46]. It is generally accepted that SA is involved in defense against biotrophic pathogens, whereas JA and ET contribute to defense against necrotrophic pathogens [47]. The effect of SA on the JA pathway can be antagonistic, synergistic, or neutral [48,49]. Previous studies have identified several members that play a role in JA-SA antagonism, including MYC2, PDF1.2, NPR1, WRKY62, GRX480, JAZs, etc. [50,51]. A recent gene-wide transcriptome study revealed a wide range of synergistic effects of JA-SA in Arabidopsis [52]. In this study, the expression of JA signaling-related genes PbeLOX1 and PbePR1b was strongly induced in the PbeSTOP2-overexpressing cell after Vp infection compared with the WT cells. Furthermore, the endogenous JA concentration was significantly higher in the transgenic cells than in the WT cells after pathogen inoculation. Differently, although the expression of the SA-related gene PbePR1 was up-regulated in PbeSTOP2-overexpressing cells, the endogenous SA content was lower than that of the WT cells. Therefore, we suggest that the disease resistance conferred by PbeSTOP2 depends on the JA signaling pathway but not the SA signaling pathway.
Furthermore, JA signaling engages in complex interactions with other phytohormones to coordinate plant immunity. Among them, IAA positively regulates necrotrophic pathogens; for instance, in Arabidopsis, IAA-deficient mutants are more sensitive to necrotrophic pathogens than WT [53]. Impairment of the auxin-activated SCF (Skp1–Cullin–F-box) ubiquitination pathway in Arabidopsis mutants axr1, axr2, and axr6 confers increased susceptibility to Plectosphaerella cucumerina and Botrytis cinerea [54]. Following Vm infection, genes associated with IAA and ABA signaling pathways exhibit downregulated expression in susceptible apples [55]. However, the role of ABA in plant disease resistance is complex and appears to vary in different plant-pathogen interactions [56]. In addition, ABA has multifaceted roles in different stages of plant defense. In the initial stage, ABA resists fungal invasion by mediating stomatal closure [57]. In the second stage, it promotes callus deposition in response to fungal infection [58]. Finally, ABA interacts with SA-, JA- and ET-dependent defense pathways. For example, ABA regulates JA-dependent resistance by inhibiting the JA response that is synergistically regulated by ET [59]. but promotes the JA branch that is antagonistic to ET [60]. In the present study, upregulation of IAA hormone concentration in PbeSTOP2-overexpressing suspension cells after infestation with Vp implies that IAA is presumably involved in PbeSTOP2-mediated disease resistance. Unusually, the endogenous ABA concentration was significantly higher in PbeSTOP2-overexpressing suspension cells than in WT cells after Vp infection. Therefore, the mechanism of ABA in the regulation of Valsa canker resistance by PbeSTOP2 requires further validation.
5. Conclusions
In summary, PbeSTOP2, a member of the C2H2-type zinc finger protein family, functions as a positive regulator of resistance to Valsa canker in both ‘Duli-G03’ cells and fruits of pear and apple. The signal pathways associated with ROS, cell death, and JA are probably involved in the immune response regulated by PbeSTOP2. Moreover, the results of expressional assay and hormone determination suggest that JA signals take crucial roles in the launching of Valsa canker resistance caused by PbeSTOP2. Therefore, the results of this study provide new insight into resistance breeding and further comprehensive prevention and control for the occurrence of Valsa canker.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Meng X. Qi X. Han Z. Guo Y. Wang Y. Hu T. Wang L. Cao K. Wang S. Latent Infection of Valsa mali in the Seeds, Seedlings and Twigs of Crabapple and Apple Trees is a Potential Inoculum Source of Valsa Canker Sci. Rep.20199773810.1038/s 41598-019-44228-w 31123304 PMC 6533284 · doi ↗ · pubmed ↗
- 2Liu X. Ao K. Yao J. Zhang Y. Li X. Engineering plant disease resistance against biotrophic pathogens Curr. Opin. Plant Biol.20216010198710.1016/j.pbi.2020.10198733434797 · doi ↗ · pubmed ↗
- 3Boller T. Felix G. A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors Annu. Rev. Plant Biol.20096037940610.1146/annurev.arplant.57.032905.10534619400727 · doi ↗ · pubmed ↗
- 4Jones J. Dangl J. The plant immune system Nature 200644432332910.1038/nature 0528617108957 · doi ↗ · pubmed ↗
- 5Aksimov I. Shein M. Burkhanova G. RNA Interference in Plant Protection from Fungal and Oomycete Infection Appl. Biochem. Microbiol.202258(Suppl. 1)S 16S 3110.1134/S 0003683822100106 · doi ↗
- 6Zhang S. Li C. Si J. Han Z. Chen D. Action Mechanisms of Effectors in Plant-Pathogen Interaction Int. J. Mol. Sci.202223675810.3390/ijms 2312675835743201 PMC 9224169 · doi ↗ · pubmed ↗
- 7Rushton P. Somssich I. Transcriptional control of plant genes responsive to pathogens Curr. Opin. Plant Biol.1998131131510.1016/1369-5266(88)80052-910066598 · doi ↗ · pubmed ↗
- 8Mitsuda N. Ohme-Takagi M. Functional analysis of transcription factors in Arabidopsis Plant Cell Physiol.2009501232124810.1093/pcp/pcp 07519478073 PMC 2709548 · doi ↗ · pubmed ↗