Stationary-phase Pseudomonas aeruginosa fluoroquinolone persisters mostly avoid DNA double-stranded breaks
Patricia J. Hare, Juliet R. González, Wendy W. K. Mok

TL;DR
Some Pseudomonas aeruginosa cells survive fluoroquinolone antibiotics without DNA damage, challenging current persistence models.
Contribution
Most stationary-phase P. aeruginosa persisters avoid DNA double-stranded breaks rather than relying on repair mechanisms.
Findings
The majority of P. aeruginosa fluoroquinolone persisters do not show DNA double-stranded breaks.
Persisters with DNA damage take longer to divide and may repair or sequester the damage.
These findings suggest new mechanisms for antibiotic persistence in P. aeruginosa.
Abstract
When susceptible bacterial cultures are treated with antibiotics, some cells can survive treatment without heritable resistance, giving rise to susceptible daughter cells in a phenomenon termed antibiotic persistence. Current models of fluoroquinolone (FQ) persistence in stationary-phase cultures posit that post-treatment resuscitation is dependent on double-stranded break (DSB) repair through RecA-mediated homology-directed repair. Previously, we reported that stationary-phase P. aeruginosa does not depend on RecA to persist. In this work, we ask whether P. aeruginosa FQ persisters from stationary-phase cultures suffer DSBs at all. We measured DSB formation in Levofloxacin (LVX)-treated cells recovering from treatment using strains expressing fluorescently labeled DSB-binding protein, Gam. We find that, surprisingly, the majority of P. aeruginosa LVX persisters survive treatment…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
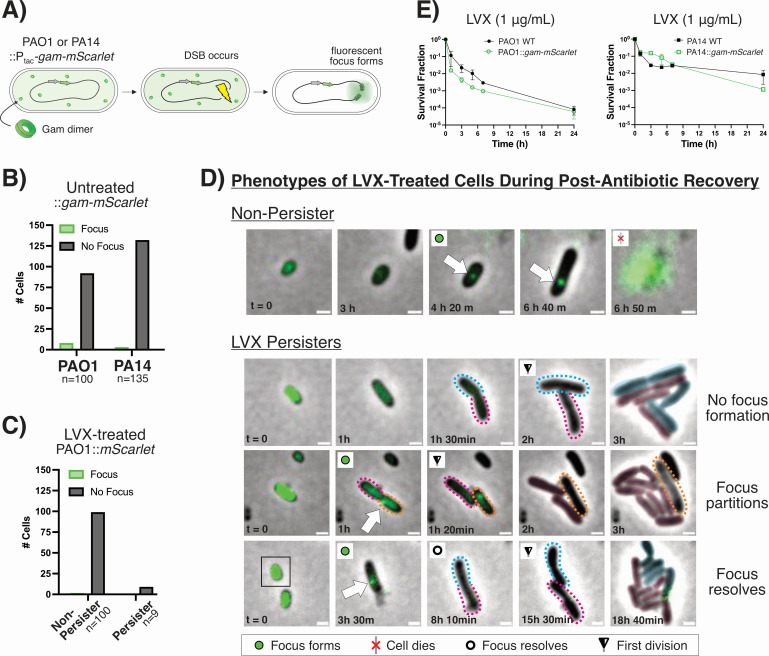 Fig 1
Fig 1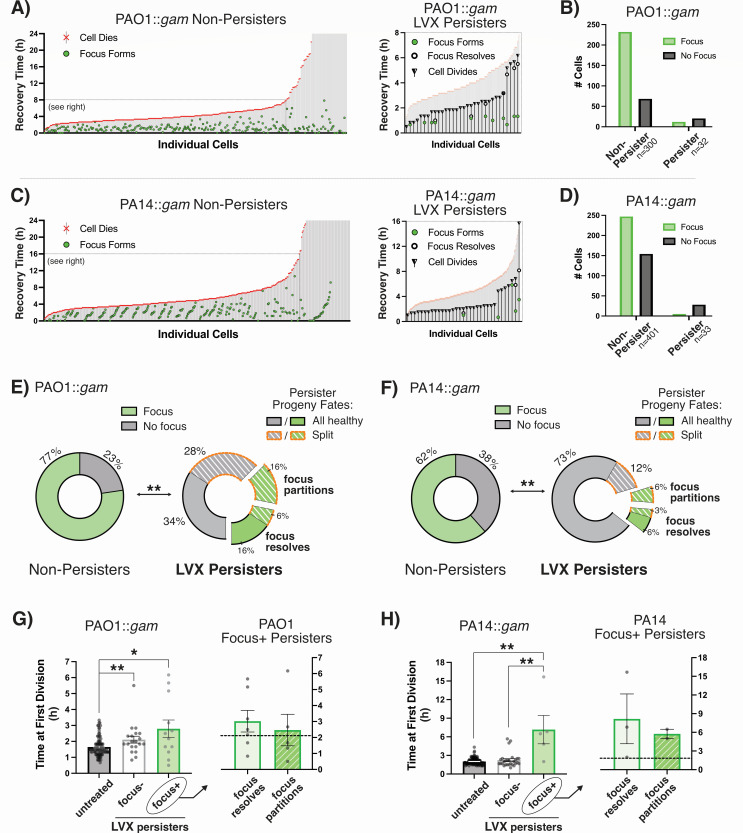 Fig 2
Fig 2- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —National Institute of Dental and Craniofacial Researchhttp://dx.doi.org/10.13039/100000072
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Bacterial Genetics and Biotechnology · DNA Repair Mechanisms
OBSERVATION
The fluoroquinolones (FQs) are a synthetic class of antibiotics that trap bacterial topoisomerases on DNA, leading to mounting topological stress and DNA double-stranded breaks (DSBs) (1, 2). Even in clonal laboratory cultures of FQ-susceptible bacterial populations, some cells can survive treatment in the phenomenon known as persistence. After treatment, persisters can lead to population resurgence and treatment failure (3, 4). Studies on FQ antibiotic persistence in stationary-phase Escherichia coli, Staphylococcus aureus, and Salmonella spp. have elucidated some commonalities across all species: (i) the majority of cells that die do so in the post-antibiotic period, not during treatment itself, and (ii) persisters engage in RecA-mediated DSB repair after FQ treatment to survive (5–8).
We previously found that FQ-treated stationary-phase Pseudomonas aeruginosa does not adhere to the first paradigm and dies readily during exposure to the FQ, Levofloxacin (LVX) (9). We also found that P. aeruginosa does not require RecA to persist after LVX treatment in our experimental conditions (9, 10). Therefore, we hypothesized that P. aeruginosa also subverts the second tenet of FQ persistence and that this gram-negative species does not engage in DNA repair in order to persist.
To test this hypothesis, we modified a Gam reporter construct that has previously been utilized for labeling DSBs in E. coli and eukaryotic cell cultures (11, 12). We expressed IPTG-inducible Gam-mScarlet from the P. aeruginosa chromosome as cells were grown to the stationary phase in basal salt medium (BSM). When DSBs occur, the fluorescently labeled Gam reporter binds to and accumulates on the dsDNA ends (Fig. 1A). This localization is visualized as a fluorescent focus. We confirmed that Gam-mScarlet binds to DSBs in P. aeruginosa by irradiating reporter strains with UV light, a stressor known to generate DSBs (11). We observed fluorescent foci in over 90% of UV-irradiated cells compared to fewer than 10% of unexposed cells (Fig. S1). Similarly, cells that have not been treated with antibiotics also rarely form foci (Fig. 1B). These data indicate that focus formation is not likely due to non-specific aggregation of Gam-mScarlet. We also treated cells bearing IPTG-inducible mScarlet with LVX and did not observe foci during recovery (Fig. 1C). From these controls, we conclude that focus formation is not attributable to non-specific mScarlet aggregation upon FQ treatment.
Fluorescently labeled Gam highlights DSBs in individual LVX-treated P. aeruginosa cells. (A) P. aeruginosa strains bearing IPTG-inducible Gam-mScarlet form fluorescent foci at sites of DSBs. mScarlet fluorescence is false-colored as green for clarity. Fluorescent foci do not form in (B) untreated gam-mScarlet strains or in (C) PAO1::Ptac-mScarlet—that lacks the Gam protein—during recovery from LVX treatment. (D) Representative images of P. aeruginosa gam-mScarlet cell fates. The colored, dotted outlines demarcate daughter cells originating from the same persister; the corresponding color masks in later frames indicate the lineage from which progenies were derived. The orange outline indicates a non-dividing daughter cell. Scale bars represent 1 µm. Representative videos of PAO1 and PA14 with each phenotype can be found in the supplemental material. (E) The survival of P. aeruginosa gam-mScarlet strains treated with 1 µg/mL LVX is not significantly different from wild-type strains (n = 2).
FQ-treated cell phenotypes are heterogeneous during recovery
To assess DSB formation in LVX-treated P. aeruginosa, cells were seeded onto nutritive agarose pads after antibiotic treatment and imaged during recovery. We observed focus formation in dead or non-dividing cells (“non-persisters”) and persisters alike (Fig. 1D). To verify that dividing cells represent persisters and not resistant mutants, we tested the LVX susceptibilities of cells that survived the initial LVX treatment (Fig. S2A). We found that post-LVX treatment cultures do not have increased LVX minimum inhibitory concentrations or proportions of LVX-tolerant cells, strongly suggesting that cells that survived LVX treatment are persistent, not resistant (Fig. S2B and C). Additionally, although Gam has been reported to obstruct the RecBCD nuclease and inhibit homologous recombination in E. coli, we did not find any differences in FQ persistence for P. aeruginosa strains with or without the fluorescent Gam construct (Fig. 1E) (13).
For persisters that formed foci, we observed that foci either partitioned into one of the daughter cells (“focus partitions”) or resolved (“focus resolves”) before division (Fig. 1D). We analyzed imaging data for individual cells over 24 h recovery and plotted the times of focus formation (if any), focus resolution, cell death, and first cell division (persisters only) (Fig. 2A and C). Analyses were carried out for a random sample of at least 100 non-persister cells for each experimental replicate and all the persister cells in the given fields of view.
*Imaging of P. aeruginosa gam-mScarlet illustrates that most LVX persisters avoid DSBs. The fates of individual P. aeruginosa::gam-mScarlet cells were tracked during recovery from LVX treatment (1 µg/mL) in (A) PAO1 and (C) PA14 (n ≥ 300 non-persisters over ≥ 2 biological replicates per strain). Cells that died or never divided after 24 h recovery were categorized as non-persisters. To the right, the fates of persisters are shown overlaid onto the non-persisters. (B and D) Aggregate data show that untreated cells and persisters rarely form fluorescent foci, but non-persisters do. (E and F) The ratios of fluorescent:non-fluorescent cells are significantly different between non-persisters and persisters for both strains (**P < 0.01; two-sided Fisher’s exact test). The pie charts show the fates of persisters that did or did not form foci and whether their progeny were all healthy (daughter cells successfully divide) or split (one daughter cell lysed or failed to divide). (G and H) The times of initial cell division are shown for each persister focus phenotype compared to untreated Gam-mScarlet strains. Statistical significance was assessed using unpaired, two-tailed Mann-Whitney tests with Welch’s correction for unequal variance (*P ≤ 0.05, *P < 0.01). The graph to the right shows the time of division for persisters that formed foci; the dashed line represents the average time to first division for non-focus-forming persisters.
The visual summaries of individual cell fates show that there is widespread post-antibiotic killing of P. aeruginosa treated with LVX (Fig. 2A and C). Most non-persisters died within the first 4.5 h (median times of death: 4.17 h for PAO1, 4.5 h for PA14). Of note, a significantly greater proportion of non-persister cells formed fluorescent foci during recovery than persister cells (Fig. 2B and D through F). This suggests that P. aeruginosa persisters mostly do not form DSBs when recovering from LVX treatment.
FQ persister progenies are a mix of viable and non-viable cells
We noticed that persisters either divided into healthy, proliferative daughter cells (“all healthy”) or the progeny were split between viable and non-dividing cells (“split”), regardless of whether a persister had Gam focus formation (14) (Fig. 2E and F). For the persisters that formed Gam foci, the likelihood that foci resolved or were partitioned into one of the daughter cells at the time of division was comparable. Of the PAO1 and PA14 persisters that partitioned foci, all seven of them gave rise to split progeny (Fig. 2E and F). Note that the daughter cells that retained foci failed to proliferate, suggesting that segregation of the Gam-labeled DSB into one daughter cell allowed the other to persist.
FQ persisters with DSBs have longer exit from lag
We observed that LVX persisters with foci formation took significantly longer to exit lag and divide than cells that were never treated with LVX (Fig. 2G and H). By comparison, the time of first division for foci-less persisters was similar to that of untreated cells. Persisters without foci divided approximately 40 min and 2 h earlier than the persisters with foci, respectively, for PAO1 and PA14. These data support a model in which persisters that avoid DSBs are able to resume growth quickly, like untreated stationary-phase cells, outpacing persisters that must cope with DSBs before dividing.
Discussion
In this study, we show that fluorescently tagged Gam can be used to track DSBs in P. aeruginosa and sheds light on the relevance of DNA breaks to stationary-phase FQ persistence in this species (10, 15, 16). Consistent with our previous data showing that RecA-mediated DSB repair is not necessary for stationary-phase P. aeruginosa FQ persisters grown in BSM, our microscopy data suggest that persisters are cells that avoid, rather than repair, DSBs (9). We observed that LVX persisters infrequently form fluorescent Gam foci (indicative of DSBs) en route to propagating after FQ treatment (Fig. 2A and C). It is possible that FQ treatment might generate DSBs with single-stranded overhangs that cannot be bound by Gam (12). However, the frequency of fluorescent foci in non-persister cells suggests that Gam-detectable breaks do occur during the post-antibiotic phase in the majority of FQ-treated cells: 77% and 62% for PAO1 and PA14 non-persisters, respectively (Fig. 2E and F). The infrequency of Gam foci in persister cells seems to support the traditional perspective that P. aeruginosa FQ persisters are cells with low metabolic activity—perhaps protected within biofilm-like aggregates—that are less susceptible to injury by antibiotics that target active cell processes (17, 18).
In keeping with this model, we found that persisters took more time to initially divide after treatment than untreated cells (Fig. 2G and H). Longer exit from lag is an established trait of metabolically quiescent cells that have increased persistence against antibiotics (19). Conversely, the delay may actually be a symptom, reflecting the time it takes for a persister to repair damage, liberate FQ-topoisomerase-DNA ternary complexes, or expel residual antibiotic before propagating (20, 21). We hypothesize that the mode of Gam focus resolution and fates of progeny indicate specific persistence mechanisms. For persisters whose fluorescent foci dissipated, we hypothesize that the longer delay until first division—compared to persisters without fluorescent foci—indicates the time it takes for break repair enzymes to displace Gam and complete repair. For persisters that partitioned foci into daughter cells, we hypothesize that those cells were effectively sorting FQ-trapped ternary complexes into their daughter cells, condemning some to die so that the others might propagate (20, 21). We expect that repair takes longer than ternary complex segregation and might explain those persisters’ slightly longer times until first division (Fig. 2G and H).
Collectively, our data suggest that P. aeruginosa FQ persisters do not fit the paradigms set by other pathogens. Furthermore, these data provide the impetus for further mechanistic studies of ternary complex segregation in FQ persister progeny. Understanding how individual cells overcome FQ treatment will inform strategies for fully eradicating susceptible populations, thereby limiting an infection’s ability to develop antibiotic resistance (4, 22, 23).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Drlica K, Zhao X. 1997. DNA gyrase, topoisomerase IV, and the 4-quinolones. Microbiol Mol Biol Rev 61:377–392. doi:10.1128/mmbr.61.3.377-392.19979293187 PMC 232616 · doi ↗ · pubmed ↗
- 2Kanj SS. 2024. Edited by KK Hall. Principles of antimicrobial therapy of Pseudomonas aeruginosa infections.
- 3Balaban NQ, Merrin J, Chait R, Kowalik L, Leibler S. 2004. Bacterial persistence as a phenotypic switch. Science 305:1622–1625. doi:10.1126/science.109939015308767 · doi ↗ · pubmed ↗
- 4Fauvart M, De Groote VN, Michiels J. 2011. Role of persister cells in chronic infections: clinical relevance and perspectives on anti-persister therapies. J Med Microbiol 60:699–709. doi:10.1099/jmm.0.030932-021459912 · doi ↗ · pubmed ↗
- 5Fanous J, Claudi B, Tripathi V, Li J, Goormaghtigh F, Bumann D. 2025. Limited impact of Salmonella stress and persisters on antibiotic clearance. Nature 639:181–189. doi:10.1038/s 41586-024-08506-639910302 PMC 11882453 · doi ↗ · pubmed ↗
- 6Mok WWK, Brynildsen MP. 2018. Timing of DNA damage responses impacts persistence to fluoroquinolones. Proc Natl Acad Sci USA 115:E 6301–E 6309. doi:10.1073/pnas.180421811529915065 PMC 6142227 · doi ↗ · pubmed ↗
- 7Völzing KG, Brynildsen MP. 2015. Stationary-phase persisters to ofloxacin sustain DNA damage and require repair systems only during recovery. m Bio 6:e 00731-15. doi:10.1128/m Bio.00731-1526330511 PMC 4556807 · doi ↗ · pubmed ↗
- 8Batchelder JI, Mahey N, Mok WWK. 2025. Post-fluoroquinolone treatment molecular events and nutrient availability modulate Staphylococcus aureus antibiotic persistence. bio Rxiv:2025.06.26.661800. doi:10.1101/2025.06.26.661800 · doi ↗