Extracellular Matrix in Human Disease and Therapy: From Pathogenic Remodeling to Biomaterial Platforms and Precision Diagnostics
Jun-Hyeog Jang

TL;DR
This paper reviews how changes in the extracellular matrix drive chronic diseases and explores new therapies and diagnostics targeting the matrix.
Contribution
The paper provides a comprehensive review linking ECM biology to disease mechanisms and translational strategies for therapy and diagnostics.
Findings
Dysregulated ECM remodeling is an upstream driver of chronic diseases like fibrosis, cancer, and metabolic disorders.
ECM-based biomaterials and precision diagnostics offer new translational opportunities for disease treatment.
Targeting ECM-cell signaling pathways could enhance anti-fibrotic and immunotherapeutic strategies.
Abstract
The extracellular matrix (ECM) is a dynamic, tissue-specific network that integrates biochemical and mechanical cues to regulate cell behavior and organ homeostasis. Increasing evidence indicates that dysregulated ECM remodeling is an upstream driver of chronic human diseases rather than a passive consequence of injury. This review summarizes principles of ECM organization, mechanotransduction, and pathological remodeling and highlights translational opportunities for ECM-targeted therapies, biomaterial platforms, and precision diagnostics. We conducted a narrative synthesis of foundational and recent literature covering ECM composition and turnover, stiffness-dependent signaling, and disease-associated remodeling across fibrosis/cardiovascular disease, cancer, and metabolic disorders, together with advances in ECM-based biomaterials, drug delivery, and ECMderived biomarkers and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
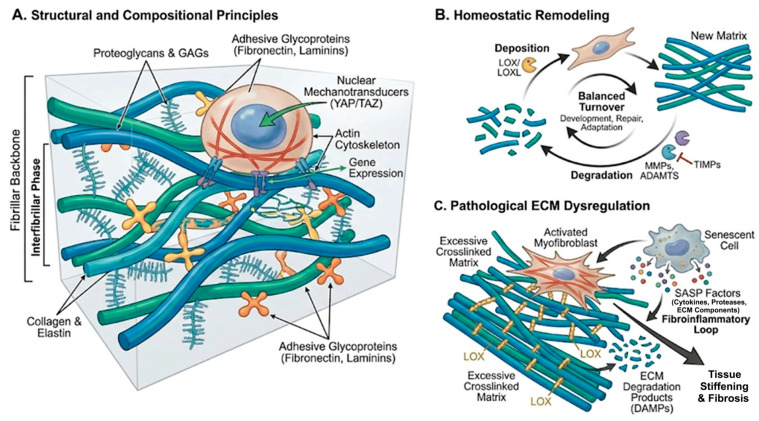 Figure 1
Figure 1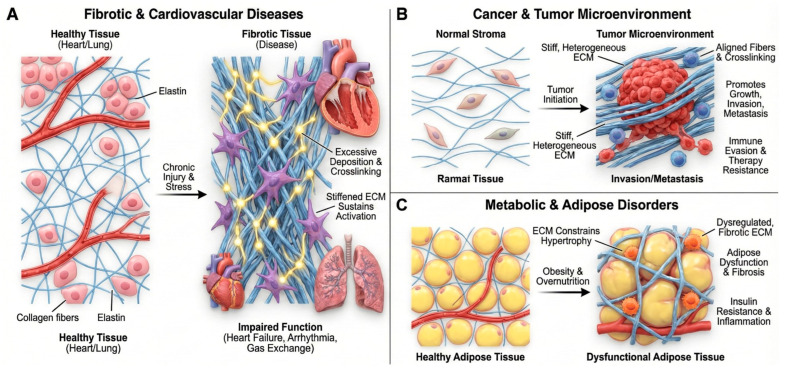 Figure 2
Figure 2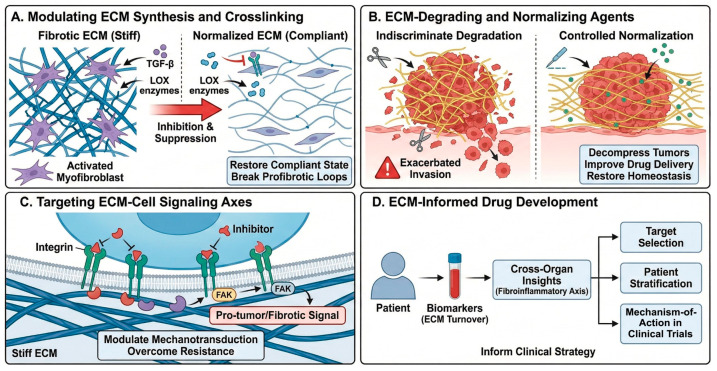 Figure 3
Figure 3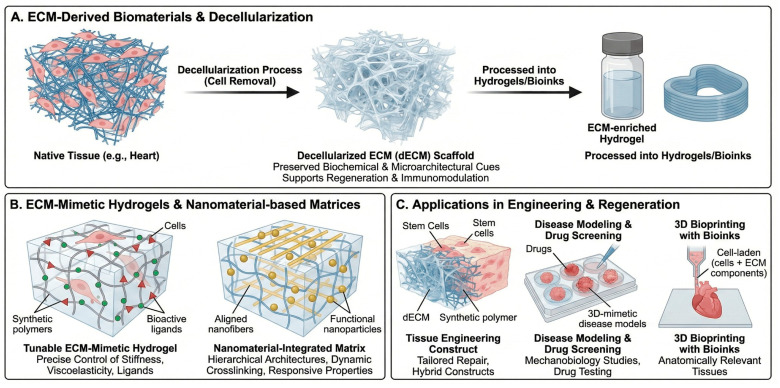 Figure 4
Figure 4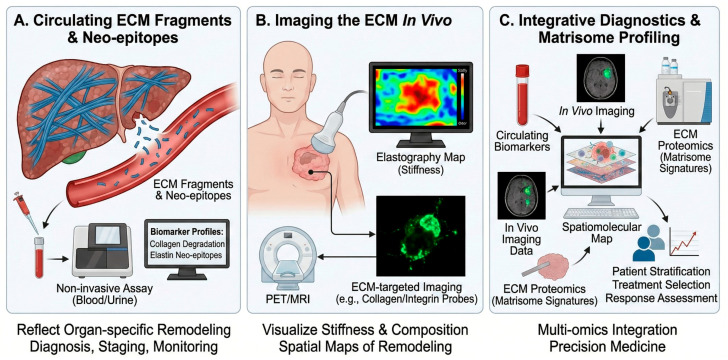 Figure 5
Figure 5 Figure 6
Figure 6- —INHA UNIVERSITY Research Grant (2025)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCell Adhesion Molecules Research · Cellular Mechanics and Interactions · Tissue Engineering and Regenerative Medicine
1. Introduction
The extracellular matrix (ECM) is a three-dimensional macromolecular network composed of collagens, elastin, proteoglycans, and glycoproteins that provides structural integrity and instructive biochemical and biophysical cues to resident cells. Far from being an inert scaffold, the ECM is a dynamic and finely regulated component of the tissue microenvironment that controls cell adhesion, migration, proliferation, differentiation, and survival through a combination of biochemical signaling and mechanotransduction pathways [1,2,3]. In this review, we frame ECM remodeling as a shared, quantifiable mechanobiological axis across major chronic diseases and highlight ECM normalization as a unifying translational principle that connects mechanism-based therapeutics, ECM-informed biomaterials, and precision diagnostics.
In healthy tissues, ECM synthesis, crosslinking, and degradation are tightly balanced by matrix-producing cells and matrix-remodeling enzymes, maintaining tissue-specific architecture and mechanical properties [2,4]. In chronic disease, this equilibrium is disrupted, leading to pathological ECM stiffening, fibrosis, calcification, or excessive degradation that actively drives disease progression rather than merely reflecting downstream damage [4,5,6,7,8,9,10,11].
Recent comprehensive reviews highlight ECM dysregulation as a central hub linking aging, chronic inflammation, fibrosis, calcification, and tumorigenesis [2,4,12]. For example, sustained fibroblast activation and maladaptive matrix deposition support progressive organ fibrosis [5,12], while matrix stiffening and altered topography promote myofibroblast persistence, endothelial dysfunction, and immune cell skewing [4,6,10,11,13,14,15].
Concurrently, the ECM is emerging as an attractive therapeutic and diagnostic axis in translational biomedicine. ECM-derived cues can be harnessed to engineer tissue-specific biomaterials and decellularized scaffolds for regeneration [16,17,18,19,20], while ECM fragments and neo-epitopes serve as minimally invasive biomarkers reflecting dynamic tissue re-modeling [4,21]. Advances in ECM-mimetic hydrogels, nanomaterial-based matrices, and matrisome-informed diagnostics align closely with the aims of translational biomedicine in developing novel biopharmaceutical products, targeted therapies, and mechanistically grounded biomarkers [8,22,23,24,25,26,27,28].
This review provides a disease-focused overview of ECM biology and its translational exploitation: (i) fundamental principles of ECM organization and remodeling in health and disease; (ii) ECM as an active driver in key human pathologies; (iii) ECM-targeted therapeutic strategies; (iv) ECM-based biomaterials and decellularized scaffolds; and (v) ECM-derived biomarkers, imaging, and integrative diagnostics with relevance to personalized medicine [1,2,8,21,23,26,27]. Compared with prior reviews that focus on single disease settings or individual translational modalities, we frame ECM remodeling as a shared, quantifiable mechanobiological axis across chronic diseases and highlight “ECM normalization” as a unifying translational principle linking therapeutics, biomaterials, and precision diagnostics (Scheme 1) [2,4,11,22,29,30,31,32,33].
2. ECM Organization and Remodeling in Health and Disease
2.1. Structural and Compositional Principles
The ECM consists of a fibrillar backbone (primarily fibrillar collagens and elastin), an interfibrillar phase of proteoglycans and glycosaminoglycans, and adhesive glycoproteins such as fibronectin and laminins (Figure 1A) [1,2]. These components assemble into tissue-specific architectures that define porosity, viscoelasticity, and anisotropy, thereby shaping cell–matrix interactions. Cells sense the ECM via integrins and other receptors, forming focal adhesions that couple to the actin cytoskeleton and to nuclear mechanotransducers such as YAP/TAZ. This mechanochemical coupling enables cells to translate matrix stiffness, topography, and strain into gene-expression programs and lineage decisions [2,6,13,34,35].
2.2. Homeostatic Remodeling
Under physiological conditions, the ECM is continuously remodeled by matrix metalloproteinases (MMPs), ADAMTS proteases, and crosslinking enzymes such as lysyl oxidases (LOX/LOXL), balanced by tissue inhibitors of metalloproteinases (TIMPs) (Figure 1B) [2,7,8,9,12,36]. Controlled deposition and degradation facilitate development, wound repair, and adaptation to mechanical load.
2.3. Pathological ECM Dysregulation
Chronic injury, metabolic stress, and aging induce persistent activation of matrix-producing cells (e.g., fibroblasts, myofibroblasts, vascular smooth muscle cells), skewing the balance towards excessive crosslinked matrix deposition, altered composition, and accumulation of bioactive matrix fragments (Figure 1C) [4,5,12]. Senescent cells with senescence-associated secretory phenotypes (SASP) secrete pro-fibrotic cytokines, proteases, and ECM components that perpetuate a fibroinflammatory loop [4]. Systemic accumulation of ECM degradation products further amplifies inflammation and can act as damage-associated molecular patterns (DAMPs) [4]. Nyström and colleagues emphasized that aging, ECM remodeling, and fibrosis are tightly intertwined, with age-related ECM stiffening predisposing tissues to chronic fibrotic remodeling and impaired resolution [13].
Beyond fibrosis, aging itself is accompanied by progressive ECM “hardening” and compositional drift, driven by non-enzymatic glycation/advanced glycation end-product (AGE) crosslinks, elastin fragmentation, impaired proteostasis, and a pro-inflammatory senescence-associated secretory phenotype that remodels the matrisome and raises basal mechanotransduction tone. Emerging interventional studies suggest that targeting the aging ECM may be feasible: senolytic therapy has shown first-in-human signals in idiopathic pulmonary fibrosis, consistent with reducing senescence-linked profibrotic ECM remodeling [37], and AGE crosslink–modifying approaches (e.g., alagebrium/ALT-711) have been evaluated clinically to improve vascular and myocardial compliance [38].
A self-sustaining cycle of ECM stiffening and mechanotransduction drives this pathological state, where increased stiffness promotes further fibroblast activation and matrix deposition. These concepts are reflected across organ systems, including lung, liver, kidney, cardiovascular system, and skin [3,4,5,6,7,8,9,10,11,12,13,14,15].
3. ECM as an Active Driver in Major Human Diseases
ECM remodeling is an active driver of pathology by reshaping tissue mechanics and cell signaling across organs. Table 1 summarizes representative ECM alterations and their translational implications across major disease contexts. The following subsections outline key disease settings in which these mechanisms are most evident.
3.1. Fibrotic and Cardiovascular Diseases
Fibrosis is characterized by excessive ECM deposition, altered crosslinking, and loss of normal matrix architecture, resulting in increased tissue stiffness and impaired function (Figure 2A). Mechanistic studies support a model in which stiffened ECM sustains myofibroblast activation and perpetuates fibrogenic signaling, thereby acting as a driver of progressive fibrosis [5,12]. In cardiac disease, pressure overload, ischemia, and metabolic stress trigger fibroblast activation and collagen-rich scar formation. ECM remodeling alters ventricular compliance, electrical conduction, and mechanosensing, contributing to heart failure and arrhythmia [14]. Reviews of cardiac fibrosis highlight ECM as both a mediator and a potential therapeutic target, including strategies to modulate collagen crosslinking, TGF-β signaling, and matricellular proteins [14]. Similarly, in pulmonary fibrosis, aberrant ECM deposition and stiffened foci create a profibrotic microenvironment that amplifies fibroblast activation, distorts alveolar architecture, and compromises gas exchange [5,6,7,8,9,10,11,12,15].
3.2. Cancer and the Tumor Microenvironment
The tumor-associated ECM is structurally and compositionally distinct from normal stroma (Figure 2B). Increased collagen density, aligned fibers, enhanced crosslinking, and deposition of matricellular proteins collectively create a stiff, heterogeneous environment that promotes tumor growth, invasion, and immune evasion [29,30,31,32]. Sleeboom et al. proposed the ECM as a “hallmark” of cancer and metastasis, integrating mechanical, biochemical, and topological cues that govern cancer-cell plasticity, migration routes, and access to vasculature [29]. ECM remodeling enzymes and crosslinking pathways (e.g., LOX/LOXL, MMPs) shape metastatic niches and influence response to chemotherapy and immunotherapy [29,30,31]. The interplay between ECM and therapy is bidirectional: cytotoxic and targeted therapies remodel ECM composition and stiffness, while pre-existing ECM states regulate drug penetration, mechanosensitive signaling (e.g., FAK, YAP/TAZ), and immune-cell infiltration [30,32]. Biophysical analyses highlight that ECM stiffness and architecture modulate T cell and NK cell migration, checkpoint expression, and susceptibility to immunotherapies [6,7,8,9,32,34,36,39,44].
3.3. Metabolic, Endocrine, and Adipose Tissue Disorders
Adipose tissue expansion in obesity is accompanied by ECM remodeling that constrains adipocyte hypertrophy, impairs vascularization, and sustains low-grade inflammation (Figure 2C). A recent review on adipose tissue development and ECM emphasizes that dysregulated matrix deposition and crosslinking are key determinants of adipose dysfunction, insulin resistance, and fibrosis in obesity and metabolic syndrome [7,8,9,10,11,40]. These findings position ECM as an upstream regulator of endocrine and metabolic homeostasis, linking biomechanical and immunometabolic pathways relevant to cardiovascular and hepatic complications of obesity.
Across fibrosis, cancer, and metabolic disease, recurrent ECM programs—stiffening/crosslinking, aligned fibrillar architecture, and protease-driven remodeling—converge on shared mechanotransduction axes (integrin–FAK and YAP/TAZ) that couple matrix state to inflammatory and pro-survival signaling. However, the strength of evidence and the translational levers differ by context: in fibrosis, feed-forward stiffening–myofibroblast loops are strongly supported by mechanistic and in vivo studies, whereas in cancer, “normalization” strategies must balance improved delivery against context-dependent risks of invasion, underscoring the need for quantitative ECM biomarkers and imaging readouts to guide clinical deployment [3,4,5,6,12,29,30,31,32,34,39,44].
4. The ECM as a Therapeutic Target
Given its central role in disease initiation and progression, the ECM is increasingly targeted by pharmacological, biological, and device-based interventions [4,22].
4.1. Modulating ECM Synthesis and Crosslinking
Anti-fibrotic strategies include inhibition of TGF-β signaling, suppression of myofibroblast activation, and targeting of crosslinking enzymes such as LOX/LOXL to reduce excessive collagen crosslinking and stiffness (Figure 3A) [4,12]. These approaches aim to restore more compliant matrix states and break profibrotic feedback loops. Boaru et al. highlighted potential therapeutic avenues targeting senescence-associated ECM remodeling and calcification, such as senolytics, anti-inflammatory agents, and regulators of mineralization [4,6,7,8,9,15].
4.2. ECM-Degrading and ECM-Normalizing Agents
ECM-degrading agents (e.g., specific hyaluronidases, collagenases) can decompress solid tumors, improve perfusion, and enhance delivery of chemotherapy and immunotherapy when applied in a controlled manner (Figure 3B) [29,30]. However, indiscriminate degradation may exacerbate invasion or metastasis; thus, “ECM normalization” rather than “ECM ablation” is now favored, aiming to restore a more homeostatic matrix architec-ture [29,30,31]. Importantly, while stromal decompression and controlled ECM normalization show strong preclinical rationale, clinical translation remains context-dependent, and indiscriminate ECM degradation can exacerbate invasion or compromise essential repair processes; therefore, dose, timing, and patient selection should be guided by quantitative ECM readouts (turnover biomarkers and ECM-targeted imaging).
4.3. Targeting ECM–Cell Signaling Axes
Inhibitors of integrins and focal adhesion kinase (FAK) are under investigation as mechanotransduction modulators in cancer and fibrotic disease (Figure 3C). A recent study by Betriu et al. demonstrated that increased stiffness in a 3D peptide scaffold model of pancreatic ductal adenocarcinoma (PDAC) downregulated FAK expression and altered mechanosignaling, underscoring matrix–FAK feedback in tumor biology [45]. Combination regimens that pair ECM-modulating agents with immune checkpoint blockade, anti-angiogenic therapies, or targeted kinase inhibitors are being explored to overcome ECM-mediated resistance [3,6,22,30,31,32,39].
4.4. ECM-Informed Drug Development
Karsdal et al. [21] synthesized cross-organ insights into ECM-associated diagnostics and therapeutics, proposing a “fibroinflammatory axis” as a common denominator across >50 chronic diseases (Figure 3D) [22]. Their framework emphasizes that understanding ECM turnover and cross-tissue signatures can inform target selection, patient stratification, and mechanism-of-action biomarkers in early-phase clinical trials.
4.5. Clinical Translation and Emerging Trials
Although many ECM-targeting strategies remain preclinical, several have advanced to clinical evaluation. In idiopathic pulmonary fibrosis, the LOXL2-blocking antibody simtuzumab was tested in a randomized phase 2 trial, illustrating both the feasibility of targeting matrix crosslinking and the importance of patient stratification and pharmacodynamic biomarkers for ECM engagement [46]. In metastatic pancreatic ductal adenocarcinoma, enzymatic hyaluronan depletion with pegvorhyaluronidase alfa (PEGPH20) was evaluated in the phase 3 HALO 109-301 trial to improve perfusion and chemotherapy delivery—an archetype of controlled matrix decompression/normalization rather than indiscriminate ECM ablation [47]. Mechanotransduction inhibitors have also entered clinical testing; for example, the focal adhesion kinase (FAK) inhibitor defactinib has been assessed in randomized phase 2 studies as maintenance therapy in malignant pleural mesothelioma [48]. Table 2 summarizes representative clinical-stage ECM-modulating approaches and their translational rationale.
5. ECM-Based Biomaterials and Decellularized Scaffolds
Biomaterial science has increasingly turned to the ECM as both a blueprint and a raw material for developing regenerative scaffolds and advanced therapeutic products [16,17,18,19,20,23,33,43].
5.1. ECM-Derived Biomaterials
ECM-derived materials, including decellularized tissues and ECM-enriched hydrogels, preserve many native biochemical motifs and microarchitectures that support cell adhesion, migration, and lineage specification (Figure 4A) [16,17,18]. Hussey et al. outlined how ECM-based materials provide instructive cues for regenerative medicine, highlighting their capacity to promote constructive remodeling and immunomodulation [17]. Noro and Vilaça-Faria recently reviewed ECM-derived materials for tissue engineering and regenerative medicine, describing approaches to decellularization, recellularization, and engineering of ECM-based scaffolds tailored to specific tissues [16]. Long et al. emphasized that decellularized ECM (dECM) scaffolds can bridge regeneration and inflammation, depending on how they are prepared and integrated into host tissues [18]. Jin and co-workers further discussed acellular ECM scaffolds as versatile platforms for organ repair, including heart, liver, and musculoskeletal tissues, providing structural templates and bioactive signals in preclinical models [8,19,26,27].
5.2. ECM Scaffolds in Tissue Engineering and Regeneration
An in-depth review by Mangani et al. systematically addressed the design and applications of ECM scaffolds in tissue engineering and regeneration, including strategies to tailor mechanical properties, matrix composition, and degradation kinetics to guide tissue-specific repair (Figure 4C) [20]. These ECM scaffolds can be used alone or in combination with synthetic polymers and bioactive molecules to engineer hybrid constructs with improved mechanical robustness and biological performance [17,20].
Perspective and limitations. Clinically, ECM-based biomaterials are attractive because they present native biochemical motifs and can promote constructive remodeling, but translation is constrained by batch-to-batch variability, incomplete decellularization (residual DNA/lipids), donor- and tissue-source heterogeneity, sterilization-induced loss of bioactivity, limited mechanical robustness for load-bearing sites, and regulatory requirements for reproducible composition and safety. Hybrid designs that combine dECM with defined synthetic polymers and standardized manufacturing/quality-control pipelines may help address scalability and consistency while retaining ECM instructiveness [16,17,18,19,20,33].
5.3. ECM-Mimetic Hydrogels and Nanomaterial-Based Matrices
ECM-mimetic hydrogels aim to recapitulate key structural and biochemical features of native matrix while allowing precise control over stiffness, viscoelasticity, porosity, and ligand presentation (Figure 4B). Nicolas et al. described fundamental concepts and applications of 3D ECM mimics for mechanobiology and drug screening, emphasizing how tunable hydrogels capture disease-relevant mechanical states [33]. Kim and Cha reviewed extracellular-matrix-mimetic hydrogels based on nanomaterials, highlighting how nanoparticles, nanofibers, and supramolecular assemblies can be integrated into hydrogels to provide hierarchical architectures, dynamic crosslinking, and responsive properties [22]. These systems are particularly relevant for controlled delivery of growth factors, small molecules, and cells, as well as for engineering disease models for drug testing. Wang et al. focused on ECM-based bioinks for 3D bioprinting, summarizing how native and decellularized ECM components can be formulated into printable inks to construct anatomically relevant, cell-laden tissues with appropriate mechanical and biochemical characteristics [6,7,9,26,27,41,42].
6. ECM in Nano, Cell, and Gene Therapies
ECM considerations are increasingly embedded into the design of biologics, cell therapies, and nanomedicines. ECM-mimetic hydrogels and scaffolds serve as delivery platforms for stem and progenitor cells, shielding them from hostile microenvironments while providing instructive cues that promote engraftment and functional integration [17,20,33,43]. For example, ECM-mimetic hydrogels based on nanomaterials can be engineered to release chemokines and growth factors in response to local enzymatic or mechanical cues, thereby enhancing tissue-specific regeneration [22]. Nanoparticles and antibody–drug conjugates can be functionalized with ECM-binding peptides or antibodies to target fibrotic or tumor-associated ECM components, improving local drug accumulation while reducing off-target toxicity [6,8,22,29,30,31,32]. ECM remodeling enzymes and ECM-bound growth factors also influence viral vector diffusion, transduction efficiency, and off-target distribution in gene therapy. Rationally modifying ECM composition or stiffness may thus improve vector delivery and safety profiles in cardiac, hepatic, or neuromuscular gene therapies, although this area remains relatively underexplored compared with regenerative scaffolds and nanomedicines.
7. ECM-Derived Biomarkers, Imaging, and Integrative Diagnostics
7.1. Circulating ECM Fragments and Neo-Epitopes
ECM turnover generates fragments and neo-epitopes that can be quantified in blood, urine, or other biofluids (Figure 5A). These biomarkers reflect organ-specific matrix remodeling and can serve as non-invasive tools for diagnosis, staging, and monitoring of chronic diseases [4,22]. Karsdal et al. [21] summarized the development of ECM-associated diagnostics, including assays for collagen formation and degradation fragments, elastin and laminin neo-epitopes, and composite biomarker panels that capture fibroinflammatory activity across organs [22]. Such assays are being applied in liver, lung, kidney, cardiovascular, and musculoskeletal diseases, as well as in oncology, to stratify patients and monitor treatment response. Boaru et al. highlighted how ECM dysregulation in aging and calcification produces characteristic fragment signatures that could be exploited as mechanistically grounded biomarkers [4,8].
7.2. Imaging the ECM In Vivo
Advances in molecular imaging enable visualization of ECM components, crosslinking, and stiffness in vivo using MRI, PET, SPECT, and ultrasound-based elastography (Figure 5B). ECM-targeted contrast agents (e.g., collagen-specific MRI probes, integrin-targeted PET tracers) provide spatial maps of matrix remodeling in tumors and fibrotic organs, complementing circulating biomarkers [22,23]. Vedarethinam and colleagues proposed an “integrative diagnostics” paradigm in which ECM imaging is combined with mass spectrometry-based matrisome profiling and multi-omics analyses to build spatiomolecular maps of cancer ECM remodeling (Figure 5C) [6,7,8,9,23,24,25,28]. This approach supports early detection, patient stratification, and response assessment in precision oncology.
Mechanistically, ECM probes achieve specificity either by binding matrix macromolecules directly (e.g., collagen- or elastin-binding peptides/antibodies) or by targeting cell-surface receptors whose expression reflects ECM remodeling (e.g., integrins). Collagen-targeted PET probes such as the collagen-binding peptide tracer ^68^Ga-CBP8 bind exposed collagen in fibrotic lesions, enabling quantitative mapping of active collagen deposition and serving as a pharmacodynamic readout in anti-fibrotic therapy studies [49,50]. Similarly, collagen-specific molecular MRI probes (e.g., the gadolinium-based peptide probe EP-3533) accumulate in collagen-rich fibrotic tissue to report spatial heterogeneity of matrix remodeling with high anatomical resolution [51]. For the ECM–cell interface, RGD-based PET tracers (e.g., ^18^F-galacto-RGD) bind activated integrins such as αvβ3 on angiogenic endothelium and tumor cells, providing a receptor-level surrogate of remodeling and invasive potential (Table 3) [52]. Together with stiffness mapping by elastography, these probes support precise, targetable in vivo ECM phenotyping (Figure 5B) and can be integrated with circulating biomarkers and matrisome proteomics for spatiomolecular stratification [22,23].
7.3. ECM Proteomics and Immuno-Oncology
Day et al. reviewed ECM-focused proteomics in immuno-oncology, emphasizing that matrisome signatures in tumors correlate with immune infiltration, immunosuppressive niches, and response to checkpoint blockade [24,25,28,43]. By resolving ECM composition and post-translational modifications, proteomics can identify targetable ECM components and generate candidate biomarkers for immunotherapy response.
8. Conclusions and Future Directions
Viewed as a cross-disease mechanobiological axis, ECM remodeling provides measurable targets for intervention and a rationale for ECM normalization strategies that can be paired with biomaterials and diagnostic readouts to enable precision medicine. The ECM has emerged from the background of histological sections to become a central, actionable axis in human disease and therapy. Foundational and recent work converge on several key themes [1,2,53,54]:
- ECM as an active disease driver. Pathological matrix remodeling, driven by chronic inflammation, senescence, and dysregulated mechanotransduction, underpins fibrosis, calcification, and tumor progression across organs [4,5,12,13,14,29]. Targeting ECM biology is therefore essential for modifying disease trajectories, not merely managing late complications [1,2,3,5,8,10,11,12].
- Convergence of ECM biology and biomaterials science. ECM-derived scaffolds, decellularized matrices, and ECM-mimetic hydrogels now constitute a robust toolbox for tissue engineering, organ repair, and in vitro disease modeling [16,17,18,19,20,33,41]. Their modularity enables integration of cells, biologics, and nanomedicines into clinically relevant constructs tailored to specific tissues and indications [1,2,16,17,26,27].
- ECM-informed precision diagnostics. Circulating ECM fragments, ECM-targeted imaging probes, and matrisome-centered proteomics are redefining how clinicians can detect and monitor tissue remodeling [4,21,22,23,43]. Integrative frameworks that combine these modalities with genomics and transcriptomics are particularly promising for precision oncology and chronic fibrotic diseases [8,23,24,25,28].
- ECM-aware therapeutic development. Incorporating ECM metrics into early-phase clinical trials—whether as enrichment biomarkers, pharmacodynamic readouts, or surrogate endpoints—can de-risk drug development and support mechanism-based patient stratification, in line with translational aims of the field [22].
- AI-enabled ECM analytics and design. Rapid advances in computer vision and machine learning enable automated quantification of collagen architecture and ECM signatures from histology and label-free microscopy (e.g., SHG), integration of matrisome-scale proteomics with clinical phenotypes for biomarker discovery, and data-driven optimization of ECM-mimicking hydrogels and bioinks for bioprinting and regenerative applications [55,56].
Key open questions remain, including whether ECM remodeling is primarily an initiating driver or a self-reinforcing consequence of chronic pathology in different organs; resolving this will require causal, longitudinal studies integrating spatial multi-omics with quantitative mechanics and validated clinical biomarkers. Future priorities include: (i) mapping tissue- and disease-specific “mechanobiological codebooks” that link ECM mechanics and composition to cellular states; (ii) integrating ECM metrics with multi-omics at single-cell and spatial resolution; (iii) designing ECM-normalizing therapies that restore homeostatic architecture without compromising necessary repair; and (iv) standardizing ECM biomarkers and imaging protocols for regulatory-grade clinical use. By bridging basic matrix biology with biomaterial engineering, nanotechnology, and clinical research, the ECM field is well positioned to deliver the next generation of biomedicines—ranging from ECM-guided cell therapies and smart hydrogels to matrisome-informed diagnostics and ECM-targeted therapeutics. This multifaceted translational potential makes the ECM a particularly compelling focus for future work in this area [1,2,16,17,24,25,28].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hynes R.O. The extracellular matrix: Not just pretty fibrils Science 20093261216121910.1126/science.117600919965464 PMC 3536535 · doi ↗ · pubmed ↗
- 2Kyriakopoulou K. Piperigkou Z. Tzaferi K. Karamanos N.K. Trends in extracellular matrix biology Mol. Biol. Rep.20235085386310.1007/s 11033-022-07931-y 36342580 PMC 9884264 · doi ↗ · pubmed ↗
- 3Humphrey J.D. Dufresne E.R. Schwartz M.A. Mechanotransduction and extracellular matrix homeostasis Nat. Rev. Mol. Cell Biol.20141580281210.1038/nrm 389625355505 PMC 4513363 · doi ↗ · pubmed ↗
- 4Boaru D.L. De Leon-Oliva D. De Castro-Martinez P. Garcia-Montero C. Fraile-Martinez O. García-González B. Pérez-González I. Alhaddadin M. Barrena-Blázquez S. Lopez-Gonzalez L. Extracellular matrix dysregulation in aging, calcification, and cancer diseases: Insights into cellular senescence, inflammation, and novel therapeutic strategies Int. J. Biol. Sci.2025216808688110.7150/ijbs.11930141281748 PMC 12631191 · doi ↗ · pubmed ↗
- 5Herrera J. Henke C.A. Bitterman P.B. Extracellular matrix as a driver of progressive fibrosis J. Clin. Investig.2018128455310.1172/JCI 9355729293088 PMC 5749528 · doi ↗ · pubmed ↗
- 6Discher D.E. Janmey P. Wang Y.-L. Tissue cells feel and respond to the stiffness of their substrate Science 20053101139114310.1126/science.111699516293750 · doi ↗ · pubmed ↗
- 7Levental K.R. Yu H. Kass L. Lakins J.N. Egeblad M. Erler J.T. Fong S.F.T. Csiszar K. Giaccia A. Weninger W. Matrix crosslinking forces tumor progression by enhancing integrin signaling Cell 200913989190610.1016/j.cell.2009.10.02719931152 PMC 2788004 · doi ↗ · pubmed ↗
- 8Bonnans C. Chou J. Werb Z. Remodelling the extracellular matrix in development and disease Nat. Rev. Mol. Cell Biol.20141578680110.1038/nrm 390425415508 PMC 4316204 · doi ↗ · pubmed ↗