Prevalence and Characterization of Methicillin-Resistant Staphylococcus aureus from Animals, Retail Meats and Market Shopping Vehicles in Shandong, China
Ting-Yu Yang, Chong-Xiang Sun, Junjie Wang, Zhiyuan You, Hao Wang, Kelan Yi, Feng-Jing Song, Bao-Tao Liu

TL;DR
This study found that MRSA is commonly present in retail meats and shopping carts in China, posing a public health risk due to its potential to spread antibiotic-resistant infections.
Contribution
This is the first report of widespread MRSA contamination in shared shopping vehicles in China.
Findings
MRSA was found in 7.5% of shared shopping vehicles, 2.2% of retail meats, and 44.4% of supermarkets had MRSA-positive shopping vehicles.
MRSA isolates from meats and shopping vehicles were genetically similar to those from humans and animals, suggesting cross-species transmission.
Retail meats carried ST6-MRSA, a strain linked to food poisoning outbreaks in China.
Abstract
Staphylococcus aureus has been recognized as an important foodborne pathogen and methicillin-resistant S. aureus (MRSA) can cause fatal infections worldwide. Of great concern is that MRSA have been found in animals and non-healthcare settings; however, knowledge about the prevalence and genetic characteristics of S. aureus, especially MRSA from animals, retail meats and market shared shopping vehicles in the same district, is limited. In this study, we collected 423 samples including handrail swabs (n = 226) of shopping trolleys and baskets from 18 supermarkets, retail meats (n = 137) and swine nasal swabs (n = 60) between 2018 and 2020 in China. S. aureus isolates were isolated and identified by PCR, and then the mecA was used to confirm the MRSA. The antibiotic resistance and virulence genes among S. aureus were also analyzed, followed by whole genome sequencing (WGS). S. aureus…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
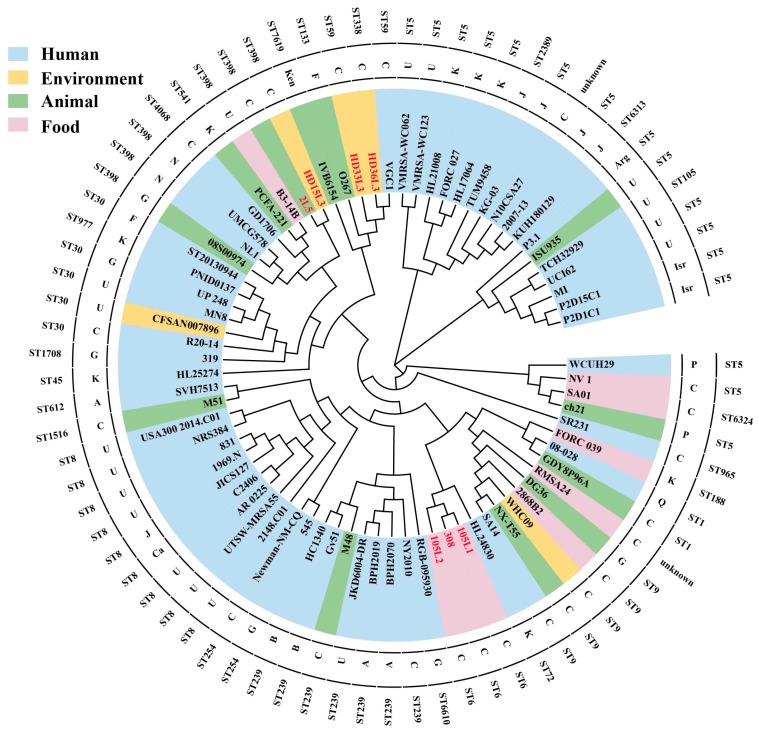 Figure 1
Figure 1- —Shandong Provincial Natural Science Foundation
- —Scientific and Technological Projects of Qingdao
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntimicrobial Resistance in Staphylococcus · Bacterial biofilms and quorum sensing · Milk Quality and Mastitis in Dairy Cows
1. Introduction
Antimicrobial resistance (AMR) is a critical One Health issue that has been linked to humans, animals and environments [1,2], and it often poses great threat to the health of both humans and animals [3]. Staphylococcus aureus (S. aureus) is a human pathogen and the presence of resistance genes exacerbates the risk of spreading antimicrobial resistances. S. aureus caused over 100,000 cases of bacteremia in the United States in 2017 [4]. In the European Union, it caused 434 foodborne outbreaks in 2015 and the estimated cost of a single outbreak of foodborne illness in a restaurant might exceed 1 million US dollars [5]. In China, S. aureus was one of the leading pathogens, responsible for causing 577 outbreaks, 9092 cases of illness and 3715 hospitalizations from 2010 to 2020 [6]. Methicillin-resistant Staphylococcus aureus (MRSA) can cause fatal infections worldwide [7], and 31.4% of all clinical isolates have been recognized as MRSA according to the China Antimicrobial Surveillance Network (CHINET) surveillance system in 2019 [8]. In most Asian countries, MRSA accounted for more than one-half of the hospital-related infections caused by S. aureus [9].
Besides those hospital-acquired MRSA (HA-MRSA) initially confined to hospitals, swine-derived ST398-type MRSA known as livestock-associated MRSA (LA-MRSA) was firstly reported to cause human infection in The Netherlands in 2005 [10]. The presence of MRSA strains in swine has been detected in several regions of China [11,12] and it is found that swine are important hosts for LA-MRSA. MRSA has also been isolated from cattle, horses, goats, rabbits, poultry, cats, dogs and food of animal origin [13,14,15,16], although the prevalence of MRSA in animals is low. Of particular concern is the fact that LA-MRSA carries AMR genes and can be transmitted between animals and humans [17]. The LA-MRSA may even cause human infections through direct or indirect contact with infected animals, posing a challenge to human health [18,19]. The presence of MRSA in the human food chain, especially in raw meats, has been reported [20,21]; however, the prevalence of MRSA varied between meats of different origins (such as beef, poultry meat and pork) and countries.
MRSA in food animals may not only cause animal diseases and contaminated food [22,23,24], but also contaminate farm environments, which are important vectors mediating the transmission of AMR or pathogens between animals and humans. Of great concern is that MRSA have been isolated and characterized in several non-farm environments such as public shared bicycles [25,26] and metro system [27]. The high volume of people in public areas and the survival of MRSA on public surfaces facilitate the transmission of Staphylococci including MRSA in such areas. Supermarket shopping is still the most common shopping form, and shared shopping vehicles in supermarkets have been an indispensable component of people’s daily life. Despite great convenience, the shared shopping vehicles could be a source of antimicrobial-resistant pathogenic bacteria, posing threat to public health. However, there remains a paucity of data regarding the distribution and features of MRSA from shopping vehicles in supermarkets. The association of S. aureus, especially MRSA from retail meats, shopping vehicles and animals, should be investigated, and the role of shopping vehicles in disseminating pathogens, especially MRSA, remains largely unknown.
China has huge amounts of supermarkets, consumers of raw meats and food animals, creating an ideal situation for the spread of S. aureus including MRSA. In this cross-sectional study, we aimed to elucidate the prevalence and molecular characteristics of S. aureus strains, particularly MRSA, among shopping vehicles within supermarkets, raw meats and swine in Shandong Province of China. The association of the MRSA isolates from different niches was also analyzed.
2. Materials and Methods
2.1. Sample Collection and Identification of Strains
A total of 423 samples from 5 cities (Qingdao, Zibo, Heze, Weifang, Dezhou) of Shandong province in China were collected between 2018 and 2020. These samples included swine nasal swabs (n = 60), raw meats (n = 137) and shared shopping vehicles within supermarkets (n = 226) (Table 1).
Nasal swabs were collected from 60 healthy pigs which were randomly selected from 2 pig farms in city Qingdao, China. Chicken meat (n = 94) and pork (n = 43) samples were purchased from nine supermarkets in city Qingdao. Each raw meat sample was aseptically homogenized and 25 g of homogenized sample was mixed with 225 mL Luria-Bertani (LB) broth (Hopebio, Qingdao, China) containing 7.5% sodium chloride and incubated at 37 °C for 16 to 24 h. The handrails of 170 shopping trolleys and the handles of 56 shopping baskets from 18 supermarkets in 5 cities were wiped with sterile cotton swabs soaked with saline. Of the 18 supermarkets, 6 were from those we collected meat samples in city Qingdao and 78 samples (60 from shopping trolleys and 18 from baskets) were collected. Three supermarkets were selected from each of the other four cities and there were 40, 39, 40 and 29 samples collected from city Heze, Zibo, Dezhou and Weifang, respectively (Table 2). Each swab was placed into sterile LB broth containing 7.5% NaCl and these samples were stored below 8 °C and processed within 24 h after sampling.
The LB broth with survived bacteria was diluted in series of 1:10 and 100 µL of appropriate dilution and was spread onto CHROMagar^TM^ Staph aureus medium (CHROMagar, Paris, France) and incubated for 24 h. The colonies with pink were suspicious S. aureus and one colony showing typical appearance was selected from each sample. DNA extraction was performed using the phenol–chloroform method as previously described, with minor modification [28]. Briefly, the bacteria in 325 μL TE buffer (10 mM Tris, 5 mM EDTA, pH 7.8) was cultured with 25 μL lysozyme solution (10 mg/mL) at 37 °C for 1 h. Lysis of cells was achieved by addition of 20 μL of SDS (20% SDS w/v in Tris (50 mM), EDTA (20 mM), pH 7.8) followed by adding 3 μL of proteinase K (20 mg/mL, Sigma, St. Louis, MO, USA) and further incubated at 37 °C for 1 h. Protein was precipitated by the addition of 200 μL NaCl (5 M) followed by centrifugation (8000 rpm) for 10 min. The supernatant was extracted with phenol/chloroform (1:1) followed by chloroform/isoamyl-alcohol (24:1) extraction. DNA was precipitated from the supernatant by adding 0.6 volumes of isopropanol and collected by centrifugation at 8000 rpm, 4 °C for 10 min. The pellet was washed with 70% ethanol and dried, followed by being resuspended in 1 mL of TE buffer. Identification of S. aureus was performed by using the nuc and femB genes as previously described [29,30].
2.2. Antimicrobial Susceptibility Testing
The antimicrobial susceptibilities of the obtained 49 S. aureus isolates to 11 antimicrobials were determined using the agar dilution method according to the guidelines of Clinical and Laboratory Standards Institute (CLSI) [31]. The 11 antimicrobial agents were as follows: penicillin, ciprofloxacin, levofloxacin, vancomycin, linezolid, gentamicin, kanamycin, clindamycin, erythromycin, tetracycline and florfenicol. The breakpoints for these antimicrobials were recommended by the CLSI [31]. The reference strain S. aureus ATCC 25923 was used for quality control.
2.3. Detection of mecA, Virulence Genes and Antibiotic Resistance Genes
All S. aureus isolates were further screened for the presence of mecA to confirm the MRSA [32]. Thirteen virulence genes encoding the Panton–Valentine leukocidin (lukF-PV/lukS-PV), enterotoxins (sea, seb, sec, sed, see), toxic shock syndrome toxin-1 (tsst-1), exfoliative toxins (eta, etb), hemolysin (hla, hlb) and the adhesion factors (fnbA, fnbB) were screened using PCR as described previously [33,34,35,36]. The common macrolide resistance genes erm(A), erm(B) and erm(C) among all S. aureus isolates were analyzed by PCR as previous described [37]. The presence of vancomycin resistance genes vanA and vanB were further investigated according to the previous method [38]. The oxazolidinones resistance genes cfr, poxtA and optrA were also screened as described previously [39]. The primers used in this study were listed in Table S1.
2.4. SCCmec Typing
SCCmec types and subtypes I, II, III, IVa, IVb, IVc, IVd and V of the MRSA isolates in this study were identified by multiplex-PCR using specific primers (Table S1), as previously described [40]. PCR conditions were set at 94 °C for 5 min, followed by 30 cycles of 94 °C for 30 s, 52 °C for 45 s and 72 °C for 90 s, ending with a final extension step at 72 °C for 10 min. The PCR amplicons were electrophoretically separated at 100 V/cm in a 1.5% agarose gel. MRSA isolates that could not be assigned to type I–V were defined as nontypeable (NT).
2.5. Whole-Genome Sequencing
All three MRSA isolates from retail meats and two randomly selected MRSA isolates from shopping vehicles in Qingdao were subjected to whole-genome sequencing (WGS). The methicillin-sensitive S. aureus (MSSA) isolate HD15L3 from shopping trolley in Qingdao and one ciprofloxacin-resistant MSSA isolate 2L5 from pig in Qingdao were also subjected to WGS. Briefly, total DNA of the seven S. aureus isolates was extracted using a TIAN amp Bacteria DNA kit (Tiangen, Beijing, China) and then sequenced using an Illumina HiSeq 2500 platform (Illumina, San Diego, CA, USA) with the 150 paired-end sequencing strategy. Illumina reads were quality checked using software FastQC version 0.11.9 (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/) (accessed on 10 June 2022). SPAdes v 3.12.0 was used to assemble the clean Illumina reads [41]. The assembled sequences were subjected to CGE (http://www.genomicepidemiology.org/services/) (accessed on 2 June 2024) to analyze the antibiotic resistance genes. Virulence genes were also analyzed using CGE (https://www.genomicepidemiology.org/services/) (accessed on 2 June 2024) and VFDB core dataset (http://www.mgc.ac.cn/VFs/main.htm) (accessed on 3 June 2024). Functional annotation was performed using the NCBI Prokaryotic Genome Annotation Pipeline server.
To track the phylogenomic relationships of the S. aureus isolates in our study and those reported from food, humans and animals, we used the seven genomes in this study and the assembled genomic sequences of 70 S. aureus isolates in the NCBI database to obtain the phylogenomic tree. The 77 genomes consisting of 56 MRSA isolates and 21 MSSA isolates included 12 genomes from animals, 51 from humans, 5 from environment and 9 from food (Table S2). SNPs were determined as we described previously [42]. The MRSA isolate HD33L3 from shopping trolley was set as reference. The phylogenetic tree was visualized using Evolview version 3 [43].
2.6. Statistical Analysis
Comparisons between genotype and antibiotic resistance frequencies in different populations (sample types, MRSA/MSSA) were performed using the Pearson Chi-square test in SPSS software (version 21). False discovery rate according to Benjamini–Hochberg procedure was applied to adjust for multiple comparisons. All tests of significance were two-tailed, and a p-value of ≤0.05 was considered to be statistically significant.
3. Results
3.1. Prevalence of S. aureus and MRSA
A total of 49 isolates were positive for both the nuc and femB genes and were classified as S. aureus. The overall prevalence of S. aureus among the 423 samples was 11.6% (Table 1). S. aureus was found in 18.3% of the swine samples, which was similar to that (14.6%, 20/137) in retail raw meats. The detection rate of S. aureus in chicken meats and pork was 17.0% (16/94) and 9.3% (4/43), respectively. Among the 226 samples from shopping vehicles, 18 (8.0%) harbored S. aureus and there was no significant difference between the isolation rate from shopping trolleys (7.6%, 13/170) and that from shopping baskets (8.9%, 5/56) (p > 0.05) (Table 1). MRSA isolates were identified in 20 samples in this study. Notably, 17 of the 226 (7.5%) shopping vehicles samples carried MRSA and the detection rate of MRSA in shopping trolleys and baskets was 7.1% and 8.9%, respectively. There were only three chicken meat samples that harbored MRSA, while no MRSA was found in pork and swine nasal swabs (Table 1).
Shopping vehicles with MRSA were found in all five cities (Table 2). Of the five cities, the highest detection rate of shopping vehicles carrying MRSA was found in Heze (17.5%, 7/40), followed by Weifang (13.8%, 4/29) (Table 2). Two of the three supermarkets in city Heze, Dezhou and Weifang harbored MRSA-positive samples. In total, 8 of the 18 supermarkets (44.4%) in this study possessed MRSA-positive shopping vehicle samples. Notably, the detection rates of MRSA in five supermarkets were above 14.3% and 28.6% of the shopping vehicles from market 9 in Heze carried MRSA (Table 2).
3.2. Antimicrobial Susceptibility and Antibiotic Resistance Genes
As shown in Table 3, 89.8% of the 49 S. aureus isolates were resistant to erythromycin, followed by clindamycin and penicillin. Resistance rates to ciprofloxacin, tetracycline, kanamycin and florfenicol were all above 30.0%. Notably, the resistance rate to ciprofloxacin and florfenicol among S. aureus isolates from animals was significantly higher than that from raw meats and that from shopping vehicles, respectively (p < 0.05) (Table 3). The resistance rate to kanamycin among S. aureus isolates from shopping vehicles was significantly higher than that from raw meats (p < 0.05) (Table 3). Among MSSA isolates, the resistance rate to ciprofloxacin, tetracycline, gentamicin, levofloxacin and florfenicol was significantly higher than that of MRSA isolates, respectively (p < 0.05) (Table 3).
Among the 49 S. aureus isolates, 44.9% and 40.8% carried macrolide resistance genes erm(B) and erm(C), respectively (Table 4). Resistance genes erm(A), vanA, vanB, cfr, poxtA and optrA were not found in this study. The detection rate of erm(B) among shopping vehicles S. aureus isolates was significantly higher than that from raw meats (p < 0.05), while the detection rate of erm(C) in S. aureus isolates among animals, raw meats and shopping vehicles was not significantly different (p > 0.05). Both the detection rate of erm(B) and erm(C) among MRSA isolates were not significantly different from MSSA isolates (p > 0.05).
3.3. Virulence Genes in S. aureus Isolates from Different Origins
Seven of the thirteen screened virulence genes were found in this study. Among the 49 isolates, 100.0%, 85.7% and 65.3% carried hla, hlb and fnbB, respectively, followed by eta (46.9%), sea (14.3%), seb (10.2%) and lukF-PV/lukS-PV (6.1%) (Table 4). The prevalence of enterotoxin genes (sea/seb) associated with food poisoning in S. aureus isolates was 22.4% (11/49) and enterotoxin genes were present in 45.0% of the 20 S. aureus isolates from retail meats. Five of the 20 MRSA (25.0%) carried enterotoxin genes. The detection rate of eta in S. aureus isolates from raw meats was significantly higher than that from shopping vehicles and that from animals (p < 0.05). The detection rate of sea among S. aureus isolates from raw meats was significantly higher than that from animals (p < 0.05). The detection rate of lukF-PV/lukS-PV and sea in MRSA isolates was significantly higher than that in MSSA isolates, respectively, while the detection rate of eta and fnbB in MSSA isolates was significantly higher than that in MRSA isolates, respectively (p < 0.05).
3.4. SCCmec Typing of MRSA Isolates and Whole-Genome Sequence Analysis
All the 20 MRSA isolates belonged to the SCCmec type IVa. WGS analysis showed that all three MRSA isolates (105L1, 105L2 and 308) from raw meats belonged to ST6 and possessed not only the mecA but also blaZ, erm(C) and grlA: S80F (Table 5). Unlike the three MRSA isolates from raw meats, both MRSA isolates (HD33L3, HD36L3) from shopping vehicles carried mecA, erm(B), aph(3′)-III and ant(6)-Ia. Compared to ST59 HD33L3, the ST338 type HD36L3 carried two additional resistance genes including tet(K) and blaZ. Both MSSA isolates (2L5, HD15L3) belonged to ST398 and carried 14 resistance genes encoding resistances to aminoglycosides, fluoroquinolones, tetracyclines, folate pathway antagonist, beta-lactams, amphenicols, macrolides, pleuromutilin and lincosamide (Table 5). An additional tet(K) (tetracycline resistance) was also found in isolate HD15L3.
All five sequenced MRSA isolates from raw meats and shopping vehicles harbored six to seven classes of virulence genes, including genes related to adherence, exoenzyme, exotoxin, effector delivery system, immune modulation, nutritional/metabolic factor and biofilm factor (Table 5). Only MRSA isolate 105L2 from raw meat carried all seven classes of virulence genes. Adherence gene (eap/map) was only found in the three MRSA isolates from raw meats, among which two lacked virulence genes for biofilm factor. Biofilm factors were found in five sequenced S. aureus isolates, including the three from shopping vehicles. Only MRSA isolate HD36L3 recovered from shopping baskets had the Panton-Valentine leucocidin toxin gene lukS-PV and the Staphylococcus enterotoxin B gene seb. All three MRSA from retail meats harbored the immune evasion cluster (IEC) with the sea and sak genes. Overall, 35 (n = 2), 32 (n = 2), 45 (n =1), 40 (n =1), and 33 (n = 1) virulence genes were detected in the seven WGS-sequenced S. aureus isolates.
A total of 23, 170 SNPs were obtained from the 77 S. aureus genomes of different sources and S. aureus isolates belonging to the same STs were clustered closely (Figure 1). All three ST6 MRSA isolates from raw meats in our study were clustered together and had a limited number of variations (362 SNPs), indicating that a close genetic relationship existed among these isolates, even for those from different samples or supermarkets. Notably, our three MRSA isolates from raw meats were also clustered closely with MRSA isolate HL24830 (CP082789) from human blood in South Korea. The MRSA isolate NX-T55 (CP031839) from animal in China showed less than 192 SNPs away from the MRSA 2868B2 (CP060141) in food, MRSA isolate WHC09 (CP077755) from animal farm bioaerosol and MRSA isolate SA14 (CP073012) from human in China. These results suggest that the exchange of MRSA isolates might occur among different niches and the MRSA in retail meats will pose a threat to public health.
Furthermore, both the MRSA isolates HD33L3 and HD36L3 from shopping vehicles in Qingdao supermarket were clustered with MRSA isolate VG1 (CP039448) from human blood and O267 (CP034102) from animal. The ST398 isolate 2L5 from animal in this study was closely related to the ST398 isolate HD15L3 from shopping trolley, which was also clustered with the animal ST398 MRSA isolate 08S00974 (CP020019) and human ST398 MRSA isolates NL1 (CP077741) and UMCG578 (CP077738). These results further suggest that exchange of MRSA isolates might occur among different niches, especially between shopping vehicles and humans.
4. Discussion
The contamination of animals and retail meats by S. aureus, especially MRSA isolates expressing virulence genes and resistance genes, remains an urgent public health issue [19]. The MRSA isolates from animals can spread to animal meats and are also a significant source of human MRSA by direct or indirect contact [44]. The prevalence of S. aureus in animals in this study (18.3%) was higher than that in animals (5.4%) from Xinjiang, China [45] and that (10.1%) in pigs in Chongqing, China [46], but significantly lower than that (65.4%) in other commercial farms from Shandong, China [47]. Unlike those previous studies [47,48,49,50], no samples from the two swine farms in this study carried ST398 MRSA, which might be due to the very low MRSA prevalence in the randomly selected farms in our study. Only one strain from one sample in this study might also inevitably miss some drug-resistant bacteria, including MRSA. S. aureus was identified in 14.6% of raw meat samples in our study, significantly lower than that (69.0%) in Danish retail meats [24] and that (33.6%) in Cambodian retail meat [51]. The prevalence of MRSA in retail meats in the current study was 2.2%, which was similar to that from Xinjiang (2.0%) China [45], but significantly lower than the detection rate of MRSA (13.1%) in retail meats in Denmark [24].
MRSA have also been detected in several public transportation systems such as public shared bicycles [25,26]. We presented that S. aureus isolates were widely distributed among shared shopping vehicles from supermarkets in Shandong, China. Of particular concern, we reported for the first time that MRSA was observed in shared shopping vehicles. Notably, eight of the eighteen supermarkets (44.4%) in this study possessed MRSA-positive shopping vehicle samples, which were distributed in all five cities studied, suggesting that MRSA isolates might have been prevalent in shopping vehicles in China. The overall high prevalence of MRSA (7.5%) in shopping vehicles in this study was higher than that in the Guangzhou metros in China (2.5%) [27], general public settings in London, UK (0.0%) [52] and shared bicycles in Chengdu, China (1.0%) [26], however, lower than that among public buses in the USA (62.5%) [53]. The high prevalence of MRSA in handrails of shopping vehicles in this study suggests that humans have contaminated the shopping vehicles, which will pose a threat to humans.
Virulence factors in S. aureus play an important role in pathogenesis [54]. A recent survey in China reported that only 3.9% MRSA strains harbored the tst-1 gene [55], and similarly, we did not find tst-1 in our study. Notably, the frequency of eta in S. aureus from human in Hainan, China was 57.3% [28] and 46.9% of the S. aureus in our study carried eta, much higher than those from human in central-southern China [56]. The Panton–Valentine leukocidin genes lukF-PV/lukS-PV has been proved to be highly pathogenic [57], and community-acquired lukF-PV/lukS-PV-associated necrotizing pneumonia infecting previously healthy people is often fatal [58]. The lukF-PV/lukS-PV-positive S. aureus isolates are more likely to cause skin and soft tissue abscesses and deep abscesses than lukF-PV/lukS-PV-negative isolates [58]. Among the MRSA isolates in this study, 15.0% carried the lukF-PV/lukS-PV gene and all lukF-PV/lukS-PV-positive MRSA isolates were from shared shopping vehicles, significantly higher than a previously study in which none of the MRSA isolates in metro system carried lukF-PV/lukS-PV [27]. However, the prevalence of lukF-PV/lukS-PV in S. aureus isolates in this study (6.1%) was lower than that in S. aureus isolates from humans in Hainan, China (47.6%) [28]. The enterotoxins are the main virulence factors that can trigger food poisoning and foodborne illness, causing fever and shock, even after the bacteria have been destroyed by heating [59]. Five classical enterotoxins, including sea, seb, sec, sed and see, have been considered to be associated with S. aureus food poisoning [60], and enterotoxin genes (sea, seb) were present in 45.0% of the S. aureus isolates from retail meats in this study, which will pose a threat to food safety. Of note, three MRSA isolates from retail meats in this study harbored the immune evasion cluster (IEC) system with the sea and sak genes, which is a specific marker of adaptation to the human host [61]. The prevalence of IEC in the 20 MRSA isolates (20.0%) was higher than that (2.8%) in MRSA isolates from retail meats in the Czech Republic [20]. In addition, MRSA isolate HD36L3 recovered from shopping basket had both the Panton–Valentine leucocidin toxin gene lukS-PV and seb, one of the five classical enterotoxins associated with food poisoning [60], suggesting that MRSA isolates in shared shopping vehicles will pose a threat to humans.
The resistance rate to ciprofloxacin and florfenicol among animal-sourced S. aureus isolates was significantly higher than that from raw meats and that from shopping vehicles, which might be related to the massive use of antibiotics in pig farms. MRSA isolates resistant to several antibiotics often result in the failure of treating infections. In this study, the MSSA isolates had higher resistance to ciprofloxacin, tetracycline and florfenicol than that in MRSA. This might be because the MSSA were mostly isolated from raw meats and animals, which were often treated with antibiotics, while MRSA were mostly from shared shopping vehicles in supermarkets. Most of the MSSA isolates might carry resistance genes, accounting for the resistances to antibiotics mentioned above, as shown in the WGS results, leading to the higher antibiotic resistances among MSSA isolates in this study. With WGS analysis, we found six resistance genes, conferring resistance to ≥3 classes of antibiotics including macrolides, in the five WGS-sequenced MRSA isolates from retail meats and shared shopping vehicles. The macrolide resistance genes erm(B) and erm(C) were not only prevalent in S. aureus isolates but also in MRSA isolates especially those from shared shopping vehicles. These results highlight the important role of shared shopping vehicles in maintaining a reservoir of antimicrobial resistance. Antimicrobial-resistant bacteria and resistance genes are constantly enriched in the environment and spread through the environment to humans via horizontal transfer [62,63].
The three MRSA isolates from retail meats in this study were ST6, a common ST type of S. aureus from food-poisoning outbreaks [64,65] and shared bicycles [26] in China. ST59, a common ST type of MRSA clone in both humans [66] and shared bicycles [26] in China, was also found in shared shopping baskets in this study. ST398-MSSA was a prevalent ST type in humans in China [66] and ST398-MSSA carrying 14~15 resistance genes was also present in animal and shopping trolley in the current study. Contact with animals infected by ST398 S. aureus has been considered one of the major risk factors for human infection. All MRSA isolates from retail meats and shopping vehicles in this study belonged to the SCCmec type IVa, which was also the most prevalent MRSA clone in hospitals in Hainan, China [28], indicating a correlation between MRSA isolates from retail meats, environment and humans. Our results suggest that both retail meats and shared shopping vehicles are important reservoirs of pathogens.
Based on WGS analysis, the MRSA isolates from retail meats were clustered closely with MRSA isolates from animals and humans in China, suggesting that the MRSA isolates in food might be derived from animals and could be transmitted to humans via the food chain. The ST398 isolate from animal in this study was closely related to the previously reported animal ST398 MRSA isolates and human ST398 MRSA isolates, supporting the possibility of ST398 MRSA transmission between animals and humans. The animal and human ST398 MRSA isolates were also clustered with the ST398 isolate from shopping trolley in this study, indicating that shared shopping vehicles are likely to play an important role in bacterial transmission. Importantly, the MRSA isolates from shopping vehicles in this study were clustered with MRSA isolates from human blood and animals. These results confirm that exchange of MRSA isolates occurred among different niches. Shopping vehicles might be contaminated from human, either from human infection or humans touching contaminated food/animals, and shopping vehicles in supermarkets may participate in transmitting MRSA isolates among humans. Combination of hand hygiene and disinfection of shopping vehicles handrails before and after use, is recommended to prevent MRSA transmission.
We acknowledge that this study has some limitations. First, the sample size was geographically limited and may not reflect the prevalence of MRSA especially in animals and retail meats from Shandong, China. Second, only a small number of isolates were selected for WGS in this study and the genomes of the remaining MRSA have not yet been analyzed, which might limit the phylogenetic inferences. The genomic information on a wide range of isolates will provide a more detailed genotypic characteristics of MRSA. Additionally, the corresponding humans have not been sampled because the humans consuming the retail meats or touching the shopping vehicles in our study are difficult to trace. Future research should include broader geographic sampling and more genome data to jointly decipher the molecular exchange mechanisms of MRSA isolates among different niches.
5. Conclusions
In summary, S. aureus isolates are widely distributed in animals, retail meats and shared shopping vehicles in Shandong province of China. We report for the first time a high prevalence of MRSA (7.5%) in shared shopping vehicles, followed by retail meats (2.2%), suggesting that both retail meats and shared shopping vehicles serve as an important reservoir of MRSA. The MRSA isolates from retail meats/shared shopping vehicles in this study were clustered with previously reported MRSA isolates from animals and humans in China, confirming that exchange of MRSA isolates occurred among different niches, which further revealed that the MRSA isolates in food and shared shopping vehicles might pose a threat to humans. Nationwide epidemiology studies about MRSA are needed to fully assess the risk of retail meats and shared shopping vehicles in spreading diseases. To our knowledge, we first reported the widespread of MRSA in shared shopping trolleys and baskets in China.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aslam B. Khurshid M. Arshad M.I. Muzammil S. Rasool M. Yasmeen N. Shah T. Chaudhry T.H. Rasool M.H. Shahid A. Antibiotic Resistance: One Health One World Outlook Front. Cell. Infect. Microbiol.20211177151010.3389/fcimb.2021.77151034900756 PMC 8656695 · doi ↗ · pubmed ↗
- 2Liu K. Wang M. Zhang Y. Fang C. Zhang R. Fang L. Sun J. Liu Y. Liao X. Distribution of antibiotic resistance genes and their pathogen hosts in duck farm environments in south-east coastal China Appl. Microbiol. Biotechnol.202410813610.1007/s 00253-023-12842-438229327 PMC 10789667 · doi ↗ · pubmed ↗
- 3Overgaauw P.A.M. Vinke C.M. Hagen M. Lipman L.J.A. A One Health Perspective on the Human-Companion Animal Relationship with Emphasis on Zoonotic Aspects Int. J. Environ. Res. Public Health 202017378910.3390/ijerph 1711378932471058 PMC 7312520 · doi ↗ · pubmed ↗
- 4Kourtis A.P. Hatfield K. Baggs J. Mu Y. See I. Epson E. Nadle J. Kainer M.A. Dumyati G. Petit S. Vital Signs: Epidemiology and Recent Trends in Methicillin-Resistant and in Methicillin-Susceptible Staphylococcus aureus Bloodstream Infections—United States MMWR Morb. Mortal. Wkly. Rep.20196821421910.15585/mmwr.mm 6809 e 130845118 PMC 6421967 · doi ↗ · pubmed ↗
- 5Bartsch S.M. Asti L. Nyathi S. Spiker M.L. Lee B.Y. Estimated Cost to a Restaurant of a Foodborne Illness Outbreak Public Health Rep.201813327428610.1177/003335491775112929656701 PMC 5958383 · doi ↗ · pubmed ↗
- 6Lu D. Liu J. Liu H. Guo Y. Dai Y. Liang J. Chen L. Xu L. Fu P. Li N. Epidemiological Features of Foodborne Disease Outbreaks in Catering Service Facilities—China, 2010–2020 China CDC Wkly.2023547948410.46234/ccdcw 2023.09137408617 PMC 10318556 · doi ↗ · pubmed ↗
- 7Gajdacs M. The Continuing Threat of Methicillin-Resistant Staphylococcus aureus Antibiotics 201985210.3390/antibiotics 802005231052511 PMC 6627156 · doi ↗ · pubmed ↗
- 8Hu F. Wang M. Zhu D. Wang F. CHINET efforts to control antimicrobial resistance in China J. Glob. Antimicrob. Resist.202021767710.1016/j.jgar.2020.03.00732205265 · doi ↗ · pubmed ↗