Microbiota-Derived Proteins Shape T Cell Responses in Healthy and Colorectal Cancer Subjects
Elena Niccolai, Giulia Nannini, Serena Martinelli, Valentina Puca, Viviana De Luca, Laura Fortuna, Fabio Cianchi, Simone Carradori, Clemente Capasso, Rossella Grande, Amedeo Amedei

TL;DR
This study shows how proteins from gut bacteria affect T cell responses differently in healthy people and colorectal cancer patients.
Contribution
The study reveals distinct immunomodulatory effects of Fusobacterium nucleatum and Akkermansia muciniphila proteins on T cell subsets in healthy and CRC subjects.
Findings
F. nucleatum increases Th0, Th2, and Tc9 cells while reducing Th1 and Treg cells in healthy donors.
A. muciniphila promotes pro-inflammatory T cell subsets like Th0, Th2, Th17, and Tc17 in healthy individuals.
Bacterial extracts caused no significant T cell changes in colorectal cancer patients.
Abstract
Background/Objectives: Fusobacterium nucleatum and Akkermansia muciniphila are key components of the human microbiota, influencing health and disease. F. nucleatum is associated with colorectal cancer (CRC) and poor prognosis through its pro-inflammatory and pro-tumorigenic activity, whereas A. muciniphila is linked to metabolic benefits and anti-inflammatory effects. This study aimed to evaluate the immunomodulatory impact of protein extracts from these bacteria on peripheral T cell responses in healthy individuals and CRC patients. Methods: Peripheral blood mononuclear cells (PBMCs) were exposed to bacterial extracts, individually or in combination, and T cell subsets were analyzed by polychromatic flow cytometry. Results: In healthy donors, F. nucleatum increased Th0, Th2, and Tc9 cell frequencies while reducing Th1, Th1/Th17, and Treg cells. Conversely, A. muciniphila promoted a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
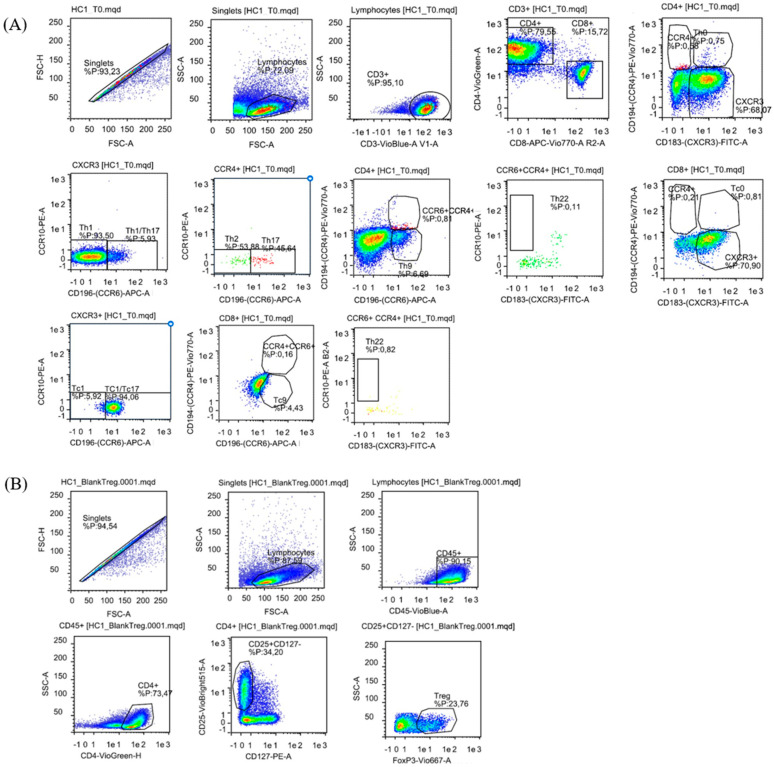 Figure 1
Figure 1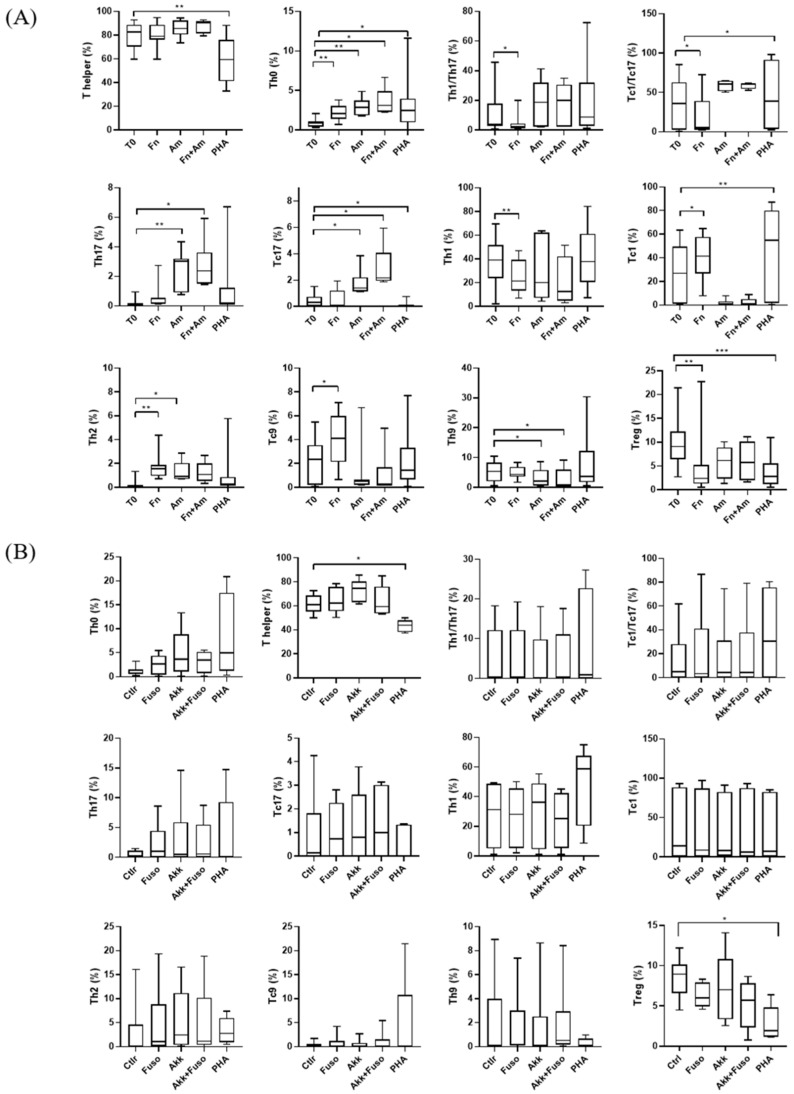 Figure 2
Figure 2- —FONZIE project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Probiotics and Fermented Foods · Cancer Research and Treatments
1. Introduction
The microbiome plays a crucial role in human health by aiding in nutrient bioconversion, protecting against pathogens, producing signaling molecules, and especially modulating immune responses. Under physiological conditions, interactions between the immune system and microbiota ensure the right coordination of innate and adaptive immune responses, enabling the most appropriate reactions [1]. However, dysbiosis, or an imbalance in the composition of the microbiota, could lead to immune dysregulation or increased susceptibility to diseases, including cancer [2].
In the large intestine, the microbiome role is relevant for maintaining mucosal and systemic immune homeostasis [3]. It interacts closely with local immune cells to modulate immune responses and exert immunomodulatory functions [1,4]. The microbiome shapes both the myeloid and lymphoid arms of the innate immune system [5], particularly through its role in developing B and T lymphocytes’ repertoire. Naïve CD4^+^ T helper (Th) cells differentiate into distinct subtypes—follicular helper T (Tfh), Th1, Th2, Th17, Th9, Th22, and regulatory T cells (Tregs)—in response to different stimuli, playing a critical role in immune regulation [6]. Maintaining the proper balance between T cell subsets is critical for immune homeostasis, with disruptions in Th1, Th17, or Tregs function linked to conditions like autoimmune diseases and cancer [7,8,9].
In colorectal cancer (CRC), the tumor microenvironment is often characterized by an imbalance in T cell subpopulations, such as Th1, Th17, and Tregs, which can influence disease progression [7]. For example, Tregs may suppress anti-tumor immunity, while Th1 cells, which are typically anti-tumorigenic, are often downregulated in the tumor microenvironment [7,10,11]. Additionally, Th17 cells can play dual roles in cancer by promoting chronic inflammation and tumor growth or, conversely, by recruiting immune cells to the tumor site [12,13,14,15,16].
Recent studies have further highlighted the complexity of microbiota–immune interactions in cancer and metabolic diseases, stressing the highly context-dependent immunomodulatory roles of specific bacterial species and their components [17,18].
Various gastric and intestinal microbial species, such as Helicobacter spp., Akkermansia muciniphila, Bacteroides thetaiotaomicron, and Bacteroides fragilis, can influence the differentiation of T cells, for example through their production of short-chain fatty acids (SCFA), contributing to the balance between effector and suppressor arms of the adaptive immune system [19,20]. Among the microbial species influencing immune responses, Fusobacterium nucleatum (F. nucleatum) and Akkermansia muciniphila (A. muciniphila) stand out. F. nucleatum is a commensal anaerobic Gram-negative bacterium of the oral microbiota. The spread of F. nucleatum to distant sites may drive extra-oral infections, systemic inflammation, and cancer [21,22,23]. Genomic analyses have revealed that F. nucleatum is enriched in human CRC tissues and linked to poor patient outcomes [24,25]. It has been shown to suppress T cell-mediated anti-tumor immunity, leading to a more immunosuppressive tumor environment [25,26,27,28,29]. Conversely, A. muciniphila is another Gram-negative anaerobic bacterium of intestinal mucus with recognized probiotic properties [30]. It is known to increase the integrity of the intestinal barrier [31] and shows beneficial effects on metabolic health [32,33,34]. A. muciniphila modulates immune responses by influencing T cell activation and differentiation, promoting an anti-inflammatory environment under homeostatic conditions [6]. While its role in CRC is less clear, some studies suggest a higher abundance of A. muciniphila in CRC patients, while others report lower levels in advanced disease stages [35,36,37,38].
The primary aim of this study was to investigate the immunomodulatory effects of protein extracts from F. nucleatum and A. muciniphila on peripheral blood T cells from healthy individuals. We sought to determine whether these bacterial extracts had the ability to skew T cell subset frequencies, thereby revealing potential probiotic or pro-inflammatory properties. In a subsequent analysis, we tested the same extracts on peripheral blood T cells from CRC patients to assess if their immunomodulatory effects were similar or altered in a pathological scenario.
Although the experimental approach is based on prolonged in vitro exposure to bacterial protein extracts, the documented changes in T cell subset distribution mirror the immunomodulatory potential of these extracts in shaping T cell polarization.
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
The bacterial reference strains used in this study were Fusobacterium nucleatum ATCC 25586 and Akkermansia muciniphila BAA-835. The strains purchased from ATCC were stored at −80 °C in glycerol stocks. Both bacteria were thawed at room temperature, plated on Fastidious Anaerobe Agar (FAA) (Lab M, Heywood, UK) plus 5% of defibrinated sterile horse blood (Oxoid Limited, Hampshire, UK), and incubated at 37 °C for 72 h in an anaerobic atmosphere (Whitley A25 Anaerobic workstation, Don Whitley Scientific Limited, West Yorkshire, UK). After the incubation time, F. nucleatum bacterial colonies on agar were collected and inoculated in Fastidious Anaerobe Broth (FAB) (Lab M, Heywood, UK) for 17 h at 125 rpm at 37 °C [39,40] whereas A. muciniphila bacterial colonies were collected and inoculated in Brain Heart Infusion Broth (BHIB; Oxoid Limited, Hampshire, UK) plus 0.3 mg/mL L-cysteine for 48 h at 125 rpm at 37 °C. After each incubation time, F. nucleatum and A. muciniphila broth cultures were centrifuged at 3200× g at 4 °C for 20 min and washed twice in sterile 0.01 M phosphate buffered saline (PBS) and centrifuged again to obtain a visible pellet corresponding to 3.7 × 10^10^ CFU and 6.41 × 10^9^ CFU, respectively.
2.2. Bacterial Extracts
The bacterial pellets were resuspended in 20 mM Tris-HCl buffer, pH 8.3, transferred to a 15 mL conical tube, placed in an ice bath, and disrupted by sonication. The sonicator (Sonoplus, Bandelin, Berlin, Germany) was set to an amplitude of 50% with a pulse sequence of 10 s ON and 120 s OFF. The bacterial suspension was sonicated for a total of 10 cycles, ensuring that the sample remained on ice throughout to maintain a low temperature. The progress of cell lysis was monitored periodically by examining the turbidity of the suspension. Following sonication, the lysate was centrifuged at 11,200× g for 15 min at 4 °C to separate the cell debris. The supernatant, containing the soluble bacterial proteins, was carefully collected and transferred to a new tube. The protein concentration of the bacterial extract was quantified using the Bradford protein assay according to the manufacturer’s protocol [41]. The resulting protein concentrations were 1.35 mg/mL for A. municiphila and 0.8 mg/mL for F. nucleatum. For all functional assays, bacterial extracts were standardized and used based on total protein concentration (µg/mL), allowing direct comparison between F. nucleatum and A. muciniphila independently of differences in bacterial growth or CFU counts. Aliquots of bacterial protein extracts were stored at −80 °C and thawed only once prior to use to avoid protein degradation and batch variability.
2.3. Study Population and Sample Collection
Peripheral blood mononuclear cells (PBMCs) were obtained from healthy control (HC) volunteers and sex- and age-matched CRC patients who were already enrolled in a previous study approved by the local Ethics Committee. Blood samples from CRC patients were collected upon their hospitalization, specifically on the day before their scheduled cancer resection. Notably, none of the CRC patients had received chemotherapy prior to blood collection, ensuring that the samples reflected a chemotherapy-naive state. For each subject, 15 mL of blood was collected in K2 EDTA collection tubes. The PBMCs were isolated using Lymphoprep^TM^ (Serumwerk, Bernburg, Germany) density gradient centrifugation and immediately used for culture experiments.
2.4. Bacterial Extract-Conditioned T Cell Cultures
To evaluate bacterial extract-driven modulation of T cell polarization, PBMCs from each subject were cultured in the presence of F. nucleatum and A. muciniphila protein extracts. 5 × 10^5^ PBMCs from each subject were cultured in 2 mL of RPMI 1640 medium enriched with 2 mM L-glutamine, 2 × 10^5^ M 2-mercaptoethanol (2-ME), and 7.5% human serum (complete medium) in 24-well plates. On day 0, PBMCs were stimulated once with bacterial extracts at different concentrations (10, 15, 20, and 50 µg/mL). The culture medium was not replaced during the first 5 days, allowing sustained exposure to bacterial protein extracts. As a positive control for cell reactivity, the cells were also stimulated with phytohemagglutinin (PHA) at a concentration of 0.1%. On the fifth day of culture, recombinant IL-2 (rIL-2) was added at 30 U/mL to support T cell expansion, and cultures were maintained for an additional 7 days. At the end of the culture period (day 12), cells were harvested and T cell subset frequencies were analyzed by polychromatic flow cytometry. This experimental protocol was designed to assess the long-term immunomodulatory effects of bacterial protein extracts on T cell polarization rather than short-term activation responses.
2.5. Cytometric Analysis of Surface and Intracellular Markers of T Lymphocytes
The cultured cells were stained with fluorochrome-conjugated antibodies at baseline (T0) and after 12 days of culture with bacterial extracts or PHA. Specifically, the following anti-human antibodies from Miltenyi Biotech were used: CD45-PercP Vio700, CD3-VioBlue, CD8-APC Vio770, CD4-VioGreen, CCR10-PE, CD183-VioBright FITC, CD194-PE Vio770, and CD196-APC. To define the various T subpopulations, a phenotype based on CD45, CD3, CD4, and CD8 staining has been combined with CD183 (CXCR3) for Th1 and Tc1, CD194 (CCR4) for Th2 and Tc2, CD196 (CCR6) for Th17 and Tc17 or Th9 and Tc9; CCR10 for Th22 [42]. The gating strategy is shown in Figure 1. Additionally, cultured cells were used to detect Tregs lymphocytes employing the Tregs detection kit (Miltenyi Biotec, Bergisch Gladbach, Germany), which included external staining with anti-human CD45-VioBlue, CD4-VioGreen, CD25-VioBright, CD127-PE antibodies, cell fixation and permeabilization, followed by intracellular staining with anti-human FoxP3-Vio667 (Figure 1B). The analysis was performed using the MacsQuant Analyzer flow cytometer (Miltenyi Biotec), with the acquisition of 100,000 events. Data were processed with FlowLogic V7 software (Miltenyi Biotec, Germany).
2.6. Statistical Analysis
Statistical analyses were performed by comparing post-culture T cell subset frequencies with baseline (T0) values in order to evaluate bacterial extract-driven modulation of T cell polarization relative to the initial immune profile of each subject. Data analyses were performed using GraphPad Prism 8.0. Continuous variables were presented as median value and interquartile range (IQR) and were compared using the Kruskal–Wallis test (Dunnett post-test). p-values less than 0.05 were considered statistically significant.
3. Results
3.1. Study Population
The median age of healthy volunteers (n = 15; 7 males, 8 females) was 67 years (49–79), and for CRC patients (n = 6; 3 females, 3 males), it was 75 years (66–80), with gender distribution balanced at 50% female in both groups. The clinical/demographical characteristics of CRC patients are reported in Table 1.
3.2. Effects of Bacterial Extracts on HC T Lymphocytes
To assess bacterial extract-driven modulation of T cell polarization relative to baseline immune conditions, T cell subset frequencies were compared between T0 and day 12 cultures. We initially tested increasing concentrations of bacterial extracts (10, 15, 20, and 50 µg/mL). Based on preliminary results, we selected 15 µg/mL as the optimal concentration for further experiments, as it was the adequate amount to observe significant effects, and lower/higher concentrations did not produce significant changes.
The presence of F. nucleatum induced significant changes in various T cell subpopulations compared to the baseline, including increased frequencies of Th0 cells (p = 0.005), Th2 cells (p = 0.002), and Tc9 cells (p = 0.034), and decreased frequencies of Th1 cells (p = 0.002), Th1/Th17 cells (p = 0.015), Tc1/Tc17 cells (p = 0.048), and Tregs cells (p = 0.005) (Figure 2). The median and interquartile range for all the T cell subsets are reported in Table 2.
On the contrary, the presence of A. muciniphila resulted in a significant enrichment of Th0 cells (p = 0.005), Th17 cells (p = 0.005), Th2 cells (p = 0.017), and Tc17 cells (p = 0.038) compared to the baseline (Figure 2).
Overall, these findings highlight that protein extracts from F. nucleatum and A. muciniphila clearly induce distinct and species-specific patterns of T cell subset modulation in healthy individuals, with partially opposing effects on key T helper and cytotoxic-associated populations.
Finally, the treatment with a combination of F. nucleatum and A. muciniphila extracts led to notable changes in T cells’ frequencies, including increased amounts of Th0 cells (p = 0.021), Th17 cells (p = 0.018), and Tc17 cells (p = 0.043), and decreased frequencies of Th9 cells (p = 0.047) (Figure 2).
3.3. Impact of Bacterial Extracts on T Cell Subsets’ Distribution CRC Patient
Treatment with F. nucleatum and A. muciniphila extracts, both alone and in combination, did not result in significant changes in the frequencies of T lymphocytes obtained from CRC patients compared to the baseline. The median and interquartile range for all the T cell populations are reported in Table 2.
4. Discussion
As previously reported, the primary aim of our study was to evaluate Fusobacterium nucleatum- and Akkermansia muciniphila-derived bacterial extract-driven modulation of systemic T cell polarization relative to the baseline immune profile of healthy subjects. We used PBMCs to assess how these bacterial extracts modulate circulating T cell subsets in a non-diseased context. Subsequently, the same extracts were tested on PBMCs from CRC patients to document if the immunomodulatory effects observed in healthy condition were preserved under pathological scenario such as cancer. PBMCs were cultured with bacterial protein extracts to allow sustained antigen exposure and subsequent IL-2–supported expansion, and immunological outcomes were interpreted as changes in T cell subset distribution relative to baseline (T0), which represents the physiological immune state of each donor [43]. The use of polychromatic flow cytometry enabled us to evaluate the distribution of specific T cell subsets. In this study, T cell subsets were defined phenotypically based on surface marker expression. While these markers are widely used to associate T cell lineages with specific effector programs, functional cytokine production was not directly assessed, and therefore functional activity cannot be directly inferred. CD4^+^ Th and CD8^+^ Tc cells differentiate into distinct subtypes, characterized by specific transcription factors and cytokine profiles, such as helper or cytotoxic Th1/Tc1, Th2/Tc2, Th17/Tc17, Th9/Tc9, Th22/Tc22, and regulatory T cells (Tregs), when properly stimulated [6]. However, this differentiation is not static: T cells can reshape into other subtypes based on physiological/pathological conditions. For example, Th17 cells can shift to a Th17/Th1 profile under high IL-12 levels, while IL-1 and IL-6 can prompt Tregs to convert into Th17 cells [44,45,46].
Th1 and Th17 cells act as effector cells in immune responses, while Tregs regulate and suppress immune activity. Imbalances favoring chronic Th17-mediated inflammation and decreasing Tregs function are implicated in autoimmune diseases [47]. Additionally, Th1 cells are crucial for cell-mediated immunity against intracellular pathogens, whereas Th2 cells support humoral immunity against extracellular parasites. Maintaining a rightly balanced Th1/Th2 response is essential for immune homeostasis, while disruptions can contribute to conditions such as allergies, asthma, and autoimmune disorders [48,49]. It is notable that we used protein fraction extracts of F. nucleatum and A. muciniphila to specifically evaluate their immunomodulatory effects.
Protein components of bacterial cells are potent antigens that can directly activate T cells, making them ideal for assessing targeted immune responses. By focusing on the protein fraction, we aimed to diminish the complexity associated with whole bacteria or non-protein components, such as lipopolysaccharides or extracellular polymeric substances, and to better characterize bacterial extract-driven modulation of T cell responses. While bacterial metabolites and other bioactive molecules are known to influence immune responses [50,51], potentially skewing the immunity towards inflammatory or regulatory profiles [52,53], the use of protein fractions was finalized to control for such variables and focus specifically on antigen-driven T cell activation.
In healthy individuals, exposure to F. nucleatum protein extracts was associated with changes in T cell subset distribution, characterized by elevated frequencies of Th0, Th2, Tc1, and Tc9 cells, alongside decreased frequencies of Th1, Th1/Th17, and Tregs cells. This suggests that F. nucleatum promotes both humoral immunity and mucosal defenses through the rise of Th2 and Tc9 subsets, while simultaneously enhancing cytotoxic responses via increased Tc1 cells [6]. However, the decrease in Th1 and Tregs may decrease regulatory and pro-inflammatory control, potentially creating a balance that could, if unregulated, contribute to immunosuppressive conditions. This altered immune profile may facilitate cancer progression by promoting chronic inflammation while weakening anti-tumor responses [54,55]. The enrichment of Th2- and Tc9-associated phenotypes is more relevant in CRC-associated immune dysregulation, as Th2-skewed responses have been linked to impaired anti-tumor immunity and immune tolerance, while Tc9 cells have been associated with immune environments characterized by limited cytotoxic efficiency and functional plasticity [56,57].
A key finding of this study is the contrasting and species-specific nature of bacterial extract-driven T cell polarization induced by F. nucleatum and A. muciniphila in healthy subjects, underscoring the heterogeneous immunomodulatory potential of individual microbiota members.
Exposure to A. muciniphila was associated with T cell polarization toward subsets frequently linked to pro-inflammatory immune responses, characterized by increased Th0, Th2, Th17, and Tc17 cell frequencies. Th17 and Tc17 cells are known for their production of IL-17, which is involved in mediating inflammation and recruiting neutrophils [58]. While beneficial in pathogen defense, this subset’s involvement in chronic inflammation can favor some diseases, such as CRC, where chronic inflammatory responses can promote tumorigenesis [59]. The apparent discrepancy between the pro-inflammatory-associated T cell phenotype observed in vitro following exposure to A. muciniphila protein extracts and the anti-inflammatory effects reported for A. muciniphila in vivo likely mirrors differences in experimental condition. In vivo, the immunomodulatory properties of A. muciniphila are influenced by microbial localization, host–microbiota interactions, and the production of metabolites that are missing in an in vitro PBMC-based system. Moreover, tissue-specific immune environments and epithelial–immune crosstalk may critically shape the clear immunological outcome. Therefore, the bacterial extract–driven modulation observed in vitro should be interpreted as a context-dependent immune polarization rather than a direct contradiction of A. muciniphila’s reported anti-inflammatory roles in vivo.
The combination of F. nucleatum and A. muciniphila extracts led to an increase in Th0, Th17, and Tc17 cells, along with a decrease in Th9 cells, suggesting a shift towards T cell subset distributions commonly associated with pro-inflammatory immune polarization and mucosal immune responses. However, the enrichment of T cell subsets usually associated with pro-inflammatory responses could, if not properly modulated, contribute to chronic inflammatory conditions. One interesting finding is that A. muciniphila does not increase Tc1 cells on its own, but when combined with F. nucleatum, it mitigates the F. nucleatum-induced increase in Tc1 cells. This finding suggests a potential regulatory role for A. muciniphila in counteracting the pro-inflammatory Tc1 response driven by F. nucleatum. The biological relevance of combined exposure experiments should be evaluated in the complexity of the dynamic gut microbiota ecosystem, where host immune cells are simultaneously exposed to multiple microbial components rather than single bacterial species. Although the in vitro simultaneous exposure does not adequately mirror the spatial and temporal heterogeneity of microbial interactions in vivo, this approach provides a simplified and controlled model to explore how different bacterial components may interactively shape T cell polarization. Therefore, the combined extract experiments offer insight into potential additive or modulatory effects that may emerge from multi-microbial exposure within the intestinal environment.
The mechanisms behind this modulation could involve complex interactions in the cytokines’ environment or antigenic signaling. Surely, further investigation is needed to understand why A. muciniphila specifically affects Tc1 cells in this scenario, while other subsets, like Th1, remain unchanged. Notably, the combination of bacterial extracts seemed to influence specific subsets, pointing to complex interactions between these microbiota members in modulating host immunity.
In the blood of CRC patients, stimulation with bacterial extracts did not obtain significant changes in circulating T cell subset frequencies compared to baseline; however, given the limited sample size and the cohort heterogeneity, the study may be underpowered to detect subtle immunomodulatory effects. This lack of detectable modulation in peripheral T cell subsets is consistent with the concept of systemic immune dysfunction in colorectal cancer, which may limit the responsiveness of circulating T cells to additional immunomodulatory stimuli. In CRC patients, the lack of statistically significant modulation in circulating T cell subsets may mirror multiple, non-mutually exclusive mechanisms. In addition to potential sequestration of antigen-responsive T cells within the tumor microenvironment or mucosal tissues, peripheral T cells may exhibit functional impairment, exhaustion, or anergic condition, as commonly documented in colorectal cancer [60]. The immunosuppressive CRC microenvironment, characterized by Tregs and myeloid-derived suppressor cells, may further obscure the detection of immune shifts in peripheral blood [61]. Future studies should focus on tumor biopsies to directly assess local immune dynamics.
This study has some limitations. The analysis of T cell subsets was restricted, and we did not perform functional assays, limiting our understanding of the broader immune implications. In addition, potential confounding factors related to the CRC cohort should be considered. Although all patients were chemotherapy naïve, disease stage heterogeneity and cancer-associated systemic immune alterations, including immune exhaustion or anergy, may have influenced peripheral T cell responses. Additionally, the limited number of CRC patients and the associated variability represent an additional limitation of the study and may have decreased the statistical power to detect significant differences. Future research should involve chronic treatment models, broader immune profiling, and functional assays to provide a finer understanding. It will also be relevant to study the immunomodulatory effects of these bacterial extracts on tumor-infiltrating lymphocytes, as local immune responses may differ significantly from those observed in peripheral blood.
Finally, exploring the microbiota in tumor biopsies and fecal samples from the same patients would help establish potential clinical correlations, deepening our understanding of the interactions between microbiota and immune responses systemically and locally.
5. Conclusions
In conclusion, our findings highlight F. nucleatum- and A. muciniphila-driven modulation of T cell subsets associated with pro-inflammatory or regulatory immune profiles.
We document that protein extracts derived from distinct gut microbiota members differentially impact circulating immune cells in healthy subjects, whereas no statistically significant modulation was detected in CRC patients under the same experimental conditions. Understanding microbiota–immune interactions is critical for the development of probiotic or microbial-based strategies aimed at modulating immune responses, with potential implications for infections, autoimmune diseases, and lastly, cancer. Our findings underscore the relevance of considering both microbial specificity and host immune context when investigating systemic immune modulation.
In sum, this study highlights four key take-home messages:
- (i)Bacterial protein extracts from distinct gut microbiota members differentially modulate circulating T cell subset distribution in healthy subjects;
- (ii)Fusobacterium nucleatum and Akkermansia muciniphila exert distinct and species-specific immunomodulatory effects on systemic T cell polarization;
- (iii)In CRC patients, systemic T cell modulation is not readily detectable under the present experimental conditions, likely mirroring disease-associated immune dysregulation and/or limited statistical power;
- (iv)These findings stress the relevance of considering both microbial specificity and host immune context when studying microbiota–immune interactions.
Further studies, including functional assays and larger, well-powered patient cohorts, will be required to better define the role of microbiota-derived components in shaping systemic and CRC-associated immune responses.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zitvogel L. Ayyoub M. Routy B. Kroemer G. Microbiome and Anticancer Immunosurveillance Cell 201616527628710.1016/j.cell.2016.03.00127058662 · doi ↗ · pubmed ↗
- 2Niccolai E. Boem F. Emmi G. Amedei A. The link “Cancer and autoimmune diseases” in the light of microbiota: Evidence of a potential culprit Immunol. Lett.2020222122810.1016/j.imlet.2020.03.00132145242 · doi ↗ · pubmed ↗
- 3Turner J.R. Intestinal mucosal barrier function in health and disease Nat. Rev. Immunol.2009979980910.1038/nri 265319855405 · doi ↗ · pubmed ↗
- 4Russell M.W. Ogra P.L. Mucosal decisions: Tolerance and responsiveness at mucosal surfaces Immunol. Investig.20103929730210.3109/0882013100372992720450281 · doi ↗ · pubmed ↗
- 5von Moltke J. Ji M. Liang H.E. Locksley R.M. Tuft-cell-derived IL-25 regulates an intestinal ILC 2-epithelial response circuit Nature 201652922122510.1038/nature 1616126675736 PMC 4830391 · doi ↗ · pubmed ↗
- 6Sun L. Su Y. Jiao A. Wang X. Zhang B. T cells in health and disease Signal Transduct. Target. Ther.2023823510.1038/s 41392-023-01471-y 37332039 PMC 10277291 · doi ↗ · pubmed ↗
- 7Thommen D.S. Schumacher T.N. T Cell Dysfunction in Cancer Cancer Cell 20183354756210.1016/j.ccell.2018.03.01229634943 PMC 7116508 · doi ↗ · pubmed ↗
- 8Niccolai E. Ricci F. Russo E. Nannini G. Emmi G. Taddei A. Ringressi M.N. Melli F. Miloeva M. Cianchi F. The Different Functional Distribution of “Not Effector” T Cells (Treg/Tnull) in Colorectal Cancer Front. Immunol.20178190010.3389/fimmu.2017.0190029375559 PMC 5770731 · doi ↗ · pubmed ↗