Placenta-Driven Evolution: Viral Gene Acquisition and PEG10’s Essential Roles in Eutherian Placenta
Hirosuke Shiura, Moe Kitazawa, Tomoko Kaneko-Ishino, Fumitoshi Ishino

TL;DR
The placenta's evolution in mammals was driven by the integration of viral genes, which played key roles in the development of viviparity and the diversification of placental types.
Contribution
The paper introduces the concept of 'placenta-driven evolution,' linking viral gene acquisition to major evolutionary transitions in placental development.
Findings
Viral-derived genes like PEG10 and PEG11/RTL1 were essential in the evolution of therian viviparity and placental differentiation.
Extraembryonic tissues like the placenta and yolk sac allowed for the functional domestication of virus-derived genes.
The placenta may have acted as a driver of eutherian evolution by enabling the co-option of additional viral genes.
Abstract
Mammalian placentation represents one of the most striking evolutionary innovations among vertebrates, and accumulating evidence indicates that virus-derived genes—particularly the metavirus-derived PEG10 and PEG11/RTL1—have played indispensable but distinct roles: PEG10 in the emergence of therian viviparity and PEG11/RTL1 in the subsequent differentiation between marsupial and eutherian placental types. Notably, the metavirus-derived SIRH/RTL gene group, which includes PEG10 and PEG11/RTL1, exhibits unique and diverse functions not only in placenta development but also within microglia of the brain. Because microglia originate from yolk sac progenitors, these findings suggest that extraembryonic tissues such as the placenta and yolk sac provided permissive environments that enabled the retention, expression and functional domestication of virus-derived sequences. Once the placenta…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
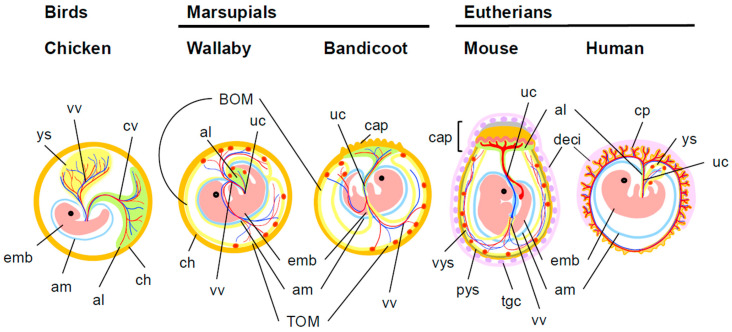 Figure 1
Figure 1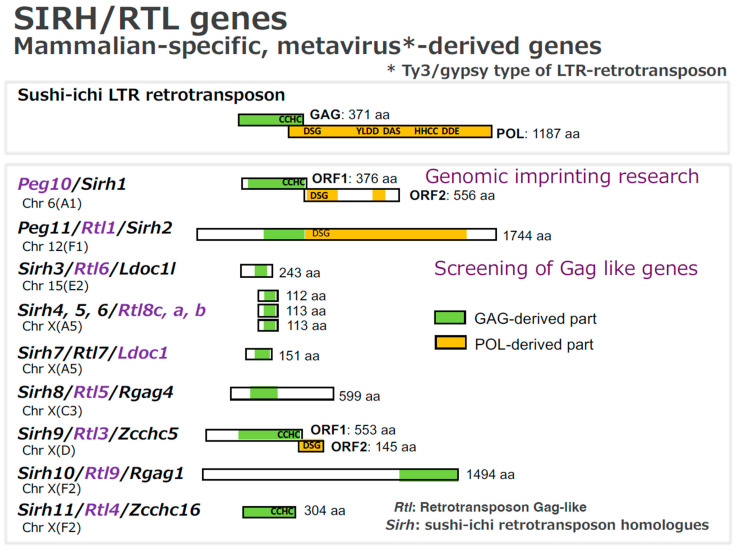 Figure 2
Figure 2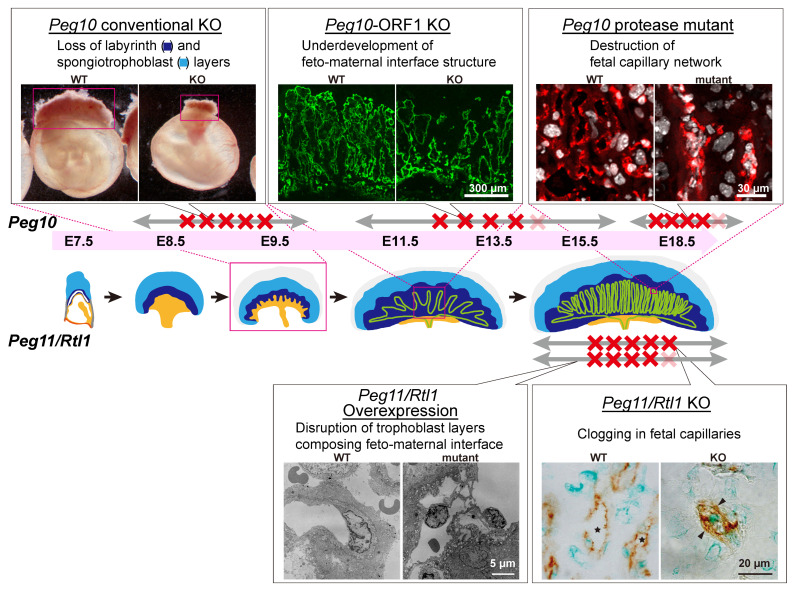 Figure 3
Figure 3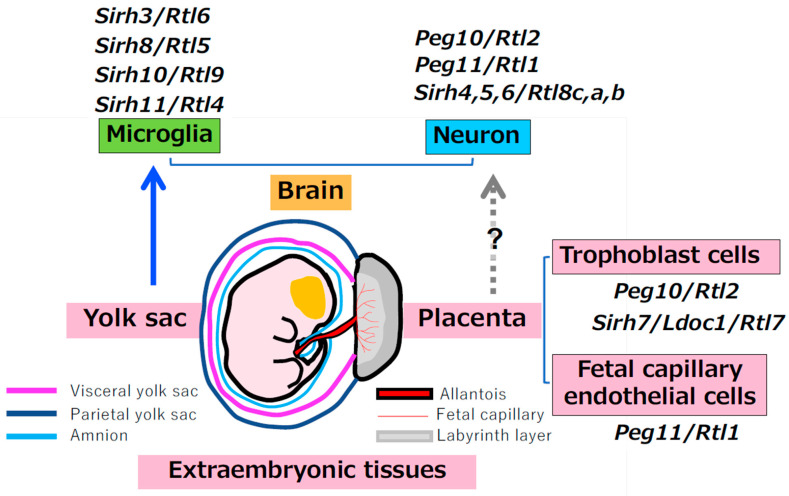 Figure 4
Figure 4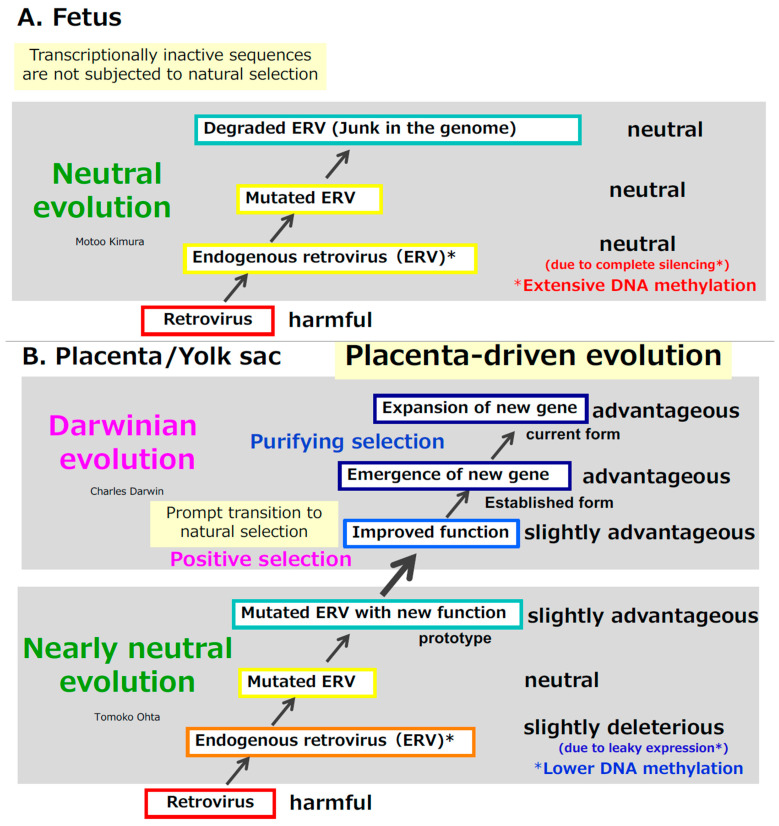 Figure 5
Figure 5- —JSPS
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEvolution and Paleontology Studies · Neuroendocrine regulation and behavior · Single-cell and spatial transcriptomics
1. Introduction
The emergence of the placenta was one of the most consequential events in mammalian evolution, enabling therian mammals to give birth to live young through viviparity [1,2,3,4]. How did this remarkable organ originate? Placental evolution likely reflects a synthesis of emergent processes—novel interactions among pre-existing biological components, such as fetal membranes and resident host genes—and innovative processes driven by the acquisition of new genes.
In eutherians, the placenta arises through fusion of the allantois and chorion (chorioallantoic placenta)—two fetal membranes conserved across amniotes (reptiles, birds, and mammals) [1,2]. In contrast, marsupial yolk sac placenta is formed through interactions between a different combination of two fetal membranes, the yolk sac and the chorion (choriovitelline placenta) [3,4]. Although these three fetal membranes are ancestrally shared among amniotes, their distinct combinations in therian mammals give rise to a fundamentally new organ with specialized and highly diversified biological functions—the placenta (Figure 1).
Large-scale knockout (KO) mouse studies have shown that placental abnormalities are extremely prevalent among embryonic lethal mutants [14,15,16]. These studies suggest that several thousand genes are required for early mammalian development [2,17], and because placental defects are observed in the majority of embryonic lethal mutants, it is likely that well over a thousand genes contribute, directly or indirectly, to placental development and function. In addition, a growing body of evidence indicates that a small number of virus-derived genes have made disproportionately important contributions to this evolutionary transformation, with PEG10 playing a particularly central role in the emergence of placentation, and PEG11/RTL1 serving as a key factor in establishing the eutherian-type of chorioallantoic placenta [18,19,20]. In the mouse placenta, at least five virus-derived genes are known to be essential for its formation, function, and maintenance [20]. Thus, the origin of the placenta depended on the acquisition of genes from external sources—primarily metaviruses and retroviruses—and the domestication of their molecular machinery. In this sense, viral gene acquisition represents a key evolutionary event underlying the establishment of viviparity in eutherian mammals. Considering how such genes were likely acquired raises an intriguing possibility.
In this review, we summarize the essential roles of virus-derived genes in placental development and evolution, explore the mechanisms by which such genes are acquired and domesticated, and propose a conceptual framework for understanding mammalian genomic innovation—the “placenta-driven evolution” hypothesis, which describes how subsequent evolutionary trajectories were shaped by the presence of the placenta through the recurrent acquisition and domestication of such virus-derived genes.
2. Results
2.1. Placental Development and Evolution: Five Virus-Derived Genes Essential for the Eutherian Placenta
Eutherian mammals possess eleven sushi-ichi retrotransposon homolog (SIRH)/retrotransposon Gag-like (RTL) genes derived from metaviruses, the formerly classified as Ty3/gypsy LTR retrotransposons in their genomes [18,19,20,21,22,23,24,25,26,27,28,29,30] (Figure 2). Among these, at least three SIRH/RTL genes are known to play essential roles in placental formation, maintenance and functions, together with two syncytin genes—retrovirus-derived envelope (ENV) genes that arose uniquely in the eutherian lineage [31,32,33,34,35].
All SIRH/RTL genes encode proteins exhibiting homology to the GAG of the sushi-ichi LTR retrotransposon. However, each protein has a distinct amino acid sequence and differs markedly in protein length, resulting in unique and functionally divergent biological roles. Among them, PEG10, PEG11/RTL1 and SIRH9/RTL4 retain POL-derived domains. Evolutionary conservation patterns also differ among the SIRH/RTL genes. PEG10 is therian-specific and conserved in both marsupials and eutherians, whereas PEG11/RTL1 and the remaining SIRH/RTL genes are eutherian-specific. Among eutherians, PEG10, PEG11/RTL1, SIRH3/RTL6, SIRH4, 5, 6/RTL8c, a, b and LDOC1/SIRH7 are conserved across all lineages examined. By contrast, SIRH8/RTL5, SIRH9/RTL4, SIRH10/RTL9 and SIRH11/RTL4 show lineage-specific degeneration or gene loss in certain species.
2.1.1. Therian-Specific, Metavirus-Derived PEG10 Has Multiple, Stage-Specific, Essential Roles in the Placenta
PEG10 is a therian-specific gene conserved in both marsupials and eutherians, representing the earliest metavirus-derived gene integrated during mammalian evolution [23,24,25,26,27,28]. PEG10 proteins retain clear homology with the GAG and POL proteins of the sushi-ichi LTR retrotransposon [23,24,25]. However, the protein similarity between PEG10 and the ancestral GAG/POL proteins is only approximately 20–30%, indicating that a substantial number of mutations must have accumulated following the original metaviral insertion before PEG10 became functionally established [24]. We refer to this type of domesticated gene as an “acquired gene”. Peg10–null mice exhibit early embryonic lethality due to the complete absence of two major trophoblast-derived structures—the labyrinth layer and spongiotrophoblast layer [28] (Figure 3, top left). Because trophoblasts are unique to the mammalian placenta [1], these findings demonstrate that PEG10 is indispensable not only for trophoblast lineage specification at the earliest stages of placental development but also for the establishment of viviparity itself.
Recent studies reveal that PEG10 performs multiple essential and stage-specific roles through its complex gene structure, which retains several characteristics of its metavirus origin. PEG10 encodes two proteins—PEG10-ORF1 and a PEG10-ORF1/ORF2 fusion protein—generated through a − 1 ribosomal frameshift mechanism conserved across retroviruses and metaviruses [23,24,25,40,41,42,43]. Both proteins are further processed by an intrinsic DSG protease, producing multiple cleaved fragments. Remarkably, all these features are conserved across therian mammals, strongly suggesting that each protein—and potentially each processed fragment—fulfills distinct roles during placental morphogenesis.
Consistent with these characteristics, PEG10-ORF1 KO mice exhibit mid- to late-gestational lethality [44], in contrast to Peg10-null mice that die much earlier. ORF1-deficient embryos display delayed development of the fetal capillary network within the labyrinth layer. At embryonic day 10.5 (E10.5), clusters of undifferentiated trophoblast cells persist, and the labyrinth trophoblast progenitor (LaTP) cells fail to appear at the appropriate developmental time [44,45,46]. In mature placentas, fetal capillary endothelial cells are normally surrounded by two syncytiotrophoblast layers (SynT-I and SynT-II) and sinusoidal trophoblast giant cells (sTGCs), all derived from LaTP cells [45,46]. In ORF1 mutants, this architecture is disorganized and coarse, compared with the fine and dense network seen in wild type placenta, and over 80% of embryos die before birth [44] (Figure 3, top middle).
A PEG10-ASG mutant—harboring a catalytic DSG → ASG substitution in the protease domain—exhibits yet another phenotype. Although placental morphology initially appears normal until mid-gestation, the fetal capillary network progressively collapses during late gestation, again resulting in more than 80% perinatal lethality [47] (Figure 3, top right).
Together, these findings demonstrate that PEG10 functions as a multi-component, multi-stage regulator of placental development. Through its complex metavirus-derived architecture, PEG10 promotes trophoblast differentiation in both the spongiotrophoblast and labyrinth layers and safeguards the fetal capillary network throughout gestation. The totality of evidence strongly supports the idea that PEG10 acquisition represented a pivotal evolutionary event, enabling the emergence of the placenta and establishing the foundation for viviparity in therian mammals.
2.1.2. Eutherian-Specific, Metavirus-Derived PEG11/RTL1 and LOC1/SIRH7/RTL7 Are Essential for the Establishment of the Eutherian-Type Placenta
While PEG10 is essential for the fundamental formation of the therian placenta, two additional metavirus-derived genes—PEG11/RTL1 and LDOC1/SIRH7/RTL7—are required for constructing the eutherian-type placenta that supports the prolonged gestation characteristic of eutherian mammals [20].
PEG11/RTL1 encodes a protein homologous to sushi-ichi GAG and POL, including an active DSG protease motif similar to that of PEG10 [48,49]. Unlike Peg 10, however, Peg11/Rtl1 is expressed in fetal capillary endothelial cells and adjacent SynT-II cells [36,50]. It plays a central role in maintaining the internal integrity of the fetal capillaries. Loss of Peg11/Rtl1 results in severe late-gestational lethality caused by occlusion of fetal capillaries due to excess proliferation and invasive activity of SynT-II cells [36,51] (Figure 3, bottom right). Conversely, Peg11/Rtl1 overexpression produces the opposite defect: excessive vacuolization and degeneration of SynT-II cells and abnormal enlargement of fetal capillary lumens, leading to fetal edema [36,51] (Figure 3, bottom left). Because Peg 10 is also expressed in SynT-II cells [47], cooperative interactions between Peg 10 and Peg11/Rtl1 likely contributed to the evolutionary elaboration of the fetal capillary system required for extended gestation.
LDOC1/SIRH7 encodes a small protein (~135 amino acids) homologous to sushi-ichi GAG [52,53]. Like PEG10, it is expressed in all trophoblast lineages and is essential for proper trophoblast differentiation and maturation. In Ldoc1/Sirh7 knockout mice, the normal three-layered placental structure is disrupted, characterized by invasion of the spongiotrophoblast layer into the labyrinth layer [52]. In addition to structural roles, the placenta functions as a major endocrine organ, producing progesterone (P4), placental lactogens in trophoblast giant cells (TGCs), and more than 20 prolactin-related hormones in spongiotrophoblast cells [54,55,56,57]. Consistent with this endocrine function, Ldoc1/Sirh7 KO mothers exhibit delayed parturition caused by impaired placental hormone regulation, particularly P4; as a result, many newborns die shortly after birth, due to abnormal maternal behavior possibly caused by delayed parturition [52]. These findings demonstrate that LDOC1/SIRH7 is indispensable not only for placental architecture but also for the endocrine regulation necessary for sustained gestation and successful birth in eutherian mammals.
2.1.3. Rodent-Specific, Retrovirus-Derived Syncytin-A and Syncytin-B Are Indispensable for Syncytiotrophoblast Formation
Syncytins have been repeatedly and independently acquired in various mammalian lineages from distinct retroviral ENV genes, yet they share a conserved function: mediating trophoblast cell–cell fusion to generate syncytiotrophoblasts [31,32,33,34,35,58,59]. In mice, the rodent-specific syncytin-A and syncytin-B genes fulfill this role, whereas humans and other primates possess syncytin-1 and syncytin-2. Targeted disruption of syncytin-A—expressed in SynT-I cells—causes embryonic lethality at mid-gestation (E12–13) [58]. Combined disruption of both syncytin-A and syncytin-B leads to embryonic lethality even earlier, demonstrating that these retrovirus-derived fusogens are essential for placental development [59].
The two syncytiotrophoblast layers surrounding the fetal capillary endothelial cells form a continuous syncytial sheet through virus-derived cell fusion. This structure is thought to provide a key immunological barrier against the maternal immune system. Consistent with this view, syncytin-B exhibits measurable immunosuppressive activity [59,60]. Because fetal-derived cells directly contact maternal tissues during placental invasion and capillary formation, protection from maternal immunity is essential for maintaining pregnancy.
According to the “baton-passing hypothesis,” syncytin genes may undergo sequential replacement whenever newly invading retroviruses contribute ENV genes with more efficient fusogenicity [60,61]. If so, the common ancestor of therian mammals—including the lineages leading to both eutherians and marsupials—may have harbored an ancestral syncytin gene critical for forming a primitive placenta.
In summary, at least five virus-derived genes—PEG10, PEG11/RTL1, LDOC1/RTL7 and two syncytin genes (syncytin-A and syncytin-B in mice and syncytin-1 and syncytin-2 in humans)—perform essential, nonredundant functions in establishing, maintaining, and elaborating the eutherian placenta. To our knowledge, the placental defects observed in Peg10 KO, PEG10-ORF1 KO, PEG10-ASG mutant, Peg11/Rtl1 KO, and syncytin KO mice are phenotypically distinct and unique, with no comparable abnormalities identified among the large and diverse collection of placental mutants reported to date [2,15,17]. This uniqueness strongly suggests that virus-derived genes have assumed specialized and nonredundant functions in placental development. Thus, their integration into mammalian genomes constituted a key evolutionary event in the origin of viviparity and subsequent diversification of eutherian mammals.
2.2. Roles of SIRH/RTL Genes in Yolk Sac-Derived Microglia of the Brain
In addition to their indispensable roles in the placenta, SIRH/RTL genes have also acquired essential functions in the eutherian brain (Table 1). Their roles in the neurons have been summarized elsewhere [20], including the overexpression of PEG10 in Angelman syndrome (AS) and amyotrophic lateral sclerosis (ALS) [62,63,64,65,66], the overexpression and deficiency of PEG11/RTL1 in Kagami–Ogata syndrome (KOS14) and Temple syndrome (TS14) [36,37,51,67,68,69,70,71], the lack of Ldoc1/Sirh7 in abnormal maternal behavior [52], and the reduced levels of Sirh4/5/6 (Rtl8c/a/b) in Prader–Willi syndrome-like phenotypes [72]. These findings reveal that SIRH/RTL genes initially expressed highly in the placenta have secondarily acquired diverse and substantial functions in the nervous system [20].
Particular interest lies in the distinct subset of SIRH/RTL genes functions in microglia, the sole immune cell lineage residing within the brain. Examples of this specialization include SIRH11/RTL4/ZCCHC16, a causative gene for autism spectrum disorder (ASD) [73,74,75], and SIRH3/RTL6, SIRH8/RTL5 and SIRH10/RTL9, which participate in pathogen defense [76,77], illustrate this specialization. The innate immune system—highly conserved across most animals—typically relies on Toll-like receptors that recognize generic pathogen-associated molecular patterns (PAMPs) [78,79,80]. However, our recent work shows that eutherians have elaborated this system further: these virus-derived SIRH/RTL genes enable microglia to directly recognize and eliminate specific PAMPs in a highly specialized manner, representing a striking example of molecular innovation through viral gene domestication [74,77]. Importantly, Sirh11/Rtl4 KO mice exhibit not only abnormal behaviors—such as increased impulsivity, decreased ability to adapt to new environments, and impaired short spatial memory—but also a delayed recovery of noradrenaline levels in the prefrontal cortex [74]. Consistent with this, the SIRH11 protein appears to respond to psychological stressors, possibly by sensing changes in noradrenaline release in the brain [75].
Microglia originate from the yolk sac and migrate into the brain during early embryogenesis [81,82]. Alongside the subgroup that functions in the placenta, this developmental origin underscores that most SIRH/RTL gene subgroups retain deep ties to extraembryonic tissues. Importantly, however, their subsequent diversification has proceeded along two major trajectories—one toward neuronal functions and the other toward microglial immune functions. Together, these dual directions demonstrate that the SIRH/RTL repertoire has been repeatedly co-opted to shape both neural circuitry and innate immune specialization in the eutherian brain.
Taken together, these observations strongly support the idea that extraembryonic tissues, such as the placenta and yolk sac, served as suitable “birthplaces” for the virus-derived genes [19,20,76,83] (Figure 4). This extraembryonic origin may provide the developmental and epigenetic conditions that enabled SIRH/RTL genes to be recruited not only into placental function but also into neuronal and microglial roles, thereby linking viral gene domestication to both placental and brain evolution in eutherian mammals.
2.3. How Were Virus-Derived Genes Acquired in Therian Mammals?—A Two-Step Evolutionary Model for the Acquisition of Viral Genes in the Placenta
Taking retroviruses as an example, once they are integrated into the germline as endogenous retroviruses (ERVs), their expression in the fetus becomes completely silenced through extensive DNA methylation [84,85]. As a result, they behave as neutral sequences. Over time, however, transcriptionally inactive ERVs accumulate mutations and become progressively degraded, as they are not subject to natural selection (Figure 5A). In this way, any newly arising mutations in ERVs within the germline cannot be functionally tested in the fetal tissues.
In contrast, in extraembryonic tissues such as the placenta and yolk sac, ERVs exhibit leaky expression owing to substantially lower levels of DNA methylation (Figure 5B) [86,87]. This creates a unique environment in which the placenta can, in effect, “test” whether these viral elements are harmful, neutral, or potentially beneficial within a relatively protected developmental context. As mutations accumulate, some ERV sequences may shift from slightly deleterious to effectively neutral and eventually to slightly advantageous. Once a sequence acquires a slightly advantageous function, Darwinian positive selection acts to refine and optimize it. In this way, an initially foreign viral sequence can ultimately become established as a functional gene. Thereafter, purifying selection preserves and stabilizes the gene within the population (Figure 5B).
Tomoko Ohta extended Kimura’s neutral theory into the “nearly neutral theory,” proposing that slightly deleterious mutations can become fixed when the effective population size is small [88,89,90]. Our scenario fits well within this theoretical framework: leaky expression in the extraembryonic tissues would have rendered ERV-derived sequences slightly deleterious, and mammals—particularly early ancestral mammals—likely had small effective population sizes, facilitating the fixation of such sequences by genetic drift.
Thus, the gene acquisition from retroviruses can be explained by a two-step evolutionary process: an initial phase driven by genetic drift, as predicted by the nearly neutral theory, followed by Darwinian natural selection acting on sequences that acquire beneficial functions. The presence of the placenta enabled the ERV sequences to be expressed and evaluated, allowing them to accumulate multiple advantageous mutations and ultimately transform into new functional genes by acting as an “experimental laboratory” or “evolutionary testing ground” [19,20,76,83] (Figure 5B).
Importantly, tissues that maintain low levels of DNA methylation during key developmental windows—most notably the placenta, the yolk sac, and even the preimplantation embryo—constitute privileged environments for the acquisition and domestication of virus-derived genes.
2.4. Placenta-Driven Evolution
Extending this logic, extraembryonic tissues—particularly the placenta—may have acted as an engine of mammalian evolution, continuously accelerating genomic innovation through the recurrent acquisition of virus-derived genes. Because the placenta represents the most prominent and influential site of such events, we refer to this process as “placenta-driven evolution” (Figure 5B).
The earliest metavirus-derived gene in vertebrates is activity-regulated cytoskeleton-associated protein (ARC), which was acquired in the vertebrate lineage around the emergence of tetrapods (350–400 Ma) [30,91]. The second acquisition corresponds to the SCAN family genes, which arose in the common amniote ancestor (~310 Ma) [91]. Although the corresponding metavirus remains unidentified, the SCAN-GAG clearly shows homology to the Gypsy/Ty3 GAG [91]. SASPase (ASPRV1) encodes a retroviral-like aspartic protease and likely originated from an ancient retrovirus or retrovirus-derived element integrated into the mammalian genome (~168 Ma). It plays an essential role in the maintenance of the mammalian skin barrier [92,93,94]. This represents an example in which a POL-derived protease has been domesticated to fulfill an essential physiological function in mammals. Although its protease domain resembles retroviral Pol-encoded proteases, the specific retrovirus or LTR retrotransposon progenitor has not been definitively identified. Later, PEG10 was acquired in the common therian ancestor approximately 148–166 Ma [29]. Importantly, aside from the expansion of SCAN genes through extensive gene duplication in the mammalian lineage, no other metavirus-derived genes appear to have been retained in the therian genomes between the ARC and PEG10 domestication events, suggesting the gene acquisition from metavirus was extremely rare during this long interval [30].
In striking contrast, following the acquisition of PEG10, eutherian mammals gained ~30 metavirus-derived genes, including PEG11/RTL1, other SIRH/RTL genes and the paraneoplastic Ma antigen (PNMA) family genes [95,96,97,98,99,100], all of which originated after the marsupial–eutherian split (~148 Ma) [30,91]. This burst of gene acquisition raises the possibility that the emergence of the eutherian type of placenta facilitated or promoted the recurrent domestication of metavirus-derived genes. This pattern strongly supports our “placenta-driven evolution” hypothesis.
In contrast, only three metavirus-derived genes have been identified in the marsupial lineage: PEG10 and two marsupial-specific genes, SIRH12 [101] and PNMA-MS1 [102]. This disparity likely reflects fundamental differences in placental biology, particularly the duration and physiological performance of the placenta. Marsupials rely primarily on a yolk sac placenta that functions only briefly during a short gestation period, whereas eutherians utilize a long-acting chorioallantoic placenta that supports sustained maternal–fetal exchange throughout extended gestation [1,2,3,4]. Such prolonged extraembryonic environments may create expanded opportunities for viral gene domestication (see Section 3).
Mammalian genomes also contain more than ten thousand ERV-derived protein-coding sequences encoding peptides longer than 80 amino acids, most of which remain uncharacterized, with the notable exception of the SIRH/RTL genes and syncytins [103]. Importantly, a subset of these sequences has recently been identified by several proteomics studies [104,105,106], indicating that they are indeed translated and should therefore be regarded as bona fide genes. A comparative genomics and proteomics study by Wang and Han (2020) identified 177 retrovirus-derived genes across vertebrates—including fish, amphibians, reptiles, birds, and mammals—138 of which are enriched in mammals, with a stark contrast of 136 in eutherians versus only 2 in marsupials [107]. This remarkable enrichment in mammals among vertebrates and in eutherians among therians strongly suggests that the eutherian lineage recruited viral genes at a substantially higher rate, likely facilitated by the permissive epigenetic environment of the chorioallantoic placenta. These patterns further support our “placenta-driven evolution” hypothesis.
Taken together, these observations support the view that the placenta first emerged as an essential reproductive organ through viral gene integration, and subsequently served as a powerful engine of “placenta-driven evolution” through recurrent acquisition and domestication of virus-derived genes.
3. Discussion
From a morphological perspective, it is notable that the eutherian chorioallantoic placenta has diversified into several structurally and functionally distinct forms—hemochorial, endotheliochorial, and epitheliochorial [108,109,110]. How such diversity arose among eutherian lineages remains an important evolutionary question. The lineage-specific acquisition of syncytin genes likely contributed to these differences, given their essential roles in trophoblast fusion and placental morphogenesis. However, it is almost certain that additional mechanisms—including gene duplication, modification of pre-existing genes, and changes in regulatory networks—have also played substantial roles. Furthermore, multiple retrovirus-derived sequences have been independently acquired in different eutherian lineages, and some have been shown to be protein-coding [103]. The placenta-driven evolution hypothesis aligns well with this view and further suggests that numerous yet-uncharacterized virus-derived genes may have contributed to lineage-specific diversification of placental morphology and function.
Moreover, determining which placental type characterized the last common ancestor of eutherians will provide a critical foundation for reconstructing the evolutionary trajectory of placental diversification. Phylogenetic analyses suggest that highly invasive types, such as hemochorial or endotheliochorial placentas, represent the ancestral state, whereas less invasive forms, such as epitheliochorial placentas, are derived [109,110]. This view is supported by recent studies of maternal-side uterine gene expression, which also favor a hemochorial ancestral condition [111]. Because both mice and humans possess hemochorial placentas, the phenotypes observed in knockout mice for PEG10, PEG11/RTL1, and SIRH7/LDOC1/RTL7—all conserved across eutherians—provide valuable insights into the developmental logic of the ancestral eutherian placenta. In this sense, these functional data offer a powerful framework for reconstructing placental evolution by linking molecular innovation to the morphological and physiological diversification of the therian as well as eutherian lineages.
Virus-derived genes appear to have disproportionately contributed to organs that underwent major evolutionary innovations in mammals, including humans, particularly the placenta and the brain. It is likely that such proteins provided pre-adapted molecular solutions to challenges unique to these organs. These observations suggest that viral gene domestication may have facilitated rapid functional diversification in evolutionarily novel tissues. In mammalian evolution, metavirus-derived SIRH/RTL genes began with the acquisition of PEG10 in the common ancestor of therian mammals [19,20]. Recent studies have demonstrated that PEG10 performs multiple, indispensable functions during placental development. In addition to driving trophoblast proliferation and differentiation in both the spongiotrophoblast and labyrinth layers at early stages [28], PEG10-ORF1 is essential for the differentiation of syncytiotrophoblasts and sinusoidal trophoblast giant cells that envelop fetal capillaries in the labyrinth [44]. Moreover, PEG10-DSG protease is required to maintain the integrity of the fully formed fetal capillary network throughout gestation [47]. Thus, a single virus-derived gene orchestrates the formation of several fundamental placental structures and ultimately maintains the fetal capillary network—the core functional unit of the placenta. These findings underscore how critical the acquisition of PEG10 was for the emergence of the eutherian placenta.
Although PEG10 is present in both marsupials and eutherians, marsupials predominantly rely on a yolk sac placenta [3,4,8,20] (Figure 1). Why, then, did these two major therian lineages diverge so markedly in the type of placenta they employ? The observation that bandicoots—a marsupial species—form a chorioallantoic placenta shortly before birth, in addition to a yolk sac placenta, indicates that marsupials are not inherently incapable of producing a eutherian-type placenta [8]. Rather, this suggests that the decisive difference lies not in developmental potential but in the genetic components required to sustain prolonged placental function. A likely factor is the presence or absence of PEG11/RTL1 [18,19,20,48,112]. Marsupial gestation periods are generally short, and in bandicoots the chorioallantoic placenta operates for only two to three days [8]. Even if such a placenta can form, the inability to maintain the fetal capillary network for an extended duration severely limits its functional capacity. Without PEG11/RTL1—an essential gene for preserving the internal integrity of fetal capillaries throughout gestation in eutherians [36]—marsupials may simply have been unable to exploit the full potential of the chorioallantoic placenta over a prolonged pregnancy. From this perspective, marsupials may have adopted a reproductive strategy optimized for short gestation, relying primarily on the yolk sac placenta despite retaining a latent capacity to form a chorioallantoic placenta. In contrast, the acquisition of both PEG10 and PEG11/RTL1 in the eutherian lineage provided the genetic foundation for maintaining a functional chorioallantoic placenta throughout long gestation. Acting together, these two viral genes were likely indispensable for the extended fetal growth characteristic of eutherian reproduction [18,19,20] (Table 2).
Building upon this idea—that differences in gene complement may have constrained the duration and functionality of the marsupial placenta—we next consider how these developmental disparities may have shaped the broader evolutionary trajectory of viral gene acquisition in therian mammals. If eutherians alone were capable of sustaining a long-acting chorioallantoic placenta, this would have fundamentally altered the selective and developmental environment in which viral genes could be domesticated. In line with the “placenta-driven evolution” hypothesis, the placenta may have served as a permissive site for the acquisition and domestication of virus-derived sequences, thereby facilitating repeated recruitment events. If this is the case, the effective duration of placental function—during which the tissue exists, remains transcriptionally active, and maintains relatively low DNA methylation—would directly influence the likelihood of viral gene acquisition. From this perspective, the striking contrast in the number of SIRH/RTL genes between eutherians—11 genes, including PEG10, PEG11/RTL1, and SIRH3 through SIRH11—and marsupials, which possess only PEG10 and SIRH12 [24,101], may reflect these markedly different placental characteristics. A similar pattern is observed for the PNMA gene family, another metavirus-derived group: eutherians retain nearly twenty PNMA genes [99], whereas marsupials possess only a single member, PNMA-MS1 [102]. These differences strongly suggest that the eutherian lineage experienced a substantially higher frequency of viral gene domestication.
The “placenta-driven evolution” hypothesis further highlights the importance of tissues characterized by reduced DNA methylation, because leaky transcription in such environments enables viral sequences to be expressed, evaluated, and potentially co-opted into functional roles in a relatively safe environment. Recent studies indicate that, unlike eutherians, marsupials do not undergo global DNA demethylation during preimplantation development [131]. Although marsupial placental tissues display lower methylation than embryonic tissues, the degree of hypomethylation is modest compared with that of eutherians [131]. This developmental difference may have further contributed to the disparity in viral gene acquisition between the two lineages. Taken together, these considerations suggest that the extended duration and epigenetic permissiveness of the eutherian placenta created a uniquely favorable environment for the recurrent acquisition and domestication of virus-derived genes. Consequently, viral gene domestication may have been tightly intertwined with the evolutionary diversification of placental form and function across therian and, more specifically, eutherian lineages.
4. Conclusions
The placenta is an evolutionarily novel organ that emerged in therian mammals. Although more than a thousand genes are thought to contribute, directly or indirectly, to placental development and function, a small number of metavirus- and retrovirus-derived genes have made disproportionately important contributions to this evolutionary innovation. Among these, PEG10, the first domesticated gene acquired in the common therian ancestor, plays multiple critical roles in placental formation and maintenance. In contrast, PEG11/RTL1, which was domesticated only in the eutherian lineage, is essential for the maintenance of placental internal vasculature, thereby enabling the prolonged gestation characteristic of eutherian mammals. SIRH7/LDOC1, another eutherian-specific gene, plays important roles of trophoblast differentiation and maturation, thereby facilitating the endocrine functions of the placenta during gestation. Together, these observations indicate that metaviral gene domestication played a central role in the emergence and subsequent elaboration of the placenta. In addition, two syncytin genes, domesticated from retroviruses in a lineage-specific manner within therians, play essential roles in the formation of syncytial layers surrounding fetal capillaries in the placenta.
These virus-derived genes were likely domesticated through a two-step evolutionary process: first, their retention by genetic drift (nearly neutral evolution) under conditions of relaxed epigenetic repression, such as low DNA methylation in extraembryonic tissues including the placenta and yolk sac, and second, their subsequent functional refinement by Darwinian natural selection.
Following the establishment of the placenta through the domestication of PEG10, the number of virus-derived genes increased markedly in therian lineages. This expansion was particularly pronounced in eutherians with prolonged gestation, exemplified by the domestication of PEG11/RTL1, in contrast to marsupials, which retain a short gestation period. Notably, all ten eutherian-specific SIRH/RTL genes examined to date play essential roles either in the placenta or in microglia, which originate from the yolk sac during early development. Together, these patterns imply that the placenta and yolk sac—and possibly other extraembryonic tissues—have acted as engines of mammalian evolution by facilitating the recurrent acquisition and functional integration of virus-derived genes. This evolutionary process can be conceptualized as placenta-driven evolution.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Rossant J. Cross J.C. Lineage Specification and Differentiation: Extraembryonic Lineages Mouse Development: Patterning, Morphogenesis and Organogenesis Tam P.L. Rossant J. Academic Press Cambridge, MA, USA 2001155174
- 2Rossant J. Cross J.C. Placental development: Lessons from mouse mutants Nat. Rev. Genet.2001253854810.1038/3508057011433360 · doi ↗ · pubmed ↗
- 3Freyer C. Renfree M.B. The mammalian yolk sac placenta J. Exp. Zool. B. Mol. Dev. Evol.200931254555410.1002/jez.b.2123918985616 · doi ↗ · pubmed ↗
- 4Renfree M.B. Marsupials: Placental mammals with a difference Placenta 201031 S 21S 2610.1016/j.placenta.2009.12.02320079531 · doi ↗ · pubmed ↗
- 5Nowak-Sliwinska P. Segura T. Iruela-Arispe M.L. The chicken chorioallantoic membrane model in biology, medicine and bioengineering Angiogenesis 20141777980410.1007/s 10456-014-9440-725138280 PMC 4583126 · doi ↗ · pubmed ↗
- 6Chen L. Wang S. Feng Y. Zhang J. Du Y. Zhang J. Ongeval C.V. Ni Y. Li Y. Utilisation of Chick Embryo Chorioallantoic Membrane as a Model Platform for Imaging-Navigated Biomedical Research Cells 20211046310.3390/cells 1002046333671534 PMC 7926796 · doi ↗ · pubmed ↗
- 7Li Z. Pollack G.H. On the driver of blood circulation beyond the heart P Lo S ONE 202318 e 028965210.1371/journal.pone.028965237856567 PMC 10586597 · doi ↗ · pubmed ↗
- 8Tyndale-Biscoe C.H. Renfree M.B. Reproductive Physiology of Marsupials Cambridge University Press Cambridge, UK 1987295, 310314