Glutamine Modulates mVOC Biosynthesis in Streptomyces alboflavus Through a gluR-Dependent Signaling Pathway and Enhances Its Inhibitory Activity Against Aspergillus flavus
Wangqiang Li, Mingguan Yang, Zehua Dong, Tong Liu, Xiuyu Liu, Dan Liu, Chengfang Ding, Laifeng Lu, Wentao Ding, Zhenjing Li, Huanhuan Liu, Zhifang Wang, Qingbin Guo, Changlu Wang

TL;DR
Glutamine boosts the production of antifungal compounds in Streptomyces alboflavus, helping it fight Aspergillus flavus more effectively.
Contribution
The study reveals a gluR-dependent signaling pathway by which glutamine modulates mVOC biosynthesis in Streptomyces alboflavus.
Findings
40 mM glutamine increased inhibition of A. flavus conidial germination by 69.0% and mycelial biomass by 64.5%.
Transcriptome analysis identified 283 differentially expressed genes, including the upregulated two-component system regulator gluR.
CRISPR/Cas9 disruption of gluR reduced antifungal mVOCs like dimethyl trisulfide and o-anisidine.
Abstract
Aspergillus flavus and its aflatoxins pose serious threats to human and animal health, negatively affecting agricultural productivity and the global economy. Although chemical preservatives are widely used, their effectiveness remains limited by increased fungal resistance and environmental concerns, highlighting the need for sustainable alternatives. Microbial volatile organic compounds (mVOCs) represent a promising biocontrol strategy. Here, we investigate how glutamine regulates mVOC biosynthesis in Streptomyces alboflavus TD-1 and enhances its antifungal activity against A. flavus. Antifungal assays showed that supplementation with 40 mM glutamine significantly enhanced inhibitory activity, leading to 69.0% inhibition of conidial germination and 64.5% inhibition of mycelial biomass. Transcriptome profiling identified 283 differentially expressed genes, including the two-component…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
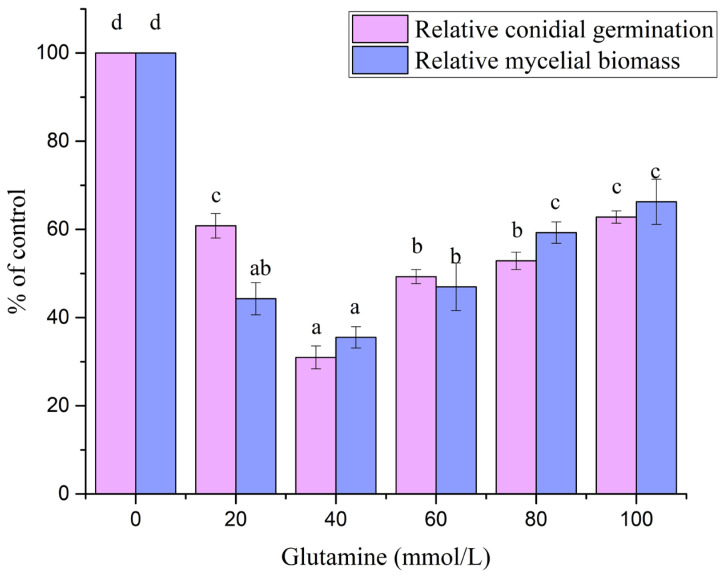 Figure 1
Figure 1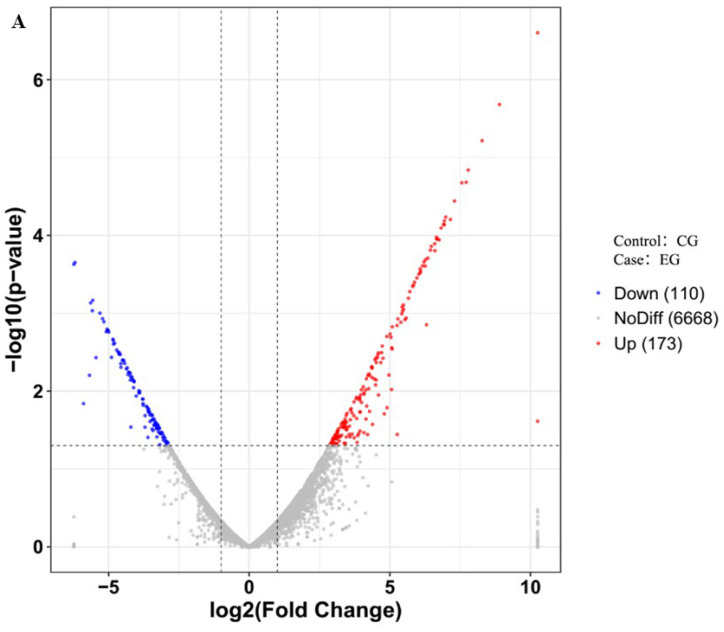 Figure 2
Figure 2 Figure 3
Figure 3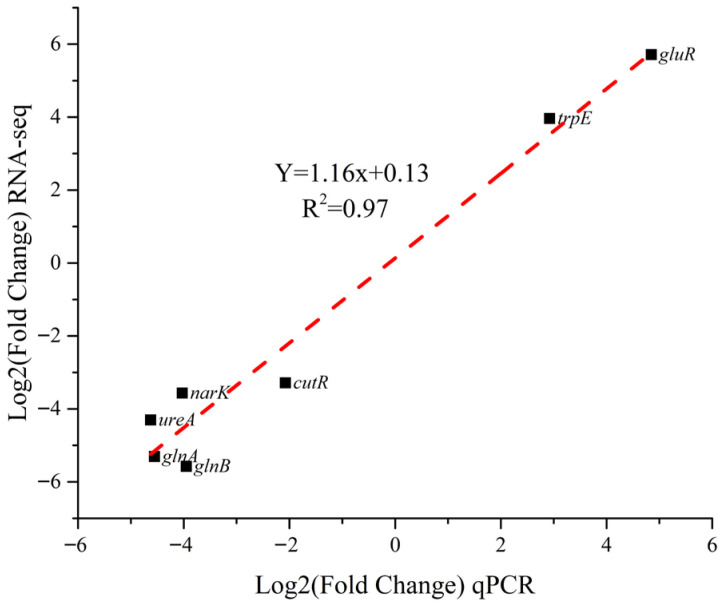 Figure 4
Figure 4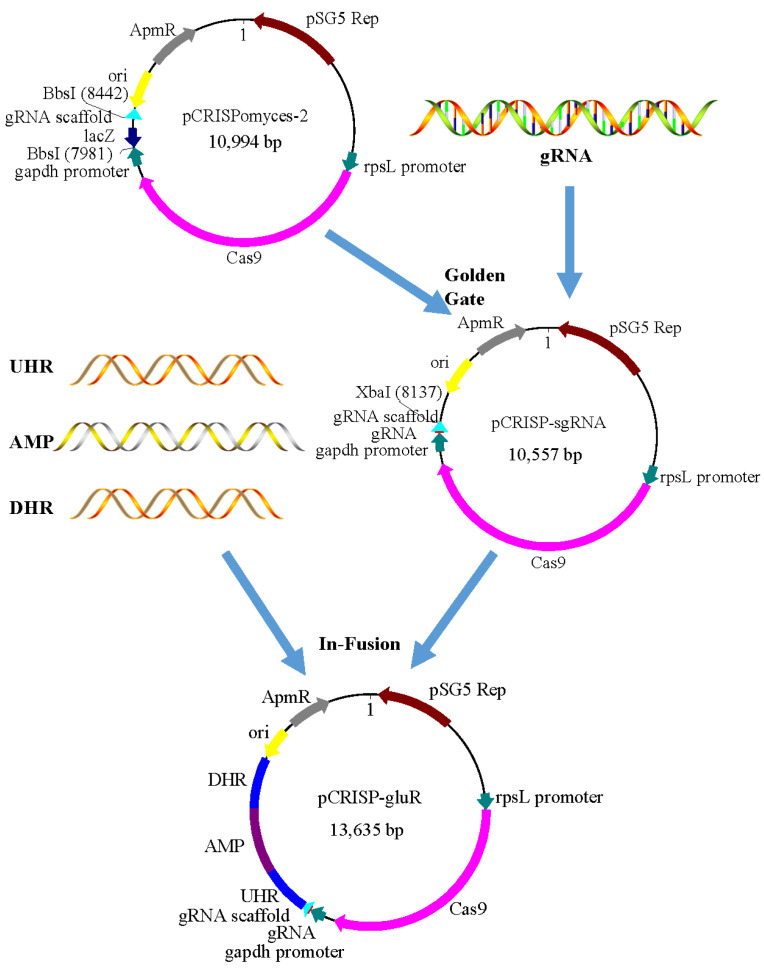 Figure 5
Figure 5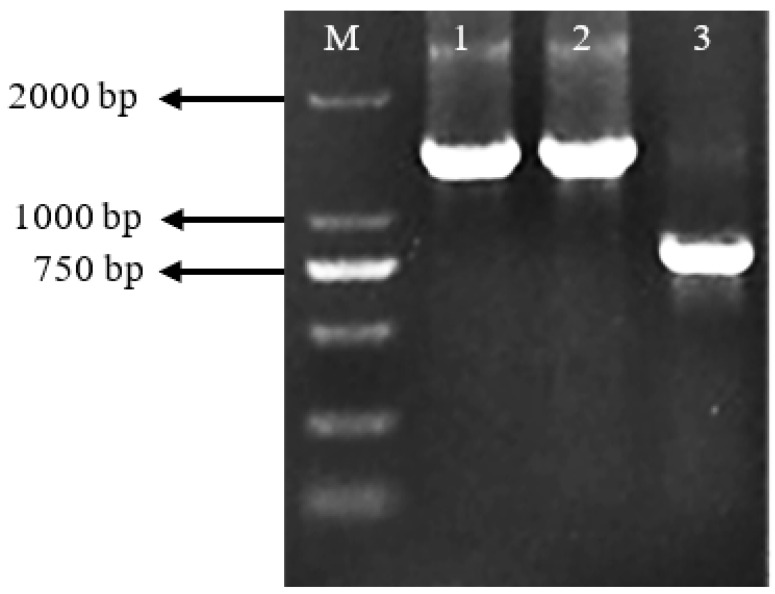 Figure 6
Figure 6 Figure 7
Figure 7- —National Natural Science Foundation of China
- —Open Project Program of State Key Laboratory of Food Nutrition and Safety, Tianjin University of Science & Technology
- —Project Program of Key Laboratory of Industrial Fermentation Microbiology, Ministry of Education, and Tianjin Key Laboratory of Industrial Microbiology
- —Youth Program of Shandong Provincial Natural Science Foundation
- —Tianjin Graduate Research Innovation Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Natural Products and Biosynthesis · Plant-Microbe Interactions and Immunity · Fungal and yeast genetics research
1. Introduction
Aspergillus flavus is a saprophytic filamentous fungus that is widespread in the environment, and is a pathogen commonly found in food and agricultural products [1,2]. A. flavus produces aflatoxins (AFs; B1, B2, G1, G2, M1, and M2), which are among the most toxic and carcinogenic compounds known to humans [3]. The effects of aflatoxins in animals include nephrotoxicity, hepatotoxicity, mutagenesis, teratogenesis, and immunosuppression [4]. Aflatoxin contamination is a major global public health concern despite the availability of multiple control strategies [5,6].
Such contamination is especially prevalent in nuts, grains, and dried fruits [7]. In the context of food processing, the widespread use of chemical fungicides in combating A. flavus and aflatoxin contamination has raised concerns due to their negative impacts on human health, including food safety, environmental pollution, plant toxicity, and chemical resistance [8,9,10]. By contrast, biological control relies on bioactive secondary metabolites from plants and microbes, offering an efficient, safe, and environmentally friendly alternative that has been shown to effectively suppress fungal pathogens [11,12,13].
mVOCs represent an environmentally friendly alternative to chemical fungicides. Being naturally volatile, carbon-based compounds, mVOCs disperse easily under ambient conditions, making them particularly suitable for agricultural applications [14,15,16,17,18]. These agents demonstrate broad-spectrum efficacy against a range of pathogens, addressing both increasing fungal resistance and environmental concerns associated with conventional fungicides [19]. Extensive research has validated the antimicrobial properties of mVOCs [20]. For instance, Muscodor albus and Streptomyces sp.—prolific producers of these compounds—have demonstrated substantial activities that prevent microbial spoilage and contamination in crops such as pistachios, dried figs, Indian senna, corn grains, and peanuts [21,22,23,24]. Among these mVOCs, dimethyl trisulfide has been reported to inhibit A. flavus growth and reduce aflatoxin production, which is critical for ensuring food safety and quality [25,26]. Furthermore, Yang et al. (2019) demonstrated that dimethyl trisulfide produced by S. alboflavus TD-1 effectively suppressed A. flavus, supporting its practical use as a biopesticide and confirming its role in biological control strategies [25]. This research highlights the potential of mVOCs for food safety management and indicates that mVOC-based inhibition could be explored as a low-residue strategy for controlling A. flavus during postharvest handling and grain storage, potentially reducing dependence on chemical preservatives [20,27].
Given the central role of Streptomyces sp. in mVOC production, elucidating their regulatory mechanisms is essential for improving biocontrol strategies. Streptomyces sp. are Gram-positive, filamentous soil bacteria characterized by high genomic guanine–cytosine (GC) content. They exhibit complex life cycles and produce a wide range of secondary metabolites with applications in medicine, industry, and agriculture [4,28,29]. Streptomyces sp. possess two-component systems (TCSs) that play a crucial role in regulating both primary and secondary metabolism [30]. These systems encompass mtrA/B, ragK/G, phoP/R, glnR, and cssR/S. Additionally, they influence the synthesis of bioactive molecules [31,32,33,34,35].
Beyond serving as a nitrogen donor in primary metabolism, glutamine also functions as a nutritional supplement that can modulate microbial physiology [36]. In microorganisms such as Streptomyces, glutamine availability influences nitrogen assimilation, amino acid biosynthesis, and secondary metabolism through regulatory systems including glnR and gluR/K [37]. As a signaling molecule, glutamine can activate or repress gene networks related to the synthesis of bioactive compounds, including antibiotics and volatile metabolites. Supplementation with glutamine has been reported to enhance growth, optimize metabolic flux toward secondary metabolites, and improve biocontrol efficacy against plant pathogens [38]. Given these properties, glutamine holds potential as a simple, cost-effective additive to boost the antifungal activity of microbial agents in agricultural applications [39].
This study investigated how exogenous glutamine supplementation influences mVOC biosynthesis in S. alboflavus TD-1 and enhances its antifungal activity against A. flavus, with a particular focus on the GluR-mediated regulatory pathway. The findings provide new insights into glutamine as a metabolic modulator for improving mVOC-based biocontrol for suppressing A. flavus growth. This is consistent with the increasing need for natural and eco-friendly solutions in the food industry, highlighting the significance of our research within the wider scope of food microbiology and public health.
2. Materials and Methods
2.1. Microorganisms
S. alboflavus TD-1 was originally isolated from soil in Tianjin, China, and identified as described previously (GenBank accession No. JX915780) [40]. This strain was maintained on Gause’s synthetic agar slants with 20% glycerol and stored at −80 °C for long-term preservation. Before experiments, cultures were grown on fresh Gause’s synthetic agar slants at 28 °C for 5 days. Mature colonies were suspended in 10 mL of sterile water containing 0.01% (v/v) Tween 80, gently scraped with a sterile cotton swab, and centrifuged at low speed. For mVOC production, 10 g of autoclaved wheat bran was placed in 250 mL conical flasks, inoculated with 2 mL of spore suspension (1 × 10^6^ sp-ore/mL), and incubated statically at 28 °C for 8 days.
A. flavus (CICC 2219), originally isolated from soy sauce residue, was obtained from the China Center of Industrial Culture Collection (Beijing, China) and cultured on PDA plates at 28 °C for 3 days. The culture was maintained at 4 °C. Conidia were harvested in 10 mL of sterile 0.05% Tween 80, filtered through two layers of autoclaved gauze to remove mycelia, and adjusted to 1 × 10^6^ conidia/mL with a hemocytometer.
2.2. Effects of S. alboflavus TD-1 mVOCs on Conidial Germination and Mycelial Biomass of A. flavus
The antifungal activity of S. alboflavus TD-1 metabolites against A. flavus was assessed using a dual-Petri dish method [41]. Two 60 mm Petri dishes were placed face-to-face and sealed with two layers of Parafilm. The lower dish contained S. alboflavus TD-1 grown on Gause’s synthetic agar supplemented with 0, 20, 40, 60, 80, or 100 mM glutamine. In this study, antifungal efficacy was evaluated using growth-related endpoints (conidial germination and dry mycelial biomass). Aflatoxin production was not quantified under mVOC exposure in this study.
For the conidial germination assay, the upper dish contained 5 mL PDA inoculated with 100 μL of an A. flavus conidial suspension (1 × 10^6^ conidia/mL). After incubation at 28 °C for 3 days, inhibitory effects were evaluated by counting conidia to determine the inhibition of conidial germination.
To determine the dry mycelial biomass, the upper dish contained 5 mL PDA inoculated with a 6 mm agar plug of A. flavus. After incubation under the same conditions (28 °C, 3 days), the mycelial mat was harvested and dried at 60 °C to constant weight. All treatments were performed in triplicate. The inhibition rate of conidial germination was calculated as follows:
where A_c_ is the number of conidia in the control, and A_t_ is the number of conidia during testing.
2.3. mVOCs Extraction and GC-MS Analysis
To analyze mVOCs associated with antifungal activity, S. alboflavus TD-1 was cultured on Gause’s synthetic agar supplemented with 0, 20, 40, 60, 80, or 100 mM glutamine in the bottom plate of a sealed dual-Petri dish system. Cultures were incubated statically at 28 °C for 8 days. mVOCs were extracted, separated, and identified, and their differential abundance (log2 (EG/CG)) was determined using headspace solid-phase microextraction (HS-SPME) coupled with gas chromatography–mass spectrometry (GC-MS).
A 2 cm SPME fiber (50/30 μm DVB/CAR/PDMS, Supelco, Bellefonte, PA, USA) was exposed to the headspace for 30 min at 45 °C to adsorb volatiles. The fiber was desorbed in the injection port of a GC/MSQP 2010 Ultra (Shimadzu, Kyoto, Japan) at 250 °C for 2 min in splitless mode. A VF-5MS capillary column (30 m × 0.25 mm i.d., 0.25 μm film; Varian Inc., Palo Alto, CA, USA) was used for separation and identification of volatiles [42]. Helium was used as the carrier gas at a constant flow of 1 mL/min. The following oven program was used: 40 °C for 3 min, increased to 150 °C at 4 °C/min, then ramped to 250 °C at 8 °C/min, and held for 6 min. The mass spectrometer was operated in electron impact (EI) mode at 70 eV, scanning from m/z 43 to 500. Compounds were identified by comparing mass spectra with the NIST 2011 Mass Spectral Library and further confirmed by comparing calculated retention indices (RI), determined using C7–C30 n-alkanes under identical conditions, with values reported in the literature.
2.4. Total RNA Isolation, RNA Sequencing, and Analysis
Mycelia were collected by filtering cultures through sterile gauze, washed with sterile PBS, and blotted dry on sterile filter paper. The harvested mycelium was immediately frozen in liquid nitrogen and stored at –80 °C until RNA extraction. Total RNA was extracted using Trizol Reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) according to the manufacturer’s instructions, and RNA quality and concentration were assessed with an Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA, USA) [43].
Ribosomal RNA was removed using the Ribo-off rRNA Depletion Kit (Vazyme Biotech, Nanjing, China). Strand-specific, indexed cDNA libraries were prepared using the SMARTer Stranded RNA-Seq Kit (TaKaRa Bio USA, Mountain View, CA, USA). Libraries were sequenced on an Illumina platform (Illumina, San Diego, CA, USA). Subsequent quality control, de novo assembly (Trinity v2.0.6), and annotation were performed as described by Grabherr et al., using BLASTx (NCBI BLAST+ v2.15.0) against the NR database and mapping to GO, KEGG, Swiss-Prot, and eggNOG databases [44].
Gene expression levels were normalized as log_2_-transformed FPKM values and analyzed using RSEM v1.3.0. Differential expression analysis was conducted using DESeq2 (v1.20.0, Bioconductor, New York, NY, USA) [45]. Genes with a fold change >2 and adjusted p-value < 0.05 were defined as differentially expressed (DEGs). GO and KEGG pathway enrichment analyses were performed to uncover the functions of DEGs.
2.5. qRT-PCR Analysis
To validate the RNA-Seq data, seven genes involved in mVOC biosynthesis were randomly selected for qRT-PCR analysis. Reactions were performed in a 25 µL volume containing 1 µL cDNA (<100 ng) on an Mx3000p instrument (Stratagene, La Jolla, CA, USA). Gene expression was normalized to 16S rRNA, and relative expression was calculated using the 2^−ΔΔCT^ method. Primer specificity was verified with melting curve analysis, and all reactions displayed a single sharp peak. All reactions included three biological and three technical replicates. Statistical significance was assessed using Student’s t-test (p < 0.05). Primers and reference genes are listed in Table 1.
2.6. CRISPR/Cas9-Mediated Deletion of gluR in S. alboflavus TD-1
E. coli DH5α was used as the host strain for cloning and plasmid construction. E. coli ET12567/pUZ8002 was used as the nonmethylating plasmid donor for intergeneric conjugation with S. alboflavus TD-1. The replicative vector pCRISPomyces-2, which harbors an apramycin resistance marker, was used as the CRISPR/Cas9 genome-editing tool. The plasmid pUC19 was used as the template for amplification of the ampicillin resistance gene. All strains and plasmids were obtained from an in-house repository.
The CRISPR/Cas9 editing vector pCRISPomyces-2 was constructed based on the method previously described by Cobb [46]. The sgRNA targeting gluR was cloned using Golden Gate assembly, and ~1 kb homologous arms flanking the gene were assembled by Gibson Assembly as described by Zeng [47]. The plasmid was transferred into S. alboflavus TD-1 by intergeneric conjugation using E. coli ET12567/pUZ8002, following the protocol outlined in Practical Streptomyces Genetics.
Genomic DNA from exconjugants was extracted, and PCR was performed to confirm integration. Primers gluR-F and gluR-R were used to verify replacement of gluR with the Amp gene, yielding an expected 778 bp PCR product. Another pair of primers, Amp-F and Amp-R, was used to verify that the Amp gene was present in exconjugants. The PCR amplification product was sequenced and compared with the GenBank reference sequence.
2.7. Assay for Inhibitory Effects of mVOCs on A. flavus
The inhibitory effects of mVOCs from S. alboflavus TD-1 and the gluR deletion mutant (TD-1 ΔgluR) on A. flavus were evaluated using a two-compartment Petri dish system to create a sealed co-culture environment. In the lower compartment, wheat bran cultures inoculated with either S. alboflavus TD-1 or TD-1 ΔgluR were incubated at 28 °C for 8 days to allow mVOC accumulation. The upper compartment contained 5 mL PDA inoculated in the center with a 6 mm plug of actively growing A. flavus. The plates were assembled vertically to form a sealed chamber (~75 mL headspace) and immediately sealed with two layers of Parafilm to prevent mVOC leakage. The assembly was incubated at 28 °C for 3 days. After incubation, the radial growth of A. flavus colonies was measured and compared between treatments. All experiments were conducted in triplicate. The inhibition rate was calculated as follows:
where D_c_ is the colony diameter of the control, and D_t_ is that of the treatment.
2.8. Statistical Analysis
Statistical analyses were performed using SPSS Statistics 22.0 (IBM, Armonk, NY, USA). Data are presented as mean ± standard deviation (SD). Differences among multiple groups were assessed via one-way analysis of variance (ANOVA). When significant effects were detected, post hoc comparisons were conducted using Fisher’s least significant difference (LSD) test. A p < 0.05 was considered statistically significant. Figures were prepared using Origin 9.0 (Origin Lab, Northampton, MA, USA).
3. Results
3.1. Effect of Glutamine on Antifungal Activity and mVOC Production in S. alboflavus TD-1
All glutamine treatments significantly inhibited A. flavus relative to the 0 mM control, as reflected by reduced conidial germination and decreased dry mycelial biomass (p < 0.05; Figure 1). The strongest effect was observed at 40 mM glutamine, where relative conidial germination decreased to 31.0% of the control (=69.0% inhibition) and relative mycelial biomass decreased to 35.5% of the control (=64.5% inhibition). Increasing glutamine beyond 40 mM did not further improve inhibition (p > 0.05 compared with 40 mM), indicating a plateau in the growth-suppression effect under the tested conditions.
To determine whether the enhanced antifungal activity was associated with increased mVOC production, headspace VOCs were profiled using GC-MS from cultures grown in the same dual-Petri dish setup, but incubated for 8 days to ensure sufficient mVOC accumulation. Across all treatments, a total of 21 mVOCs were identified, spanning hydrocarbons, ketones, terpenoids, alcohols, ethers, and other compounds (Table 2).
GC-MS profiling identified 21 VOCs, including hydrocarbons, ketones, terpenoids, and ethers (Table 2). Notably, supplementation with 40 mM glutamine led to marked increases in four VOCs with reported antifungal activity: dimethyl trisulfide, o-anisidine, anisole, and 1,5-cyclooctadiene. Their abundances increased 1.55- to 10.56-fold compared with the control, with the largest increases noted for 1,5-cyclooctadiene (10.55-fold) and dimethyl trisulfide (6.17-fold) (Table 2).
3.2. Transcriptomic Response of S. alboflavus TD-1 to Glutamine Supplementation
Two sequencing libraries were prepared from the control group (CG) and the glutamine-supplemented group (EG) to examine the transcriptomic response of S. alboflavus TD-1 to glutamine addition. The total mRNA reads for CG and EG were 43,292,342 and 44,852,360, respectively. The raw reads had an average length of 380 bp, and total data output exceeding 6.5 Gb for each group. Quality assessment showed that more than 92% of bases had Q30 scores (>99.9% base call accuracy), and more than 96% achieved Q20 scores (>99% accuracy).
Differential gene expression analysis was performed using the DESeq2 package, applying a threshold of q-value < 0.05 and |log_2_FoldChange| > 1. A total of 283 differentially expressed genes (DEGs) were identified in the EG compared to CG, including 173 upregulated and 110 downregulated genes.
GO analysis showed significant enrichment in biological processes such as urease activity, oxidoreductase activity, urea metabolic process, and urea catabolic process (Figure 2A). KEGG pathway analysis revealed 20 significantly enriched pathways, including amino acid metabolism, nitrogen metabolism, atrazine degradation, sulfur metabolism, and fatty acid metabolism (Figure 2B).
Based on GO and KEGG annotations, several enriched pathways—particularly those involved in nitrogen metabolism, amino acid metabolism, fatty acid metabolism, and sulfur metabolism—have been reported to provide precursors for the biosynthesis of mVOCs, including nitrogen- or sulfur-containing volatiles as well as lipid-derived compounds. These associations suggest potential links between glutamine supplementation and modulation of metabolic routes that could influence mVOC biosynthesis (Table 3, Figure 2C). However, these relationships are inferred from pathway annotation rather than direct functional evidence, and further experimental validation is required to confirm their specific roles in mVOC production.
To assess the reliability of the RNA-Seq data, seven DEGs (cutR, glnA, glnB, gluR, narK, trpE, and ureA) were randomly selected for validation by qPCR. The expression patterns of these genes were highly consistent with the RNA-Seq results (Figure 3, R^2^ = 0.97, p < 0.01). This strong correlation validated the robustness and accuracy of the transcriptomic analysis.
3.3. CRISPR/Cas9-Mediated Deletion of gluR
The CRISPR/Cas9 vector pCRISPomyces-2 was used to delete the gluR gene and generate the mutant strain TD-1 ΔgluR carrying the ampicillin resistance gene. The workflow of CRISPR–Cas9 gluR knockout is shown in Figure 4.
Exconjugants were verified by PCR using gluR-F/gluR-R and Amp-F/Amp-R primers. In the wild-type strain, a 778 bp fragment of gluR was amplified, while in the ΔgluR mutant, a 1394 bp fragment (1227 bp Amp gene + 167 bp flanking sequence) was obtained (Figure 5). DNA sequencing confirmed successful replacement of gluR with the Amp gene.
3.4. Effects of gluR Deletion on Phenotype, Antifungal Activity, and mVOC Profiles
Deletion of the gluR gene in S. alboflavus TD-1 resulted in noticeable phenotypic changes. The ΔgluR mutant displayed altered colony coloration, with reduced red pigment production and more vigorous aerial mycelium (Figure 6A1,A2). In contrast, the general morphology of the mycelial network remained comparable to that of the wild-type strain (Figure 6B1,B2).
Functionally, the inhibitory activity of mVOCs produced by the ΔgluR mutant against A. flavus was significantly reduced. The wild-type strain inhibited A. flavus growth by 68.96%, whereas inhibition by the ΔgluR mutant was only 50.57% (Figure 6C1,C2), highlighting the importance of gluR in regulating antifungal capacity.
GC-MS analysis further revealed substantial differences in mVOC composition between the two strains (Table 4). In the ΔgluR mutant, concentrations of sulfides, ethers, most terpenes, and ketones were markedly reduced, whereas hydrocarbons, aldehydes, and some terpenes were elevated relative to the wild-type strain. Importantly, five antifungal mVOCs—anisole, dimethyl trisulfide, o-anisidine, 1,5-cyclooctadiene, and β-pinene—were consistently lower in the mutant, together with other bioactive compounds such as D-limonene, 2-methylisoborneol, and isoledene.
Taken together, these results demonstrate that gluR deletion not only alters the colony phenotype of S. alboflavus TD-1 but also reduces its antifungal activity, primarily through downregulation of key antifungal mVOCs. This finding indicates that gluR plays a dual regulatory role in mVOC biosynthesis, exerting both positive and negative effects on metabolite production, with its positive regulation of compounds such as dimethyl trisulfide and o-anisidine being particularly critical for effective inhibition of A. flavus.
4. Discussion
A. flavus is a major postharvest concern because it readily contaminates cereals, nuts, and other commodities and can lead to aflatoxin accumulation, posing a direct food safety risk [17,48]. mVOCs produced by biocontrol microbes have therefore attracted growing interest as low-residue antifungal agents, particularly in enclosed or semi-enclosed settings such as storage and packaging systems [18,19]. A persistent practical challenge is that mVOC emission is highly sensitive to culture conditions, and is often difficult to reproduce across different media and environments. In this study, our results indicate that glutamine supplementation can be used as a simple lever to enhance the antifungal mVOCs of S. alboflavus TD-1 under the conditions tested. In addition, the results also suggest that GluR is involved in linking nutrient status to mVOC-mediated antagonism [20].
This enhancement is supported by consistent phenotypic outcomes. At 40 mM glutamine, TD-1 showed the strongest inhibition, suppressing A. flavus conidial germination by 69.0% and reducing dry mycelial biomass by 64.5% relative to the control. Together, these readouts reflect both early developmental arrest and reduced vegetative accumulation, which are directly relevant to limiting fungal establishment during storage and handling.
An important feature of the response was its non-linearity. Antifungal performance peaked at 40 mM glutamine and did not improve further at concentrations of 60 mM and above. This pattern is consistent with the view that glutamine acts not only as a nitrogen source but also as a regulatory metabolite capable of reshaping secondary metabolism [16,36]. From an application standpoint, the finding argues for dose optimization rather than simple nutrient maximization, and it raises the possibility that excessive nitrogen signaling may shift metabolism away from the mVOC subset most responsible for antifungal activity. At least two explanations are plausible. Nitrogen-sufficiency signaling could trigger feedback regulation that alters carbon–nitrogen allocation and limits precursor supply for particular mVOC pathways [49]. In addition, higher glutamine levels may alter the local physicochemical environment in ways that broadly reprogram secondary metabolism. These possibilities are not mutually exclusive, and future work that combines glutamine gradients with intracellular metabolite measurements and absolute mVOC quantification should help clarify whether the decline at higher concentrations reflects precursor limitation, physiological shifts, or both [50].
mVOC profiling provides a useful bridge between glutamine supplementation and enhanced antifungal activity. TD-1 produced a defined mVOC repertoire, and glutamine mainly altered the relative abundance of multiple compounds rather than generating an entirely new mVOC profile. It is important to note that the changes reported here are relative and derived from GC-MS peak-area comparisons between experimental and control conditions (log2 (EG/CG)), rather than absolute headspace concentrations. Under treatment with 40 mM glutamine, several compounds increased markedly, with the largest changes reaching approximately 12.47-fold. Notable increases were observed for 1,4-dimethyladamantane, 1,5-cyclooctadiene, and azulene, along with other hydrocarbons and aromatics. Glutamine also elevated compounds that have been repeatedly associated with antimicrobial activity in microbial volatile compounds, including dimethyl trisulfide as well as anisole and o-anisidine [26,51].
mVOCs typically act as blends, and antifungal outcomes often reflect mixture effects rather than the action of a single compound. The improved antagonism observed here is therefore likely to result from both the increased abundance of inhibitory mVOCs reported in the literature and a shift in the overall blend that changes how the mixture interacts with the target fungus. Nevertheless, these mixture-level mechanisms remain inferential at present. The most direct way to strengthen causal attribution, particularly in a food context, is to test candidate mVOCs at headspace-relevant levels and then examine whether reconstructed mixtures recapitulate the activity of the natural blend. Linking inhibition to absolute emissions, rather than relative changes alone, will also be essential [19,20].
Transcriptomic data provide a broader metabolic rationale for the observed volatile compounds remodeling. Glutamine induced 283 differentially expressed genes, with enrichment in pathways related to amino acid metabolism, sulfur metabolism, fatty acid metabolism, and transport or regulatory functions. Several genes associated with nitrogen assimilation and urea-related metabolism were downregulated, consistent with a nitrogen-sufficiency state that may redistribute flux. The strong induction of sulfonate utilization and transport genes, such as ssuC and ssuD, offers a plausible connection to increased sulfur volatiles including dimethyl trisulfide. Changes in fatty acid metabolism genes, including fabH and fabG, are also consistent with shifts in precursors that contribute to hydrocarbon- and ketone-type mVOCs [34,52]. Although transcriptomics cannot by itself assign each mVOC to a specific biosynthetic route, it highlights sulfur handling and lipid metabolism as high-priority nodes for targeted validation [30,49].
Our data further support a role of GluR in glutamine-responsive regulation of antifungal mVOC output. Glutamine increased gluR expression, and deletion of gluR attenuated mVOC-mediated antagonism, with inhibition decreasing from 68.96% in the wild-type strain to 50.57% in ΔgluR. GC-MS comparisons were consistent with this phenotype and indicated broad mVOC shifts in the mutant, including reduced levels of candidate antifungal compounds such as dimethyl trisulfide and anisole. These findings support a working model in which GluR contributes to a nutrient-responsive regulatory program that promotes production and/or emission of an antifungal mVOC blend [37]. In addition, the mechanistic boundary should be stated clearly. The ΔgluR phenotype may reflect direct regulatory control of mVOC-associated pathways, but it could also arise indirectly through altered growth or emission capacity. Complementation of ΔgluR, normalization of mVOC emissions to biomass, and identification of downstream targets would provide the most convincing confirmation [52].
From the perspective of foods, translating these findings to real commodities requires careful attention to matrix and headspace effects [14,38]. mVOC concentrations and composition in sealed Petri dish systems can differ substantially from those in grain bulks or packaged foods because ventilation, adsorption to matrices, and humidity-dependent partitioning all influence effective exposure. In addition, some mVOCs, particularly sulfur-containing and aromatic compounds, may affect sensory properties [20,27]. Any practical storage or packaging application should evaluate not only antifungal efficacy but also sensory acceptability, regulatory constraints, and safety, and should directly quantify aflatoxin outcomes where relevant. Another practical question is how reliably glutamine can steer mVOC output in complex substrates, where it may be consumed by other microbes or interact with the commodity matrix. Moving forward, it will be important to quantify key mVOCs absolutely across glutamine levels and then test performance directly in commodity systems under storage-relevant temperature and humidity [23,38]. Importantly, inhibition of A. flavus growth does not necessarily predict aflatoxin outcomes because aflatoxin biosynthesis can be uncoupled from biomass accumulation, and stress conditions may suppress growth while maintaining or even increasing toxin production. In this study, we did not quantify aflatoxins under VOC exposure; therefore, our conclusions are limited to growth suppression and the glutamine/GluR-dependent modulation of mVOC profiles. Direct measurement of aflatoxins in mVOC-exposed cultures and in commodity-relevant storage systems will be necessary to determine whether the observed growth inhibition is accompanied by decreased aflatoxin production/accumulation under mVOC exposure.
Taken together, these results show that glutamine can modulate the TD-1 volatile compounds in a way that improves antifungal performance under controlled conditions and that GluR is implicated as a regulatory node linking nutrient sensing to mVOC-mediated antagonism. The work provides a mechanistic starting point for improving the robustness of mVOC-based suppression of A. flavus and outlines the practical steps needed to translate laboratory observations into postharvest applications.
5. Conclusions
This study demonstrated that glutamine has a significant influence on mVOC production in S. alboflavus TD-1. The gluR gene was found to positively regulate the biosynthesis of sulfides, ethers, most terpenes, and ketones, while exerting negative regulatory effects on hydrocarbons, aldehydes, and certain terpenes. Transcriptomic analysis combined with the gluR deletion mutant revealed that glutamine regulates amino acid and nitrogen metabolism through the GluR two-component system, thereby modulating mVOC biosynthesis and enhancing the biocontrol efficacy of S. alboflavus TD-1 against A. flavus. Taken together, these findings highlight the dual role of glutamine as a metabolic substrate and regulatory signal, linking nutrient sensing to secondary metabolism and antifungal activity. Beyond advancing our mechanistic understanding, this work offers promising directions for developing glutamine-enhanced mVOCs as a sustainable strategy to suppress A. flavus growth, with aflatoxin outcomes requiring direct validation in commodity-relevant systems. Nevertheless, further research is needed to validate these findings under in vivo and field conditions, to optimize application methods, and to assess ecological safety and scalability.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wang P. Ma L. Jin J. Zheng M. Pan L. Zhao Y. Sun X. Liu Y. Xing F. The anti-aflatoxigenic mechanism of cinnamaldehyde in Aspergillus flavus Sci. Rep.201991049910.1038/s 41598-019-47003-z 31324857 PMC 6642104 · doi ↗ · pubmed ↗
- 2Gilbert M.K. Mack B.M. Lebar M.D. Chang P.-K. Gross S.R. Sweany R.R. Cary J.W. Rajasekaran K.J.J.o.F. Putative core transcription factors affecting virulence in Aspergillus flavus during infection of maize J. Fungi 2023911810.3390/jof 9010118 PMC 986128036675939 · doi ↗ · pubmed ↗
- 3Wang H. Liu M. Zhang Y. Zhao H. Lu W. Lin T. Zhang P. Zheng D. Rapid Detection of Aspergillus flavus and Quantitative Determination of Aflatoxin B(1) in Grain Crops Using a Portable Raman Spectrometer Combined with Colloidal Au Nanoparticles Molecules 202227528010.3390/molecules 2716528036014519 PMC 9414248 · doi ↗ · pubmed ↗
- 4Sun Y. Yao Z. Long M. Zhang Y. Huang K. Li L. Alveolar Macrophages Participate in the Promotion of Influenza Virus Infection by Aflatoxin B 1 at an Early Stage Toxins 2023156710.3390/toxins 1501006736668886 PMC 9863124 · doi ↗ · pubmed ↗
- 5Abdelhamid H.N. Mahmoud G.A.E. Sharmoukh W. A cerium-based MO Fzyme with multi-enzyme-like activity for the disruption and inhibition of fungal recolonization J. Mater. Chem. B 202087548755610.1039/D 0TB 00894 J 32716461 · doi ↗ · pubmed ↗
- 6Hu Z. Yuan K. Zhou Q. Lu C. Du L. Liu F.J.F.c. Mechanism of antifungal activity of Perilla frutescens essential oil against Aspergillus flavus by transcriptomic analysis Food Control 202112310770310.1016/j.foodcont.2020.107703 · doi ↗
- 7Dini I. Alborino V. Lanzuise S. Lombardi N. Marra R. Balestrieri A. Ritieni A. Woo S.L. Vinale F. Trichoderma Enzymes for Degradation of Aflatoxin B 1 and Ochratoxin A Molecules 202227395910.3390/molecules 2712395935745082 PMC 9231114 · doi ↗ · pubmed ↗
- 8Ahmad M.F. Ahmad F.A. Alsayegh A.A. Zeyaullah M. Al Shahrani A.M. Muzammil K. Saati A.A. Wahab S. Elbendary E.Y. Kambal N.J.H. Pesticides impacts on human health and the environment with their mechanisms of action and possible countermeasures Heliyon 202410 e 2912810.1016/j.heliyon.2024.e 2912838623208 PMC 11016626 · doi ↗ · pubmed ↗