Identification and Functional Analysis of tgfb2b Gene in Ovarian Development of Chinese Tongue Sole (Cynoglossus semilaevis)
Xihong Li, Kaili Zhang, Yue Zhang, Zhijie Li, Zhangfan Chen, Hongyan Wang, Songlin Chen, Na Wang

TL;DR
This study identifies the tgfb2b gene's role in ovarian development in Chinese tongue sole fish and explores its regulation and interactions with other genes and miRNA.
Contribution
The study reveals the functional role of tgfb2b in ovarian development and its regulation by C/EBPα, c-Jun, and a specific miRNA in Chinese tongue sole.
Findings
tgfb2b expression increases during ovarian development and is localized in oocytes but not in testis.
C/EBPα and c-Jun repress tgfb2b promoter activity, while its knockdown reduces downstream and sex-related gene expression.
A miRNA (novel-m0083-3p) inversely regulates tgfb2b and directly targets it, as confirmed by dual-luciferase assays.
Abstract
Transforming growth factor β (TGF-β) superfamily members are critical in teleost sex determination and differentiation. Tgfb2b is an important TGF-β ligand gene exhibiting dominant expression in the ovary of Chinese tongue sole (Cynoglossus semilaevis), yet its function in sex regulation remains unclear. In the present study, the gene expression pattern, transcriptional regulation, and knockdown effect were examined. Its expression persisted and showed a gradual increase throughout ovarian development from 3 months to 1.5 years post-hatching. In situ hybridization (ISH) revealed that the gene was distributed across oocytes at stages I–III, while scarcely detectable in the testis. The transcriptional factors CCAAT/enhancer binding protein α (C/EBPα) and Jun proto-oncogene AP-1 transcription factor subunit (c-Jun) could repress the activity of tgfb2b promoter. In vitro knockdown of tgfb2b…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
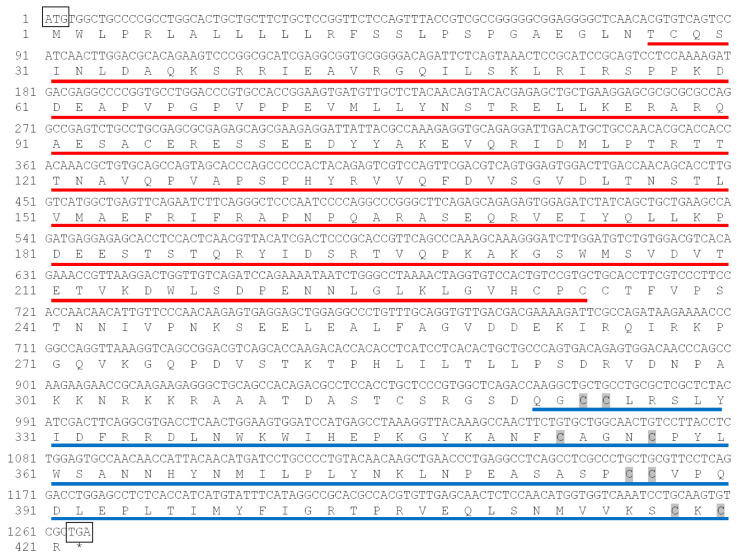 Figure 1
Figure 1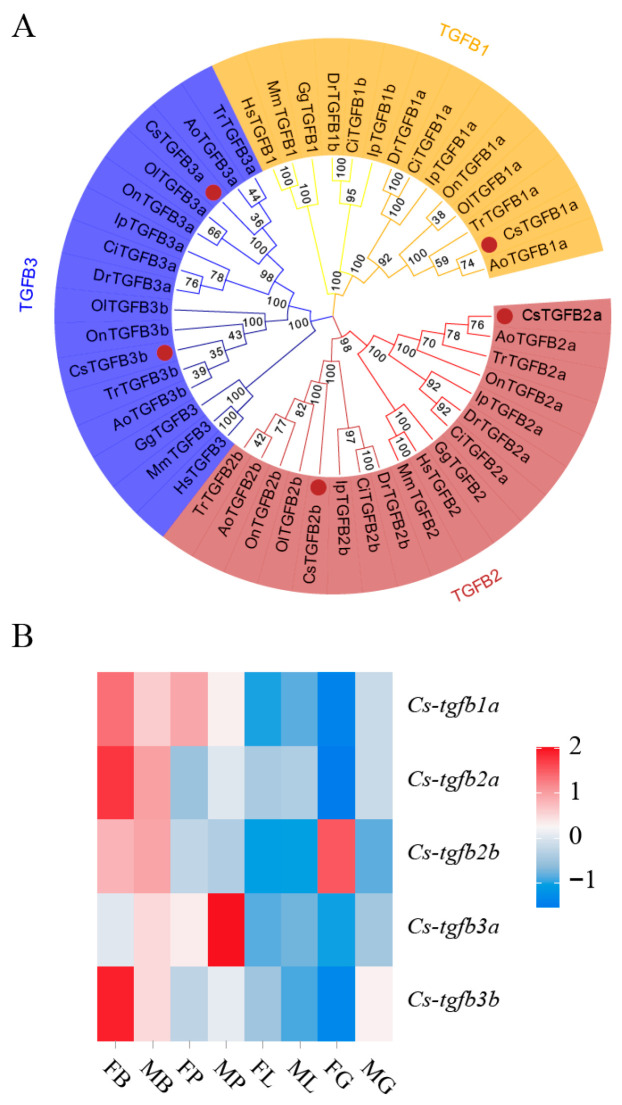 Figure 2
Figure 2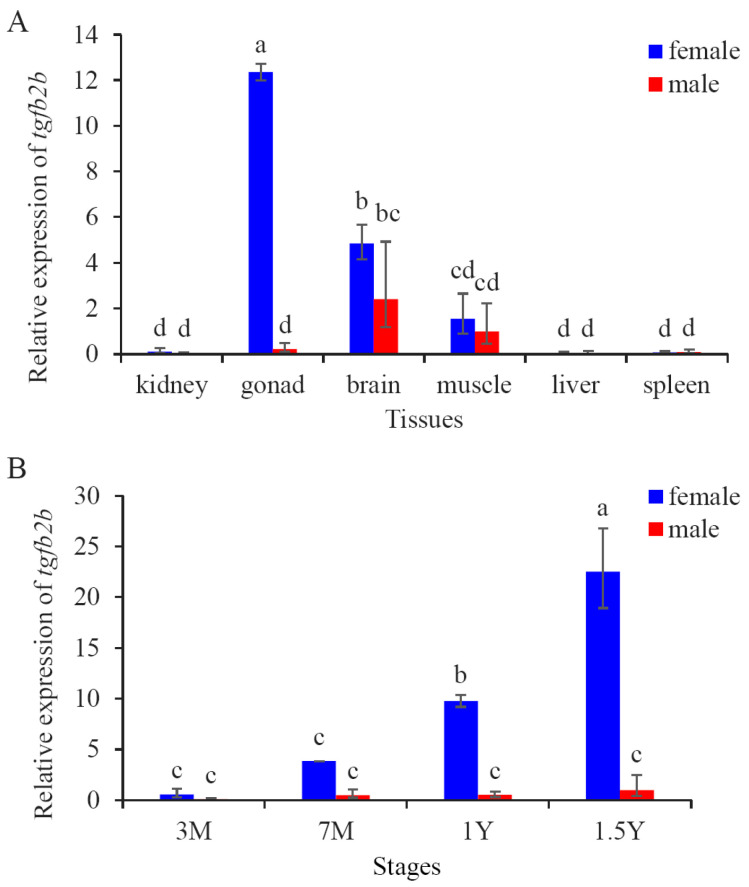 Figure 3
Figure 3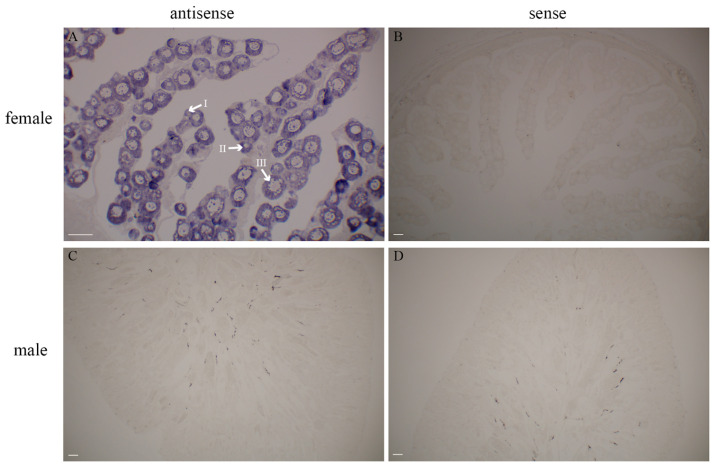 Figure 4
Figure 4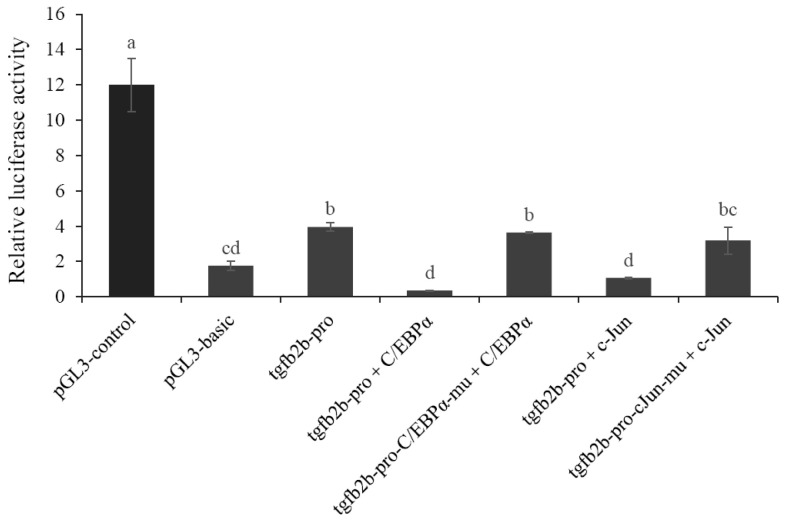 Figure 5
Figure 5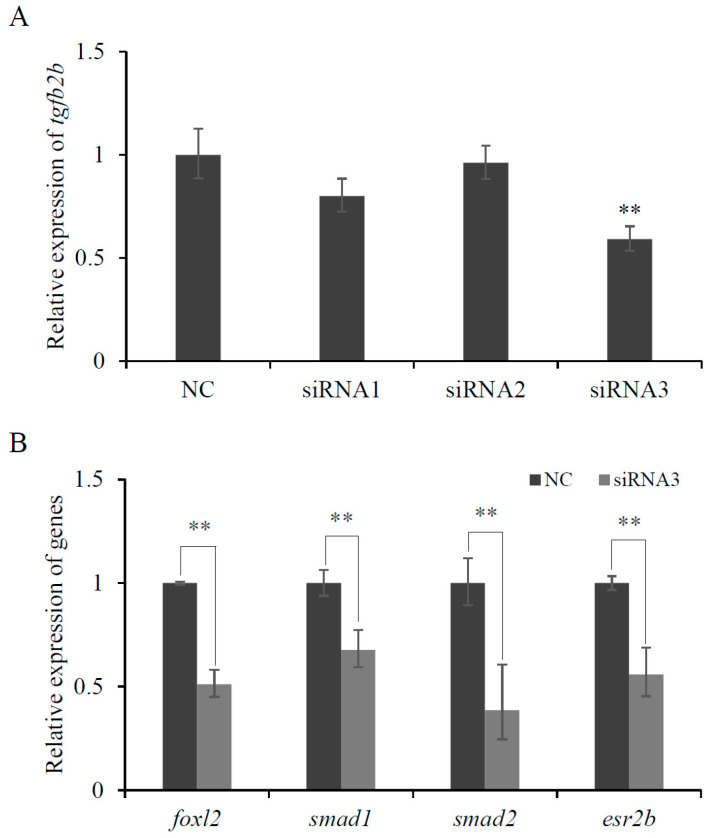 Figure 6
Figure 6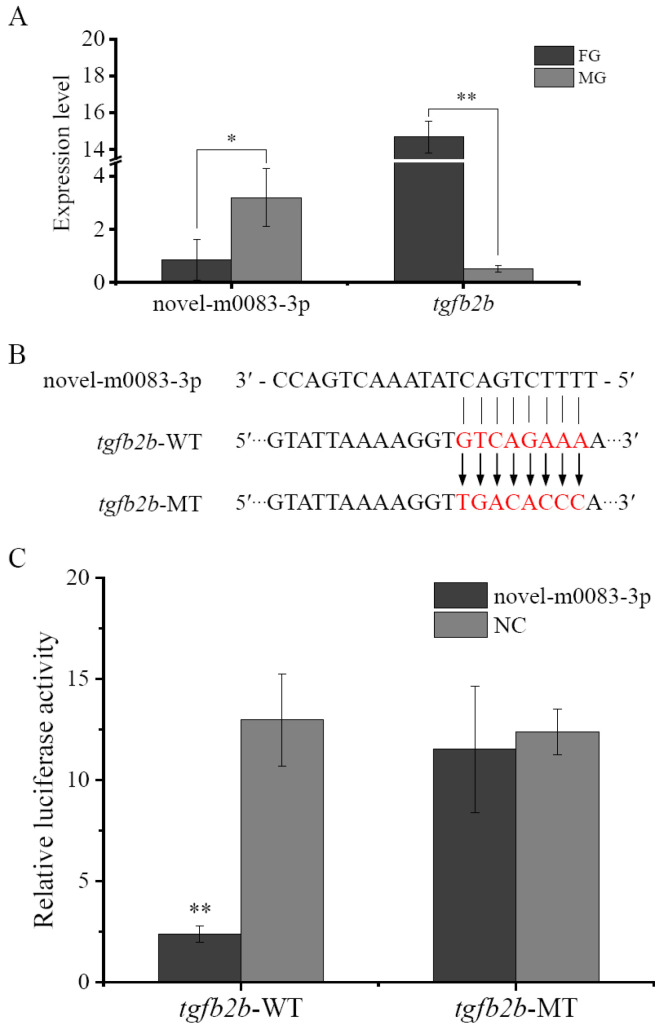 Figure 7
Figure 7- —STI2030-Major Projects
- —National Key Research and Development Program of China
- —Central Public-interest Scientific Institution Basal Research Fund, YSFRI, CAFS
- —Taishan Young Scholar Project of Shandong Province
- —China Agriculture Research System
- —Central Public-interest Scientific Institute Basal Research Fund, CAFS
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic and Clinical Aspects of Sex Determination and Chromosomal Abnormalities · Reproductive biology and impacts on aquatic species · TGF-β signaling in diseases
1. Introduction
As an economically important mariculture fish, the Chinese tongue sole (Cynoglossus semilaevis) has a female heterogametic ZW sex-determining system [1,2]. It exhibits sexual dimorphism that the females mature later and grow 2–4 times bigger than the males [2]. However, a high percentage of the males in farmed populations give rise to huge production and economic losses. Explaining the molecular mechanism underlying female sex formation and making effort for high-female-ratio fry will be helpful for the sustainable development of the aquaculture.
It is well known that sex development (including sex determination, differentiation, and maturation) is an important process, by which undifferentiated gonad becomes a testis or an ovary. It can be controlled genetically (genetic sex determination, GSD), environmentally (environmental sex determination, ESD), or by a combination of the two categories [3]. In fish species with GSD, various sex-determining genes have been identified. Several of them (e.g., dmy, dmrt1, and sox3-Y) are transcription factors, while many others including anti-Müllerian hormone (amh) receptor type II (amhr2), Y chromosome-linked duplicate of amh (amhy), gonadal soma-derived factor (gsdf), growth differentiation factor 6 (gdf6), and bone morphogenetic protein receptor (bmpr) are tightly associated with transforming growth factor β (TGF-β) superfamily [3,4,5]. Even in Oryzias latipes and O. dancena that separately utilize dmy and sox3-Y as sex-determining gene, mutations of the superfamily genes gsdf and amhr2 could cause dysregulation of testis/ovary development [6,7]. Other members of the superfamily such as gdf9, bmp15, and glial cell line-derived neurotrophic factor (gdnf) are also widely researched to be instrumental in sex differentiation [8,9,10]. For example, the oocyte-secreted factors gdf9 and bmp15 play important roles in follicle development, oocyte maturation, and steroidogenesis [8,9]. In zebrafish, gdf9 is presented in differentiating or differentiated gonads, with an expression pattern closely relating to ovarian development [11]. Knockdown of this gene decreases the number of female fish, indicating its involvement in facilitating oocyte/ovary differentiation [11]. Bmp15 deficiency in females also leads to sex reversal in the mid- to late-juvenile stage and results in fertile males, which implies the requirement in female differentiation and maintenance [12]. Regarding the different isoforms of TGF-β ligand genes (tgfb1/2/3), tgfb1 has been indicated to suppress zebrafish oocyte maturation, too [13]. However, those studies are mainly focused on part molecules of the superfamily, while relevant reports on tgfb2 and tgfb3 are still inadequate and need more investigations.
Due to genome duplication, tgfb genes often display paralog pairs (suffixing with “a” and “b”) in teleost [14]. Based on previous transcriptome data of C. semilaevis [15], we found five genes (tgfb1a, tgfb2a, tgfb2b, tgfb3a, and tgfb3b) and compared their expressions in somatotropic and reproductive tissues. Especially, tgfb2b was highest expressed in female gonad, with a significant sexually dimorphic pattern. In this study, to explore the potential role of tgfb2b in fish ovarian differentiation and development, the gene sequence was cloned and its expression profiles in different tissues and at different developmental periods were analyzed. In situ hybridization, promoter transcriptional activity, small interfering RNA (siRNA)-mediated knockdown effect, and microRNA (miRNA)-targeted regulation of the gene were also characterized. The results would contribute to better understanding the biological function of TGF-β superfamily in teleost.
2. Materials and Methods
2.1. Fish Samples
Fish individuals used for this study were cultured at Haiyang breeding base (Shandong, China). Before the experiments, they were anesthetized with MS-222 to reduce the pain. Genetic sex of the fish was determined using the previously reported method with the primers sex-F and sex-R (Table 1) [16]. Tissues including kidney, gonad, brain, muscle, liver, and spleen were randomly sampled from three female and three male 1-year-old (1 Y) C. semilaevis. At the same time, gonads of the fish at different developmental periods, such as 3 months, 7 months, 1 year, and 1.5 years post-hatching (3 M, 7 M, 1 Y, and 1.5 Y), were also gathered. Each period contained at least three female and three male individuals. The samples were subsequently stored at −80 °C until RNA isolation. The fish experiment was inspected and approved by the Animal Care and Use Committee at the Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences (Approve No.: YSFRI-2023006, approval date: 2 March 2023).
2.2. Cell Culture
Human embryonal kidney (HEK) 293T cells (ATCC^®^ CRL-3216™) purchased from ATCC (Manassas, VA, USA) and C. semilaevis ovarian (CSO) cells established by our lab [17] were used in this study. HEK 293T cells were grown in DME/F-12 medium added with 10% fetal bovine serum (FBS) (Gibco, Auckland, New Zealand) at 37 °C in a 5% CO_2_ incubator. L-15 medium comprising 15% FBS and bFGF (Invitrogen, Carlsbad, CA, USA) was selected for CSO cells, and the cells were cultured at 24 °C. One day prior to transfection, the cells were seeded in 12- or 24-well plates and allowed to grow to a density of 70–80%.
2.3. RNA Isolation and cDNA Synthesis
Total RNA of the tissues or cells was isolated using TRIzol reagent (Invitrogen). RNA quantity and quality was assessed via a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA) and agarose gel electrophoresis. Reverse transcription from the purified RNA to complementary DNA (cDNA) was conducted using HiScript^®^ III RT SuperMix with gDNA wiper (Vazyme, Nanjing, China), following the product instruction.
2.4. Gene Sequence Cloning and Analysis
According to the sequence information of tgfb2b in GenBank (ID: 103390834), specific primers tgfb2b-CDS-F and tgfb2b-CDS-R (Table 1) were designed to clone its coding sequence (CDS). The amplification product was ligated into pEASY-T1 vector (TransGen, Beijing, China) and sequenced by Qingdao Branch of TsingKe Biotech Co., Ltd. (Qingdao, China). Predicted domain of the sequence was analyzed using NCBI Conserved Domain Database (CDD, https://www.ncbi.nlm.nih.gov/cdd (accessed on 9 November 2023)) and SMART (http://smart.emblheidelberg.de/ (accessed on 9 November 2023)). After BLAST search at NCBI (http://www.ncbi.nlm.nih.gov/BLAST/ (accessed on 9 November 2023)), homologous amino acid sequences of different TGF-β isoforms from seven fish species (Danio rerio, Oryzias latipes, Oreochromis niloticus, Takifugu rubripes, Ctenopharyngodon idella, Ictalurus punctatus, and Anarrhichthys ocellatus) and three common non-fish species (Homo sapiens, Mus musculus, and Gallus gallus) were acquired (Supplementary Table S1). Multiple sequence alignment was completed adopting ClustalW program. A phylogenetic tree was generated by MEGA7 software (version 7.0.26) using Neighbor-Joining method with 1000 bootstrap replicates. Then, the tree was beautified and visualized in EvolView (http://www.evolgenius.info/evolview (accessed on 9 November 2023)). In addition, the normalized fragments per kilo base of transcript per million mapped reads (FPKM) values of C. semilaevis tgfb genes were obtained from our previous transcriptome data [15]. According to the values, an expression heatmap was drawn on Omicshare website (https://www.omicshare.com/tools (accessed on 9 November 2023)).
2.5. Expression Detection in Different Tissues and Periods
The expression patterns of tgfb2b in different tissues and at different developmental periods of female and male C. semilaevis were detected through quantitative real-time (qPCR) analysis. The above synthesized cDNA from 6 tissues (kidney, gonad, brain, muscle, liver, and spleen) at 1 Y and from the gonads at 4 developmental periods (3 M, 7 M, 1 Y, and 1.5 Y) were used for the analysis. The reliable reference gene β-actin was selected as internal control, and the primers were listed in Table 1. According to the standardized procedures, the reactions were performed using THUNDERBIRD™ Next SYBR^®^ qPCR Mix (TOYOBO, Tokyo, Japan) on a Quant Gene 9600 instrument (BIOER, Hangzhou, China). Each experiment included at least three biological replicates. Gene relative expression level was assessed using 2^−ΔΔCt^ method [18]. The data was presented as mean ± standard deviation (mean ± SD). Statistical analysis was performed by one-way analysis of variance (ANOVA) and least significant difference (LSD) multiple comparison with the online OmicShare tool (https://www.omicshare.com/tools (accessed on 18 April 2024)). p < 0.05 was accepted for significant difference.
2.6. In Situ Hybridization (ISH)
Ovary and testis sections of 1 Y C. semilaevis were prepared by our lab. Briefly, parts of the gonads were fixed using 4% paraformaldehyde, dehydrated by graded ethanol, embedded in paraffin, and cut into slices of about 5 μm thickness. A fragment (609 bp in length) of tgfb2b CDS was cloned with a pair of probe primers tgfb2b-SP6-F and tgfb2b-T7-R (Table 1). The amplified fragment was used to produce digoxigenin (DIG)-labeled antisense/sense probe with T7/SP6 RNA polymerase (Roche, Mannheim, Germany). Afterwards, the RNA probes were purified by lithium chloride (LiCl) precipitation. The detailed operation steps of ISH were elaborated in our previous studies [8]. Finally, the signals were examined by BCIP/NBT Chromogen Kit (Solarbio, Beijing, China) and photographed on a Nikon Eclipse 80i microscope (Nikon, Tokyo, Japan).
2.7. Promoter Amplification and Activity Analysis
The predicted promoter sequence (2041 bp upstream) of tgfb2b was amplified with the infusion primers tgfb2b-pF and tgfb2b-pR (Table 1). Using the One Step Fusion Cloning Mix (Toroivd, Shanghai, China), the amplification product was ligated into HindIII-digested pGL3-basic vector (Promega, Madison, WI, USA) to generate a recombinant luciferase reporter plasmid called pGL3-tgfb2b-pro. Then, the plasmid was transfected into HEK 293T cells in a 24-well plate. pGL3-basic (negative control) and pGL3-control (positive control) were also transfected to the cells, respectively. Each group contained 4 wells, and pRL-TK was used as internal reference. Lipo8000™ Transfection Reagent (Beyotime, Shanghai, China) was selected for the transfection referring to the instruction. After transfection for approximately 48 h, cells were harvested to asses firefly and Renilla luciferase activities by the Dual-Luciferase Reporter Gene Assay Kit (Beyotime) and a microplate reader (Thermo, Vantaa, Finland). The relative luciferase activity of each group was calculated as the average ratio of firefly to Renilla luciferase activity. Statistical analysis was performed the same as described in Section 2.5.
At the same time, transcription factor (TF) binding sites in tgfb2b promoter were searched on PROMO website (http://alggen.lsi.upc.es/ (accessed on 20 November 2023)). CDSs of possible TFs were amplified from C. semilaevis and inserted into pcDNA3.1 vector. In addition, point mutations of the binding sites in tgfb2b promoter were realized by Fast Site-Directed Mutagenesis Kit (Tiangen, Beijing, China). The primers were listed in Table 1. Cell co-transfections with pGL3-tgfb2b-pro (wild- or mutant-type) and TF plasmids were conducted in 24-well plates, following the procedure of Lipo8000™ Transfection Reagent (Beyotime). Each group contained 4 wells, too. Luciferase assay and statistical analysis were carried out with the above-mentioned methods.
2.8. RNA Interference (RNAi)
Three siRNA oligonucleotides (Table 2) targeting tgfb2b gene were designed and synthesized from Sangon Biotech Co., Ltd. (Shanghai, China). They were separately transfected into CSO cells with the help of riboFECT^TM^ CP Transfection Kit (Ribobio, Guangzhou, China). A randomly synthesized and nonspecific siRNA from the same company was applied as the negative control (NC). In addition, cyanin 3 (cy3)-labeled siRNA was used to determine the transfection efficiency by observing cy3 fluorescence under a fluorescence microscope (Olympus, Tokyo, Japan). The cells were collected after transfection for 24 h. RNA extraction, cDNA synthesis, and qPCR assay were subsequently carried out with the same methods as described above. Expression levels of tgfb2b and other related genes such as SMAD family members (smad1 and smad2), forkhead box L2 (foxl2), and estrogen receptor 2b (esr2b) were assessed using 2^−ΔΔCt^ method [18]. Statistical analysis between siRNA-transfected group and NC group was performed by t-test. p < 0.05 or p < 0.01 was considered as significant difference or extremely significant difference, respectively.
2.9. miRNA Prediction, Validation, and Transfection
Our whole transcriptomic data analysis [19] suggested that tgfb2b might be regulated by the miRNA novel-m0083-3p. Expression bar graphs of the miRNA and tgfb2b in C. semilaevis gonads were plotted based on the transcriptomic data [19]. To further verify the relationship between tgfb2b and the miRNA, a 3′-untranslated (3′-UTR) fragment of tgfb2b that comprised the binding site with novel-m0083-3p was cloned using the primers tgfb2b-0083-F and tgfb2b-0083-R (Table 1). The sequence was double-digested with XhoI and SalI and ligated into pmir-GLO vector, generating a GLO-tgfb2b-WT reporter plasmid. A mutant plasmid called GLO-tgfb2b-MT was also obtained by use of Fast Site-Directed Mutagenesis Kit (Tiangen) and the mutation primers (tgfb2b-0083-muF and tgfb2b-0083-muR, Table 1). Meanwhile, novel-m0083-3p mimics (Table 2) and a random-synthesized negative control (NC) were ordered from Sangon Biotech (Shanghai). Using Lipo8000™ Transfection Reagent (Beyotime), GLO-tgfb2b-WT/MT and novel-m0083-3p mimics/NC were transfected to HEK 293T cells, respectively. Two days later, the luciferase activity was measured by Dual-Luciferase Reporter Gene Assay Kit (Beyotime). The experiment was conducted in three replicates. Statistical analysis between every two groups was performed the same as described in Section 2.8.
3. Results
3.1. Sequence Characterization of tgfb2b
As shown in Figure 1, CDS of C. semilaevis tgfb2b gene contained 1266 bp. It encoded 421 amino acids with a TGF-β propeptide region in 27–234 residues, a TGF-β domain in 322–420 residues, and 8 conserved cysteines (C). As expected, the phylogenetic tree (Figure 2A) displayed that various TGF-β amino acid sequences were clustered into three clades (TGFB1, TGFB2, and TGFB3 clades). Each clade can be further divided into two small groups (e.g., TGFB2a and TGFB2b). Among them, C. semilaevis TGFB2b was closely grouped with the homologous sequences from O. latipes and I. punctatus. Expression heatmap revealed that, among different isoforms of C. semilaevis, only tgfb2b gene was highly expressed in female gonad (Figure 2B).
3.2. Expression Pattern and Cyto-Location of tgfb2b
To further evaluate the tissue expression of tgfb2b, qPCR experiments were completed in six tissues from 1 Y female and male C. semilaevis. The prominent expression of this gene appeared in ovary, but little expression was detected in testis (Figure 3A). It also showed moderate expression levels in brain and muscle, slightly higher in female than in male (Figure 3A).
The expression patterns of tgfb2b in gonads across distinct developmental periods were inspected, too. It was continuously expressed and showed gradual upregulation in ovaries from 3 M to 1.5 Y (Figure 3B). Nevertheless, the expression levels were constantly low in male gonads.
ISH results displayed the cellular localization of tgfb2b in gonads. In the ovary of 1 Y female individual, oocytes at various stages (stage I, II, and III) were obviously seen (Figure 4A). By contrast, the gene was scarcely distributed in C. semilaevis testis (Figure 4C). No expression signals were discovered with sense probes (Figure 4B,D).
3.3. Transcriptional Regulation of tgfb2b Promoter Activity
Dual-luciferase detection results (Figure 5) showed that relative luciferase activity of the cells transfected with tgfb2b promoter was 1.24-fold higher than that of pGL3-basic group, which indicated that it had a remarkable transcriptional activity in initiating gene expression. Moreover, the results also displayed that the activity could be significantly repressed by the TFs including CCAAT/enhancer binding protein α (C/EBPα) and Jun proto-oncogene AP-1 transcription factor subunit (c-Jun) (Figure 5). When the promoter sequence was mutated at C/EBPα or c-Jun binding site, the inhibitory effect was removed and the activity recovered back to the original level (Figure 5).
3.4. Knockdown Effect of tgfb2b siRNA in CSO Cells
To explore the function of tgfb2b, three specific siRNAs were transfected into CSO cells. In siRNA3-transfected group, the expression of tgfb2b declined to about 59% of the control group (Figure 6A). It showed extremely significant knockdown effect (p < 0.01) (Figure 6A). Thereafter, the expressions of several related genes such as foxl2, smad1, smad2, and esr2b were also detected to be noticeably downregulated (Figure 6B).
3.5. The Influence of Novel-m0083-3p on tgfb2b
As shown in Figure 7A, the expression of the miRNA novel-m0083-3p in female gonad was significantly lower than that in male gonad, appearing an opposite expression pattern with tgfb2b. To further confirm the regulatory relationship, recombinant luciferase reporter plasmids that contained wild-type and mutant-type binding sites (Figure 7B) were co-transfected with novel-m0083-3p mimics into HEK 293T cells, respectively. Compared with NC group, novel-m0083-3p mimics extremely significantly diminished the relative luciferase activity of the cells transfected with wild-type tgfb2b 3′-UTR (Figure 7C). However, no negative influence was detected when the cells were transfected with mutant plasmid (Figure 7C). The results proved that tgfb2b could be directly targeted and suppressed by novel-m0083-3p.
4. Discussion
TGF-β superfamily members participate in a wide range of cellular processes including cell growth and differentiation. They are reported to be crucial to sex development and gonad function [4,5,13,20]. In C. semilaevis, tgfb2b showed marked female-biased expression in the gonad, which pointed to its functional involvement in ovarian regulation. Like most sequences of the superfamily in other species [8,14], the gene encoded amino acids with a large prodomain region and a C-terminal mature TGF-β domain. It also comprised several conserved cysteine residues, which were helpful to form disulfide bonds, fold stabilized dimer, and then bind to specific receptor [14]. The conserved structure might be a key to the biological activity of this molecule.
In this study, five TGF-β paralogs of C. semilaevis were analyzed. They showed various expression patterns in different tissues. Among them, tgfb2b was highest expressed in ovary, while tgfb1a, tgfb2a, tgfb3a, and tgfb3b were mainly expressed in other tissues like brain and pituitary. Tgfb1a and tgfb3a were also quantified to be prominently expressed in skin of the fish, but with low expression level in muscle [21]. Their expressions exhibited time-dependent changes after Vibrio harveyi infection [21]. In addition, the genes were also revealed to be involved in early response to heat stress in the brain of female and male C. semilaevis [22]. This diversity suggests that TGF-β isoforms may exist with varying functions at different body parts [14].
Many TGF-β superfamily genes including bmp15, gdf9, activin, and tgfb1 are often observed with high abundance in ovary and serve as modulators of ovarian development/function [8,13,23,24]. In zebrafish, mutation of the oocyte-secreted factor bmp15 could start normal female development but switch sex to fertile male during the juvenile stage, proving that the factor is required for female sexual development and phenotype maintenance [12]. Another example, tgfb1, is detected in ovarian follicles at differing developmental periods, and can influence both gonadotropin- and 17α, 20β-dihydroxyprogesterone-triggered oocyte maturation in vitro [13]. However, as a member of TGF-β superfamily, the role of tgfb2b in fish ovary is not well understood. Coincidently, in the present study, the gene was tested to be ovary-enriched. Its expression throughout ovarian developmental processes from 3 months to 1.5 years after hatching tended to be gradually increased. It was also examined with localization signals in oocytes of different stages, from early small stage I through stage II and mature stage III. In C. semilaevis, at the early developmental period, there is nearly no difference between female and male gonads. Sex determination of the fish is prosed at about 50 days after hatching, and gonadal differentiation is initiated at 56–62 days. Nevertheless, for the reason of hysteresis, cellular differentiation often appears in the ovary at 3–4 months, along with the emergence of ovarian cavity. Afterwards, the oocyte goes on to differentiate, exhibiting obvious differentiation at 6–7 months and reaching sexual maturity at 1.5–2 years old [2,25,26]. Considering the fish ovarian developmental performance and the gene expression profile, these findings further implied the importance of tgfb2b in female differentiation and development of C. semilaevis.
The promoter activity of tgfb2b gene was successfully validated via dual-luciferase assay. After searching for transcription factor binding sites in the promoter region, we found that C/EBPα and c-Jun could efficiently repress its promoter activity. The two factors are known to bind with growth- and sex-related genes, adjusting gene expression and undertaking roles in sex regulation [27,28,29]. The binding of C/EBPα to cyp3a41 promoter can be impacted by sexually dimorphic growth hormone (GH) secretion and attributes to high gene expression level in female [30]. Moreover, C/EBPα/β could regulate ovulation and luteinization-related genes, affecting ovulation via dose- and preovulatory stage-dependent manners [27]. The transcriptional mediator is also involved in terminal differentiation of granulosa cells during luteinization [31]. Likewise, our findings that C/EBPα suppressed the activity of female-biased tgfb2b in C. semilaevis supported its potential role in female gonad differentiation and development. Additionally, the expression of mice tgfb2 is mediated by c-Jun transcription factor and involved in the establishment of Sertoli cell polarity [29]. Another JUN family member, JUNB, can bind to and decrease both the mRNA and protein levels of tgfb2, which then functions in cell proliferation and ovarian tumor aggressiveness [28]. Consistent with this, the binding of c-Jun to the promoter of C. semilaevis tgfb2b suppressed its transcriptional activity. However, the precise molecular mechanisms by which these transcription factors regulate tgfb2b function require further elucidation.
After in vitro knockdown of tgfb2b in CSO cells, the expression levels of several sex-related genes (e.g., foxl2, smad1, smad2, and esr2b) were declined. SMAD proteins are intracellular effectors that can be phosphorylated by TGFβ-stimulated receptors and deliver the signal. Female smad1/5 double conditional knockout (dKO) mice develop fertility defects and granulosa cell tumors with significant expression changes in TGF-β family genes, implying the molecule as part of a crucial pathway in ovary [32]. Foxl2 is also reported to be modified by TGF-β superfamily members [3,33]. It plays a pivotal role in ovarian differentiation by boosting the expression of cyp19a (an important female-related gene encoding aromatase that is responsible for estrogen synthesis) and antagonizing male pathway gene expression [34]. Deficiency of foxl2 leads to the failure of female gonad development, even female-to-male sex reversal, and the mutant phenotype could be rescued by 17β-estradiol (E2) [34,35]. In C. semilaevis, foxl2 and esr2b were both downregulated in response to RNAi of tgfb2b. The data indicated that TGF-β/SMAD signaling contributed to the fish female development, potentially via regulation of foxl2-cyp19a-esr axis.
Dual-luciferase reporter assay analysis proved that C. semilaevis tgfb2b could be bound and negatively regulated by the miRNA novel-m0083-3p. It was consistent with their converse expression profiles in gonads. In other species, tgfb2 is also reported to be targeted and transcriptionally or post-transcriptionally controlled by miRNAs [36,37,38]. Inhibition of tgfb2 by miR-301a-5p participates in germline cell differentiation and spermatogenesis [36], while regulation of miR-31 on tgfb2 and smad2 affects the developmental potential of oocytes [37]. Similarly, our results suggested the relevance of miRNA in TGF-β pathway-joined sex development of the fish. The detailed role and action mechanism should be forwardly examined.
5. Conclusions
In summary, tgfb2b exhibited female-dominant expression in gonad of C. semilaevis. It was continuously expressed and displayed a progressive increase throughout ovarian developmental periods from 3 months to 1.5 years after hatching. Transcriptional regulation, siRNA knockdown, and miRNA-targeted experiments supported that the gene played a considerable role in ovary differentiation and development. Further investigations on the crosstalk between tgfb2b and foxl2-cyp19a-esr will be essential to elucidate the broader sex-determination regulatory network in fish.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhou L. Yang A. Liu X. Du W. Zhuang Z. The karyotype of the tonguefish Cynoglossus semilaevis J. Fish. China 200529417419
- 2Chen S. Zhang G. Shao C. Huang Q. Liu G. Zhang P. Song W. An N. Chalopin D. Volff J.N. Whole-genome sequence of a flatfish provides insights into ZW sex chromosome evolution and adaptation to a benthic lifestyle Nat. Genet.20144625326010.1038/ng.289024487278 · doi ↗ · pubmed ↗
- 3Yu H. Du X. Chen X. Liu L. Wang X. Transforming growth factor-β (TGF-β): A master signal pathway in teleost sex determination Gen. Comp. Endocrinol.202435511456110.1016/j.ygcen.2024.11456138821217 · doi ↗ · pubmed ↗
- 4Pan Q. Kay T. Depince A. Adolfi M. Schartl M. Guiguen Y. Herpin A. Evolution of master sex determiners: TGF-β signaling pathways at regulatory crossroads Philos. Trans. R. Soc. B 20213762020009110.1098/rstb.2020.0091 PMC 827350734247498 · doi ↗ · pubmed ↗
- 5Kitano J. Ansai S. Takehana Y. Yamamoto Y. Diversity and convergence of sex-determination mechanisms in teleost fish Annu. Rev. Anim. Biosci.20241223325910.1146/annurev-animal-021122-11393537863090 · doi ↗ · pubmed ↗
- 6Imai T. Saino K. Matsuda M. Mutation of Gonadal soma-derived factor induces medaka XY gonads to undergo ovarian development Biochem. Biophys. Res. Commun.201546710911410.1016/j.bbrc.2015.09.11226408909 · doi ↗ · pubmed ↗
- 7Morinaga C. Saito D. Nakamura S. Sasaki T. Asakawa S. Shimizu N. Mitani H. Furutani-Seiki M. Tanaka M. Kondoh H. The hotei mutation of medaka in the anti-Mullerian hormone receptor causes the dysregulation of germ cell and sexual development Proc. Natl. Acad. Sci. USA 20071049691969610.1073/pnas.061137910417535919 PMC 1887600 · doi ↗ · pubmed ↗
- 8Shi R. Li X. Cheng P. Yang Q. Chen Z. Chen S. Wang N. Characterization of growth differentiation factor 9 and bone morphogenetic factor 15 in Chinese tongue sole (Cynoglossus semilaevis): Sex-biased expression pattern and promoter regulation Theriogenology 202218211912810.1016/j.theriogenology.2022.02.00335151001 · doi ↗ · pubmed ↗