Deletion of tonB1 in Pseudomonas aeruginosa impairs zinc homeostasis and pathogenicity
Wenwen Li, Yu Zheng, Guifeng Wang, Juanli Cheng, Wei Xiao, Xin Ma, Panxin Li, Walter J. Chazin, Jinshui Lin

TL;DR
Deleting the TonB1 gene in Pseudomonas aeruginosa disrupts zinc balance and reduces its ability to cause disease.
Contribution
TonB1 is shown to be essential for zinc homeostasis and virulence in Pseudomonas aeruginosa.
Findings
Deleting tonB1 reduces zinc uptake and intracellular zinc levels in Pseudomonas aeruginosa.
The tonB1 mutant shows increased susceptibility to calprotectin and reduced virulence in infection models.
TonB1 is critical for bacterial fitness in Galleria mellonella hemolymph under zinc limitation.
Abstract
Many bacterial pathogens must acquire metal ions for proliferation and pathogenesis. In gram-negative bacteria, the TonB system is crucial for nutrient uptake. Previous research indicates that Pseudomonas aeruginosa uses the energy transduction protein TonB1 for iron uptake. Although zinc and iron are essential for P. aeruginosa, it is unknown whether TonB1 is also important for its zinc uptake. Here, a tonB1 deletion mutant was constructed from P. aeruginosa PAO1. Inductively coupled plasma mass spectrometry and other methods revealed that the tonB1 mutation significantly altered zinc homeostasis, as evidenced by diminished zinc uptake capacity, and affected other zinc-related phenotypes in P. aeruginosa, such as increased susceptibility to the host-secreted nutritional immunity protein calprotectin (CP), reduced oxidative stress resistance, impaired motility, and attenuated virulence…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
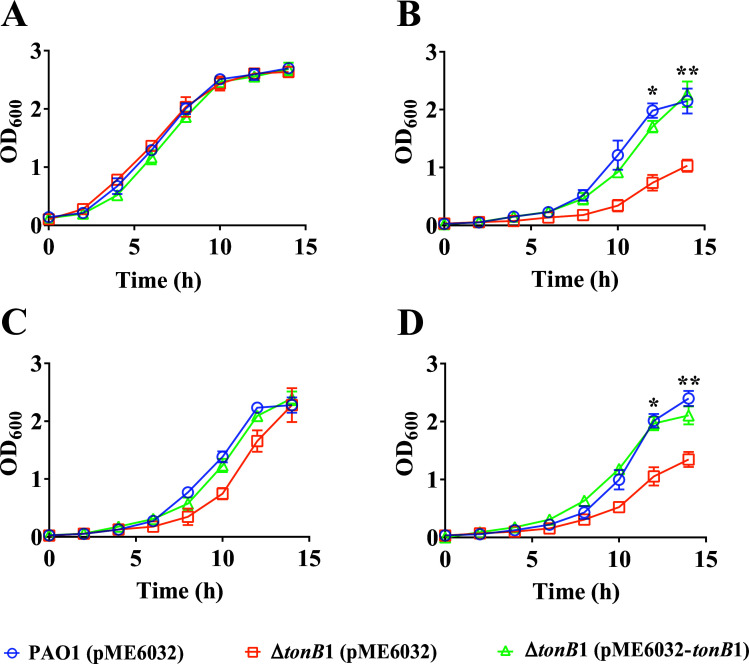 Fig 1
Fig 1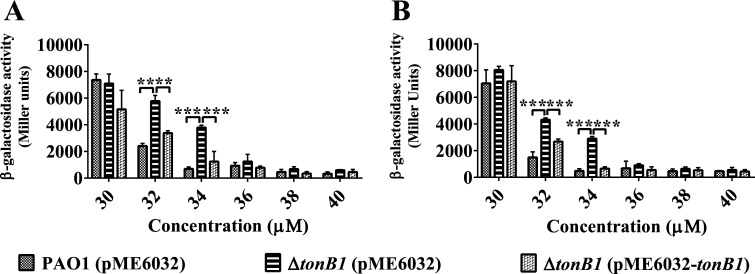 Fig 2
Fig 2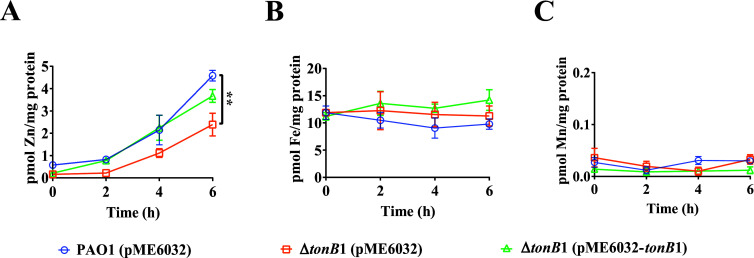 Fig 3
Fig 3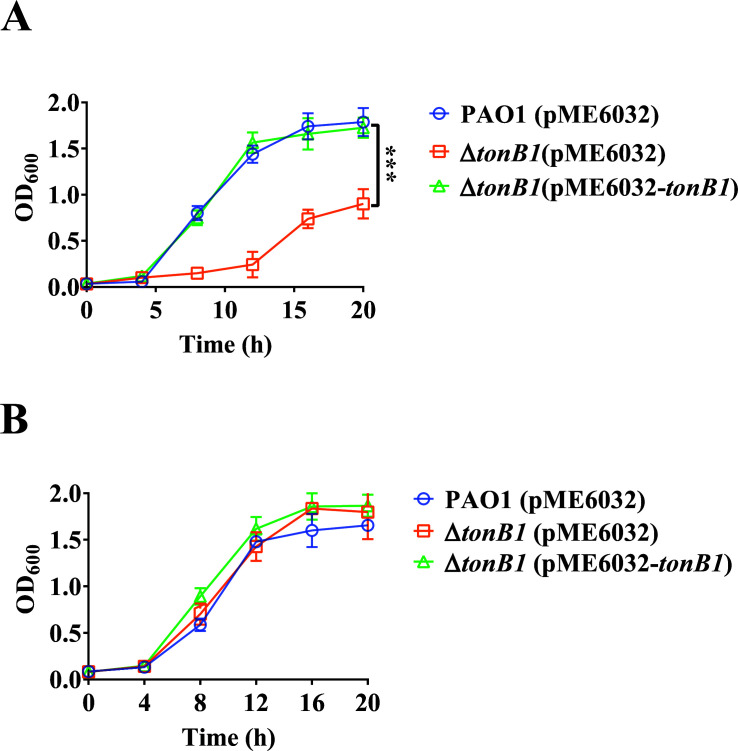 Fig 4
Fig 4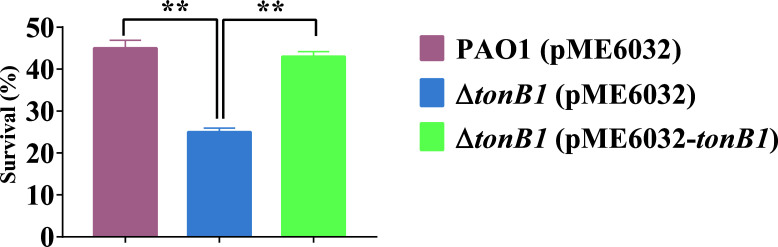 Fig 5
Fig 5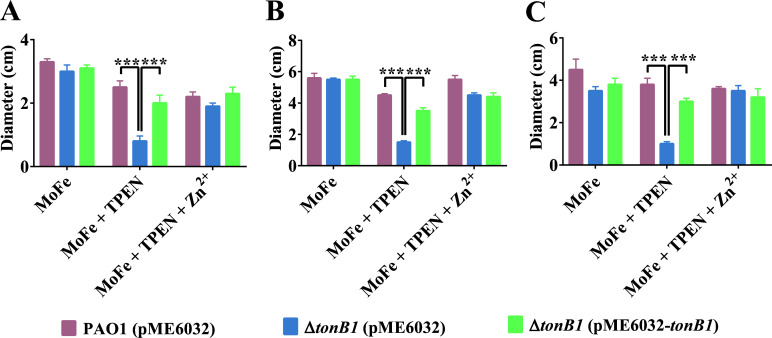 Fig 6
Fig 6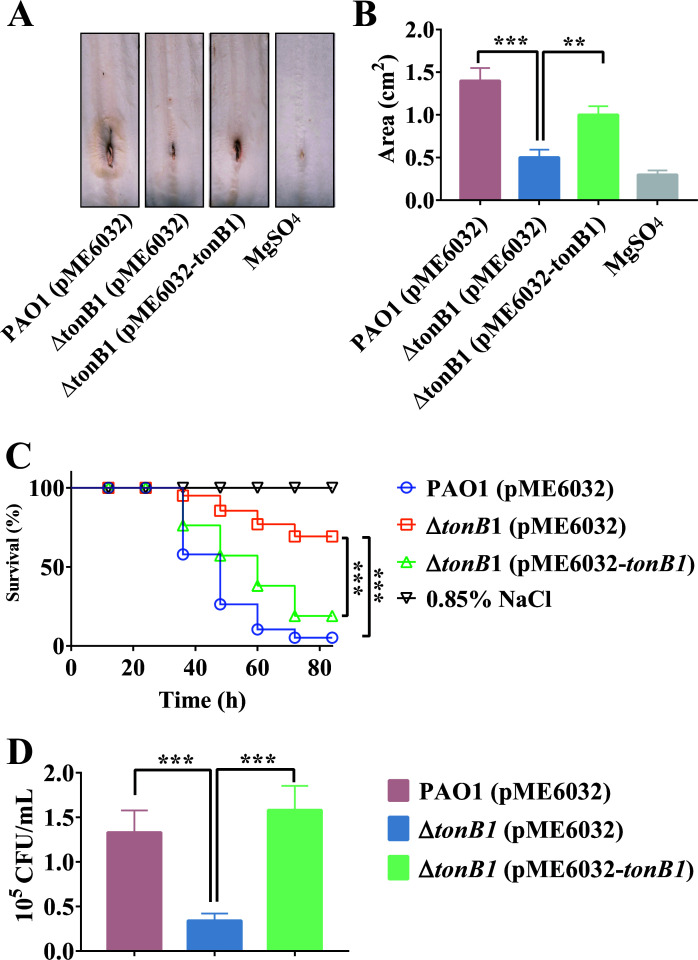 Fig 7
Fig 7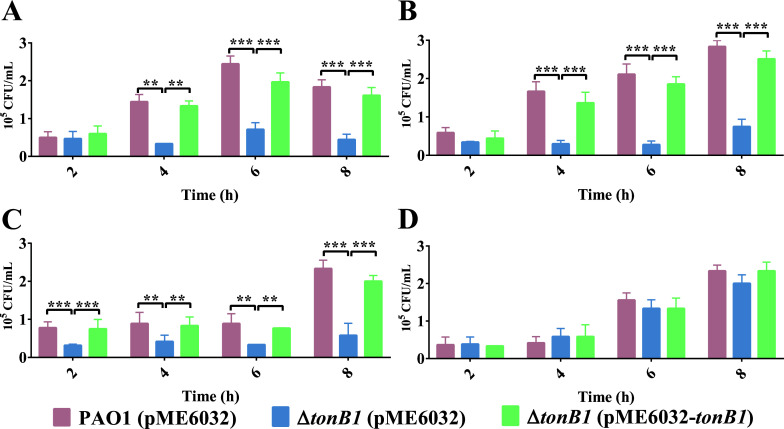 Fig 8
Fig 8- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Genetics and Biotechnology · Trace Elements in Health · Plant Micronutrient Interactions and Effects
INTRODUCTION
Zinc is an abundant metal element in organisms and is an essential trace micronutrient crucial for the growth of all life (1). Most zinc in cells is not free, but protein-bound. Bioinformatics analyses have suggested that approximately 4% of bacterial proteins contain non-heme iron, whereas 5%–6% contain zinc (2, 3). Zinc serves as a structural and catalytic cofactor for proteins, with numerous biological roles in bacteria (4). For example, zinc is incorporated into multiple transcription factors to mediate regulatory functions (5) and is also associated with fundamental metabolic enzymes, including RNA/DNA polymerases (6), alcohol dehydrogenases (7), and isomerases (8). Moreover, zinc is critical for the growth and infection of pathogenic bacteria in host organisms (9). Consequently, the sequestration of metal ions in vertebrate hosts serves as a significant defense mechanism to limit the growth of invading pathogens via a strategy known as “nutritional immunity” (10). Calprotectin (CP) plays a central role in nutritional immunity and is known to chelate various divalent ions, including iron [Fe(II)], zinc [Zn(II)], manganese [Mn(II)], and nickel [Ni(II)] (11). In particular, zinc plays a significant role in the pathogenicity and antibiotic resistance of pathogenic bacteria (12). Indeed, the development of new antimicrobial drugs includes strategies to interfere with or inhibit cellular zinc uptake (13).
Various clinical infections, such as bacteremia, urinary tract infections, respiratory infections, and burn infections, are frequently caused by Pseudomonas aeruginosa as the primary pathogenic strain (14, 15). P. aeruginosa is a gram-negative opportunistic pathogen (16) and is one of the most life-threatening bacteria owing to its significant antibiotic resistance capacities. Consequently, P. aeruginosa has been listed by the World Health Organization as a priority pathogen for research and development of new antibiotics (17, 18).
Transition metals, such as iron, zinc, and copper, are critical for microbial growth, necessitating a diverse array of transport systems (19). The active transport of these metal ions in gram-negative bacteria is facilitated by TonB systems comprised of cytoplasmic membrane proteins ExbB-ExbD and the periplasmic protein TonB (20). TonB spans the periplasm and provides energy to outer membrane receptors, thereby enabling the active transport of compounds across the membrane (21). Consequently, TonB has a vital role in nutrient acquisition (22). The P. aeruginosa genome encodes three tonB genes: tonB1 (PA5531) (23), tonB2 (PA0197) (24), and tonB3 (PA0406) (25) that encode proteins of 342, 270, and 319 amino acid (aa) residues, respectively. Previous studies have identified TonB1 as the primary TonB protein involved in P. aeruginosa iron transport (23, 24, 26). The tonB1 mutant shows impaired growth under iron-limiting conditions, along with defects in siderophore-mediated iron transport and heme utilization (27). Similar to iron, zinc is one of the abundant transition metals in organisms (28). In this study, we investigated the function of tonB1 in zinc acquisition by P. aeruginosa and evaluated its potential impact on the pathogenicity of this organism.
RESULTS
TonB1 is involved in P. aeruginosa zinc uptake
The strain background utilized in these studies was the P. aeruginosa reference strain PAO1 and its derived ΔtonB1 mutant. Previous studies have shown that the ΔtonB1 P. aeruginosa strain exhibits a growth defect in media without additional iron supplementation (26). Consequently, strain growth was investigated in a chemically defined medium (M9) containing only trace metal elements. As in previous reports (26), the ΔtonB1 mutant strain exhibited significant growth defects in M9 medium without additional iron supplementation (Fig. S1A). In M9 medium, as the concentration of added FeCl_3_ gradually increased, the growth defect phenotype of the ΔtonB1 mutant strain was gradually restored (Fig. S1). It was not until the concentration of added FeCl_3_ reached 400 μM that growth of the ΔtonB1 mutant strain fully returned to the level of the wild-type strain PAO1 (Fig. S1F). Here, we defined M9 medium supplemented with 400 μM FeCl_3_ as MFe medium, which was used consistently in subsequent experiments to ensure optimal growth conditions for the ΔtonB1 mutant. After supplementation with 200 μM of the metal chelator N,N,N′,N′-tetrakis (2-pyridylmethyl) ethylenediamine (TPEN), which has a particularly high affinity for zinc among several metals, the ΔtonB1 mutant strain (pME6032) exhibited a significant growth defect compared with the wild-type PAO1 (pME6032) and the complemented ΔtonB1-Com strain (pME6032-tonB1). The defect was then rescued by the addition of 100 μM ZnSO_4_, restoring growth to wild-type levels; the addition of 100 μM FeCl_3_ failed to restore growth (Fig. 1). Notably, both the wild-type PAO1 strain carrying the pME6032 plasmid and the complemented ΔtonB1-Com strain harboring pME6032-tonB1 displayed complete restoration of growth, confirming the specificity of the observed zinc-dependent growth defect in the ΔtonB1 mutant. The results demonstrated that TonB1 is involved in both Fe and Zn transport.
*Effect of tonB1 mutations on the growth of P. aeruginosa. (A) Growth curves of the P. aeruginosa wild-type strain, the ΔtonB1 mutant, and its complemented strain in MFe medium. (B) Supplemented with 200 μM TPEN. (C) Supplemented with 200 μM TPEN and 100 μM ZnSO4. (D) Supplemented with 200 μM TPEN and 100 μM FeCl3. All of the data represent the results of at least three independent experiments. Error bars indicate standard deviations. *: P < 0.05; *: P < 0.01.
P. aeruginosa possesses two primary zinc uptake systems: the ZnuABC uptake system (29) and the CntOLMI (also known as ZrmABCD) uptake system (30, 31). In addition to these characterized uptake systems, the PA4063-PA4066 operon, which encodes a MacB-family efflux transporter, has been shown to be transcriptionally regulated by zinc availability and the zinc-responsive regulator Zur (32). Although this operon was not originally associated with zinc uptake, its expression was monitored in this study as a potential indicator of intracellular Zn status. To investigate the potential involvement of TonB1 in zinc acquisition, we used Zn-responsive promoters from cntO and PA4063 to assess whether tonB1 inactivation affects the expression of genes associated with zinc homeostasis. Specifically, we constructed lacZ transcriptional fusion reporter strains, which contained either the PcntO promoter from the cntOLMI operon or the P_PA4063_ promoter from the PA4063–PA4066 operon. The impacts of tonB1 deletion on the expression of these two operons were assessed by measuring β-galactosidase activity in MFe medium supplemented with 50 μM TPEN, while varying Zn^2+^ concentrations. Notably, these culture conditions did not affect the growth of the P. aeruginosa strains tested (Fig. S2). When Zn^2+^ was supplemented at concentrations of 32–34 μM, the expression levels of both the cntOLMI and PA4063–PA4066 operons were significantly elevated in the ΔtonB1 (pME6032) mutant strain compared with the wild-type PAO1 (pME6032) and the complemented ΔtonB1 (pME6032–tonB1) strain (Fig. 2). Outside this narrow window, the phenotypic differences are masked at concentrations below 32 μM, because these operons are strongly activated even in the wild-type strain, and above 34 μM, because their expression becomes strongly repressed. The 32–34 μM range thus represents a transition zone for zinc-responsive expression. Despite this narrow observable range, these findings provide compelling evidence that TonB1 has a functional role in maintaining zinc homeostasis in P. aeruginosa.
*Effect of exogenous addition of Zn2+ on the expression of cntOLMI and PA4063-66. Cells were grown in MFe medium supplemented with 50 μM TPEN, while varying Zn2+ concentrations, and these culture conditions did not affect the growth of strains. Levels of cntOLMI and PA4063-66 transcription in P. aeruginosa mutant strains PAO1 (pME6032), ΔtonB1 (pME6032), and ΔtonB1 (pME6032-tonB1) were monitored using the cntO′–lacZ and PA4063′–lacZ transcriptional fusions, respectively. (A) Expression of the cntOLMI operon. (B) Expression of the PA4063–PA4066 operon. The graphs show the mean and standard deviation of two experiments performed in five replicates each time. **: P < 0.01; **: P < 0.001.
To further analyze the role of TonB1 in zinc uptake by P. aeruginosa, zinc-starved cells of PAO1 (pME6032), ΔtonB1 (pME6032), and ΔtonB1 (pME6032-tonB1)—where zinc starvation was achieved by pre-culturing in MFe medium supplemented with 50 μM TPEN—were cultured in MFe medium containing 34 μM ZnSO₄, followed by measurement of intracellular metal ion concentrations at various time points. This zinc-deficient cultivation had no impact on bacterial growth (Fig. S2). However, the intracellular zinc content of PAO1 (pME6032) cells gradually increased over time. In contrast, the ΔtonB1 (pME6032) mutant exhibited a significantly slower increase in intracellular zinc compared to the wild-type PAO1 (pME6032) cells (Fig. 3A). Additionally, significant differences in intracellular iron and manganese ion concentrations were not observed between the two strains (Fig. 3B and C). Furthermore, the complemented strain ΔtonB1 (pME6032-tonB1) exhibited increased intracellular zinc concentrations relative to those of the wild-type phenotype under the same culture conditions, while the intracellular iron and manganese ion concentrations remained unaffected. In identical experimental conditions, the tonB1 mutation did not lead to detectable effects on intracellular concentrations of nickel, cobalt, or copper ions (Fig. S3). Thus, mutation of tonB1 significantly altered the zinc homeostasis of P. aeruginosa under these experimental conditions.
*Effect of a tonB1 mutation on the intracellular metal ion content of P. aeruginosa. P. aeruginosa PAO1 (pME6032), ΔtonB1 (pME6032), and ΔtonB1 (pME6032-tonB1) were cultured in MFe medium supplemented with 50 μM TPEN to mid-log phase growth. Intracellular metal ion concentrations were determined using inductively coupled plasma mass spectrometry (ICP-MS) at different time points. (A) Variation in intracellular zinc concentrations. (B) Variation in intracellular iron concentrations. (C) Variation in intracellular manganese concentrations. All of the data represent the results of at least three independent experiments. Error bars indicate standard deviations. *: P < 0.01.
The tonB1 deletion enhances sensitivity of P. aeruginosa to calprotectin
The above results indicated that tonB1 is associated with zinc uptake by P. aeruginosa, suggesting that it may influence zinc-dependent phenotypes in this pathogen. CP, a heterodimeric EF-hand calcium-binding protein composed of S100A8 and S100A9 subunits, is a zinc-chelating protein secreted by neutrophils and macrophages. CP is highly abundant in certain innate immune cells and accounts for approximately 50% of cytoplasmic proteins in neutrophils (33). This protein contributes to host defense by mediating nutritional immunity, a mechanism that involves the sequestration of various divalent metal ions, including but not limited to zinc and manganese, at sites of infection (11). The ability of pathogens to counteract the antimicrobial effects of CP is largely dependent on their capacity to enhance metal uptake pathways or compete with CP for essential metals (34). To assess the impact of the tonB1 deletion on CP sensitivity, we evaluated bacterial growth in a defined medium supplemented with 200 μg mL^−1^ CP. A CP variant that is unable to bind transition metals (CP*) was used as a control (33). As shown in Fig. 4, the growth of ΔtonB1 (pME6032) was significantly inhibited by functional CP (Fig. 4A), whereas complementation with tonB1 restored the resistance of ΔtonB1 (pME6032-tonB1) to a level similar to that of PAO1 (pME6032). By contrast, no significant differences in growth were observed when CP was replaced with CP* (Fig. 4B). These findings suggest that tonB1 has a critical role in counteracting CP-mediated metal sequestration, potentially linked to its role in metal acquisition, and its deletion enhances the susceptibility of P. aeruginosa to CP.
Effect of CP on the growth of the P. aeruginosa tonB1 mutant. (A) Growth curves of P. aeruginosa wild-type strain, ΔtonB1, and its complemented strain in a defined medium supplemented with 200 μg mL−1 CP. (B) Medium was also supplemented with 200 μg mL−1 CP for the negative control. All of the data represent the results of at least three independent experiments. Error bars indicate standard deviations. **: P < 0.001.
The tonB1 deletion reduces oxidative stress resistance in P. aeruginosa
Oxidative stress in bacteria is induced by reactive oxygen species (ROS) that damage macromolecules, such as proteins and DNA, leading to cell apoptosis (35). The generation of ROS to combat pathogenic infections is also a crucial immune defense mechanism in hosts (36). To counteract the harmful effects of oxidative stress, bacteria have evolved multiple defense strategies. In addition to eliminating ROS via superoxide dismutase and catalase, bacteria also take up divalent metal ions (such as zinc) to compete with ferrous ions and inhibit the Fenton reaction, thereby reducing oxidative damage caused by ROS to cells (37). To investigate the impact of tonB1 on the oxidative stress resistance of P. aeruginosa, we evaluated the survival rates of PAO1 (pME6032), ΔtonB1 (pME6032), and ΔtonB1 (pME6032-tonB1) in MFe medium supplemented with 50 μM TPEN, as well as 1 mM H_2_O_2_. Under conditions that did not affect growth (Fig. S2), the survival rates of PAO1 (pME6032) and ΔtonB1 (pME6032-tonB1) were significantly higher than those of ΔtonB1 (pME6032) after H_2_O_2_ treatment, with no significant difference observed between PAO1 (pME6032) and ΔtonB1 (pME6032-tonB1) survival rates (Fig. 5). These results indicated that the deletion of tonB1 significantly reduces the resistance of P. aeruginosa to oxidative stress, likely associated with altered zinc homeostasis, in addition to the known role of TonB1 in iron transport.
*Effect of H2O2 on the growth of the P. aeruginosa tonB1 mutant. Survival rates of each strain following treatment with 1 mM H2O2 in MFe medium supplemented with 50 μM TPEN. All of the data represent the results of at least three independent experiments. Error bars indicate standard deviations. *: P < 0.01.
The deletion of tonB1 inhibits P. aeruginosa motility
P. aeruginosa exhibits three types of motility, including swarming, swimming, and twitching (38). Previous studies have demonstrated that zinc has a crucial role in regulating bacterial motility (39–42), including in P. aeruginosa (43). To investigate the effect of tonB1 on P. aeruginosa motility under zinc-limiting conditions, we compared the motility of various strains—including PAO1 (pME6032), ΔtonB1 (pME6032), and ΔtonB1 (pME6032-tonB1)—across different growth conditions. To overcome the iron-dependent growth defects exhibited by the ΔtonB1 mutant, a motility medium supplemented with 400 μM FeCl_3_ (designated MoFe medium) was used to ensure optimal growth of this strain in all assays. Notably, the addition of FeCl_3_ did not impair the motility phenotypes of any tested strain, as confirmed by the results shown in Fig. 6, thereby validating the use of MoFe medium as a consistent and reliable condition for subsequent experiments involving motility assessment under zinc limitation. When 500 μM TPEN was added to the MoFe medium to induce zinc limitation, the ΔtonB1 (pME6032) strain exhibited significantly impaired motility in comparison with both the wild-type PAO1 (pME6032) and the complemented strain ΔtonB1 (pME6032-tonB1). This defect was evident across all three major modes of motility: swarming, swimming, and twitching (Fig. 6). Importantly, supplementation with 400 μM ZnSO_4_ under these zinc-chelated conditions fully restored motility in the ΔtonB1 mutant to wild-type levels, confirming that the observed impairments were specifically attributable to zinc deficiency rather than nonspecific physiological stress (Fig. 6). Detailed representative images of swarming, swimming, and twitching motility are provided in Fig. S4 to S6, respectively. These findings suggest that tonB1 has a critical role in maintaining zinc homeostasis in P. aeruginosa, and its deletion leads to impaired motility under zinc-limited conditions, possibly due to the associated disruption in zinc availability.
*Effect of tonB1 deletion on P. aeruginosa (A) swarming, (B) swimming, and (C) twitching motility. The PAO1 (pME6032), ΔtonB1 (pME6032), and ΔtonB1 (pME6032-tonB1) strains were spotted onto MoFe medium containing 500 μM TPEN with or without 400 μM ZnSO4, and their motility diameters were measured. All of the data represent the results of at least three independent experiments. Error bars indicate standard deviations. **: P < 0.001.
The tonB1 deletion reduces P. aeruginosa virulence
To analyze the impact of tonB1 on P. aeruginosa virulence, the pathogenic capacities of different strains were evaluated using Chinese cabbage and Galleria mellonella larvae as infection models. Compared with PAO1 (pME6032), ΔtonB1 (pME6032) significantly reduced P. aeruginosa virulence against both Chinese cabbage and G. mellonella larvae, while complementation with tonB1 restored the virulence phenotype to levels comparable to PAO1 (pME6032) (Fig. 7A through C). Additionally, colony counts from G. mellonella infected with PAO1 (pME6032), ΔtonB1 (pME6032), and ΔtonB1 (pME6032-tonB1) revealed that the bacterial load of ΔtonB1 (pME6032) was significantly lower than for PAO1 (pME6032) and ΔtonB1 (pME6032-tonB1) (Fig. 7D). These data indicate that the tonB1 deletion mutation attenuates the virulence of P. aeruginosa.
*Deletion of tonB1 reduces P. aeruginosa virulence. (A and B) Lesion areas of Chinese cabbage treated with different P. aeruginosa strains. (C) Survival curves of G. mellonella larvae treated with different P. aeruginosa strains. (D) Bacterial colony counts from G. mellonella larvae treated with different P. aeruginosa strains. All of the data represent at least three independent experiments. Error bars indicate standard deviations. **: P < 0.001.
The tonB1 deletion affects the adaptability of P. aeruginosa in G. mellonella hemolymph
When the strains were added to G. mellonella hemolymph without supplemental metal ions, the ΔtonB1 (pME6032) bacterial abundances did not significantly change over time, while the abundances of PAO1 (pME6032) and ΔtonB1 (pME6032-tonB1) significantly increased after 4 h (Fig. 8). The bacterial abundances of ΔtonB1 (pME6032) were significantly lower than those of PAO1 (pME6032) and ΔtonB1 (pME6032-tonB1) after 4 h (Fig. 8A). When 1 μM FeCl_3_ or 1 μM ZnSO_4_ was added to hemolymph, the bacterial abundances of ΔtonB1 (pME6032) did not increase and remained significantly lower than those of PAO1 (pME6032) and ΔtonB1 (pME6032-tonB1) (Fig. 8B and C). However, when both 1 μM FeCl_3_ and 1 μM ZnSO_4_ were added to hemolymph, the bacterial abundances of ΔtonB1 (pME6032) significantly increased after 4 h, reaching levels comparable to those of PAO1 (pME6032) and ΔtonB1 (pME6032-tonB1), with no significant differences observed among them (Fig. 8D). Thus, tonB1 deletion compromises zinc and iron acquisition in P. aeruginosa, thereby diminishing its fitness in G. mellonella hemolymph.
*Deletion of tonB1 reduces the adaptability of P. aeruginosa in G. mellonella hemolymph. Variation in bacterial load within the hemolymph of G. mellonella infected with different strains. Experiments were conducted (A) without metal ion supplementation, (B) supplemented with 1 μM FeCl3, (C) supplemented with 1 μM ZnSO4, or (D) supplemented with 1 μM FeCl3 and 1 μM ZnSO4. All of the data represent at least three independent experiments. Error bars indicate standard deviations. **: P < 0.01; **: P < 0.001.
DISCUSSION
In this study, we demonstrated that TonB1, previously characterized primarily for its role in iron acquisition in P. aeruginosa, also plays a critical role in maintaining zinc homeostasis. The tonB1 deletion mutant exhibited significant growth defects under zinc-limiting conditions, alongside reduced intracellular zinc levels and dysregulation of key zinc uptake-related genes. These physiological impairments were associated with a range of downstream phenotypic consequences, including heightened susceptibility to CP-mediated nutritional immunity, reduced oxidative stress resistance, impaired motility, and attenuated virulence in both plant and insect infection models. This work not only expands the known functions of TonB1 in P. aeruginosa, but also supports the growing recognition that TonB proteins are not solely involved in iron transport in gram-negative bacteria.
The involvement of TonB in zinc acquisition is not entirely without precedent. Recent studies have begun to uncover the role of the TonB system in the uptake of other transition metals beyond iron. For example, in Neisseria meningitidis, the TonB-dependent receptor ZnuD has been shown to mediate zinc acquisition, and its expression is upregulated under zinc-limiting conditions (44). A similar mechanism has been observed in Caulobacter crescentus, where the TonB-dependent transporters ZnuK and ZnuL are highly expressed during zinc starvation and are essential for bacterial survival under such conditions, serving as key components of zinc scavenging systems (45). Furthermore, a more complex and sophisticated zinc acquisition system has been identified in Burkholderia thailandensis, involving a Zur-regulated type VI secretion system (T6SS) and a TonB-dependent outer membrane heme transporter, HmuR (46). In this system, zinc uptake is facilitated by T6SS-mediated secretion of a zinc-binding effector protein, TseZ, which collaborates with HmuR to drive the active transport of zinc across the outer membrane (46). Our findings in P. aeruginosa are consistent with and extend the above observations, providing direct experimental evidence that TonB1 has a non-redundant role in zinc homeostasis and pathogenicity.
Our experimental conditions were designed to specifically assess the role of TonB1 in zinc uptake by supplementing the culture medium with high concentrations of iron (400 μM FeCl_3_), a necessary measure to overcome the iron-dependent growth defects of the ΔtonB1 mutant, as previously reported (26, 27) (also see Fig. S1). Under these iron-replete conditions, loss of TonB1 still led to altered zinc homeostasis, as evidenced by reduced intracellular zinc levels measured using inductively coupled plasma mass spectrometry (ICP-MS; Fig. 3A), and increased expression of zinc-responsive promoters, such as PcntO and P_PA4063_, in the mutant background (Fig. 2), suggesting compensatory activation of alternative zinc acquisition systems. Under these optimized conditions, the ΔtonB1 mutant also exhibited sensitivity to the zinc chelator TPEN, which could only be rescued by exogenous zinc, not iron, demonstrating that the observed zinc-related phenotypes are linked to the requirement for TonB1 under these specific conditions (Fig. 1A through D). The intracellular iron concentration in P. aeruginosa PAO1 cells reaches 4.230 mM when cultured in LB medium, whereas the iron content in the LB medium itself is 4.3 μM (47, 48). Although the externally added iron concentration in our experiments was supraphysiological for P. aeruginosa, the use of 400 μM FeCl_3_ was essential to uncouple the zinc-related phenotypes from the iron-dependent growth defect of the ΔtonB1 mutant. Collectively, these findings strongly support an important and specific role for TonB1 in zinc homeostasis under our experimental conditions, independent of its well-established function in iron transport. We acknowledge that the supraphysiological iron concentration used here could represent a caveat, as metal ratio disequilibrium is known to inhibit the import systems of non-cognate metals (49). While we cannot fully rule out that high iron may have partially exacerbated the phenotype by competing for shared import machinery, the stark contrast in growth between the mutant and wild type under zinc-limiting conditions specifically argues that the primary defect is linked to disrupted zinc homeostasis resulting from the loss of TonB1 function.
Our experimental approach utilized a high concentration of FeCl₃ (400 µM) specifically to overcome the well-characterized iron-auxotrophy of the ΔtonB1 mutant (26, 27). This design was a deliberate strategy to circumvent the primary iron-acquisition defect, thereby enabling us to dissect and focus on the potential role of TonB1 in zinc homeostasis. Consequently, under these iron-replete conditions, the lack of a significant defect in intracellular iron accumulation in the ΔtonB1 mutant (Fig. 3B) is likely attributable to the accessibility of excess environmental iron via TonB1-independent pathways. For example, Fe³^+^ can be reduced to Fe²^+^ by phenazines produced by P. aeruginosa, with subsequent uptake via the FeoABC system (50, 51). This compensatory mechanism effectively masks the expected iron-uptake defect in our assay, allowing us to isolate and study the zinc-related phenotypes associated with the tonB1 deletion.
The zinc uptake systems in P. aeruginosa mainly include the ABC transporter ZnuABC and the CntOLMI (also known as ZrmABCD) system (30, 31). Although both systems are functional, our data indicate that the absence of TonB1 leads to a significant reduction in zinc uptake, even in the presence of these dedicated zinc transporters. This suggests that TonB1 may either directly facilitate zinc uptake or indirectly regulate the expression or function of these systems, or that its deletion triggers a broader physiological disruption affecting metal homeostasis. The elevated expression of cntO and PA4063 in the ΔtonB1 mutant under zinc-limiting conditions further supports the hypothesis that TonB1 has a central role in maintaining zinc homeostasis.
One of the most striking phenotypic consequences of impaired zinc uptake in the tonB1 mutant was its heightened sensitivity to CP, a key component of the host’s nutritional immunity strategy. In the presence of functional CP, growth of the tonB1 mutant was significantly inhibited, whereas the complemented strain and wild-type strain were resistant (Fig. 4A). This result underscores that TonB1 is necessary to resist CP-imposed metal starvation, a critical aspect of bacterial pathogenesis. Regarding the use of CP, we acknowledge that its metal-binding buffer contains β-mercaptoethanol, which can reduce Fe^3+^ to Fe^2+^, a known ligand for CP (52, 53). However, in our experiments, the ΔtonB1 mutant exhibited sensitivity to CP despite being grown in medium supplemented with 400 μM FeCl_3_. Given that TonB1 is essential for iron uptake via siderophores and heme (23, 26), the high iron concentration theoretically compensates for the loss of TonB1. Therefore, the observed sensitivity to CP is potentially due to the mutant’s inability to acquire zinc, rather than iron, under these conditions. We cannot, however, exclude the possibility that other TonB1-dependent processes contribute to this phenotype.
The mutant also exhibited reduced survival under oxidative stress conditions (Fig. 5), which may be attributed to the role of zinc in stabilizing cellular components and in protecting against ROS via mechanisms such as competitive inhibition of the Fenton reaction (37). Another key finding of this study is the role of TonB1 in bacterial motility. Under zinc-limiting conditions, the tonB1 mutant showed significant impairments in all three forms of motility—swarming, swimming, and twitching—relative to the wild-type and complemented strains (Fig. 6). The motility defects were rescued upon zinc supplementation, indicating that they were specifically owing to zinc deficiency rather than a general metabolic perturbation. This result aligns with previous reports showing that zinc is essential for the proper functioning of flagellar and type IV pilus systems, which are critical for bacterial motility and host colonization (43). The motility defects observed upon tonB1 deletion may therefore stem from disrupted zinc homeostasis or from other TonB1-mediated functions.
Finally, the deletion of tonB1 in P. aeruginosa significantly attenuated its virulence (Fig. 7), as demonstrated in both the Chinese cabbage and G. mellonella infection models. This decreased pathogenicity is correlated with impaired bacterial fitness in G. mellonella hemolymph, particularly under metal-limited conditions (Fig. 8). Whereas the addition of either iron or zinc alone failed to restore the growth capacity of the ΔtonB1 mutant, supplementation of both metals together rescued its fitness to levels comparable with those of the wild-type and complemented strains in G. mellonella hemolymph. These findings suggest that TonB1 has a critical role in the acquisition of both iron and zinc, or in coordinating their homeostasis, and that both metals contribute to the overall virulence of P. aeruginosa. While our data strongly associate the tonB1 deletion with zinc-related defects, the precise mechanistic link between TonB1, zinc uptake, and each specific phenotype requires further investigation. Alternative explanations, such as pleiotropic effects of the mutation on cellular physiology, cannot be fully ruled out.
In summary, our study provides compelling evidence that TonB1 in P. aeruginosa is not only essential for iron uptake but also has a central role in maintaining zinc homeostasis under the conditions tested. This association underscores the pleiotropic functions of TonB proteins in bacterial physiology and pathogenicity. Given the increasing recognition of metal homeostasis as a viable target for antimicrobial therapy, our findings suggest that TonB1 may be a promising target for the development of novel therapeutics aimed at disrupting bacterial nutrient acquisition and virulence.
MATERIALS AND METHODS
Bacterial strains and growth conditions
Bacterial strains and plasmids used in this study are shown in Table S1. Escherichia coli strains were grown at 37°C in either LB or TSB medium. P. aeruginosa strains were grown at 37°C in either LB, TSB, M9 (54, 55), or MFe medium. MFe medium was specifically defined as M9 medium supplemented with 400 μM FeCl_3_, a formulation introduced to support optimal growth of the ΔtonB1 mutant. P. aeruginosa PAO1 was the parent strain of all derivatives used in this study. To generate the tonB1 mutant, pK18mobsacB derivatives were transformed into relevant P. aeruginosa strains through E. coli S17-1-mediated conjugation and screened as previously described (56). During screening, FeSO_4_ (40 μM) was added to the agar to assist growth (26). To achieve overexpression or complementation in various P. aeruginosa strains, pME6032 derivatives were transformed into relevant P. aeruginosa strains and induced by the addition of 1 mM isopropyl-β-D-1-thiogalactopyranoside (IPTG). The following antibiotics were used at the indicated concentrations for P. aeruginosa: kanamycin (30 µg/mL), chloramphenicol (30 µg/mL), gentamicin (200 µg/mL), and tetracycline (160 µg/mL for liquid growth or 200 µg/mL for solid growth). The following antibiotics were used at the indicated concentrations for E. coli: kanamycin (30 µg/mL), gentamicin (10 µg/mL), and tetracycline (20 µg/mL).
Plasmid construction
The construction of the knockout plasmid was conducted, as previously described, with some modifications (56). Briefly, construction of the tonB1 gene recombinant suicide plasmids for deletion was achieved by amplification of 954 bp upstream and 883 bp downstream fragments flanking the tonB1 gene using the primer pairs tonB1 Up F/tonB1 Up R and tonB1 Low F/tonB1 Low R, respectively (Table S2). The upstream and downstream polymerase chain reaction (PCR) fragments were then ligated using overlapping PCR, and the resulting PCR products were inserted into the Bam HI/Hind III sites of the suicide vector pK18mobsacB to generate the plasmid p-tonB1. The gentamicin resistance cassette from p34s-Gm was subsequently inserted into the same Hind III site of p-tonB1 to yield the recombinant suicide plasmid pK18-ΔtonB1.
To construct the complementation plasmid pME6032-tonB1, PCR-amplified tonB1 was inserted into the EcoR I/Bgl II sites of pME6032, generating the recombinant plasmid pME6032-tonB1. The recombinant plasmids pBBR1MCS-5-tonB1 were constructed using the same method.
cntO′–lacZ transcriptional fusions were constructed via PCR amplification of the 737 bp upstream DNA region of the cntO gene using the primer pairs cntO F/cntO R (Table S2). The PCR amplification products were directly cloned into the pMini-CTX*:: lacZ* vector (56), yielding lacZ reporter constructs. The recombinant plasmids PA4063′-lacZ were then constructed using the same method (Table S1).
Growth assays
Growth assays were conducted as previously described, but with some modifications (54). Briefly, P. aeruginosa strains were grown overnight in LB medium supplemented with 400 μM FeCl_3_. Overnight cultures were harvested, and the cells were washed twice with M9 medium to remove residual nutrients before subculturing. The washed cells were then transferred to an appropriate fresh medium and adjusted to an initial OD_600_ (optical density at 600 nm) of 0.01. The cultures were incubated at 37°C, and OD_600_ measurements were taken every 2 h over a 24-h period.
β-galactosidase assays
β-galactosidase assays were conducted, as previously described, but with modifications (57). Briefly, 200 µL of the bacterial culture was transferred to measure OD_600_. Then, 50 µL of culture was mixed with 420 µL of Z buffer, 20 µL of chloroform, and 10 µL of 0.1% sodium dodecyl sulfate. The solutions were thoroughly mixed and incubated at 30°C for 1 h. After incubation, reactions were initiated by adding 100 µL of 4 mg/mL o-nitrophenyl-β-D-galactopyranoside (Sigma, St. Louis, MO, USA). Once color developed in the reaction mixture, the reaction was quenched by adding 250 µL of 1 mol/L Na_2_CO_3_, and the reaction time was recorded. The mixture was then centrifuged at 12,000 rpm for 3 min, followed by OD_420_ and OD_550_ measurements of the supernatant. β-galactosidase activity was then calculated in Miller units (MUs) according to the following equation:
Measurement of intracellular metal ion concentrations
Intracellular metal ion concentrations were evaluated based on previous descriptions (55). P. aeruginosa strains were grown in 5 mL LB medium supplemented with 400 μM FeCl_3_ at 37°C with shaking at 200 rpm for 20 h. Cells from 1 mL of culture medium were harvested and washed twice with M9 medium, followed by sub-culturing (1:100) in MFe medium that was supplemented with 1 mM IPTG and 50 μM TPEN until exponential phase growth was achieved. After centrifugation at 2,000 × g for 10 min at 4°C, pellets were resuspended in PBS containing 1 mM EDTA, washed twice under the same conditions, and suspended in PBS. The bacterial suspensions were divided into four aliquots, each supplemented with 0.4% glucose, 400 μM FeCl_3_, and 34 μM ZnSO_4_. The aliquots were then incubated at 37°C (200 rpm) for 0, 2, 4, or 6 h, followed by centrifugation at 2,000 × g for 20 min at 4°C. Pellets were then washed twice with PBS-EDTA and once with PBS, then weighed to determine wet cell mass. Bacterial lysis was performed using the BugBuster (Novagen) reagent at 5 mL per gram wet pellet, followed by 20 min of gentle rotation. Total protein content was then quantified via the Bradford assay (Bio-Rad). Samples were diluted 100-fold in 3% nitric acid to a final volume of 10 mL and analyzed by ICP-MS (Varian 802-MS instrument). Buffer-matched standards and dilution factors were used for calibration. All experiments were conducted in triplicate with at least three biological replicates.
Calprotectin resistance assays
CP resistance assays were conducted as previously described, but with some modifications (58). P. aeruginosa strains were cultured overnight at 37°C and in 5 mL of LB liquid medium supplemented with 400 μM FeCl_3_ with shaking at 200 rpm. To assess CP resistance, bacteria were inoculated into a growth medium composed of 60% M9 minimal medium and 40% CP buffer (20 mM Tris-HCl, pH 7.5, 100 mM NaCl, 5 mM β-mercaptoethanol, 3 mM CaCl_2_, and 500 μg mL^−1^ CP or CP* with 400 μM FeCl_3_. CP and the CP* variant, unable to bind transition metals, were produced as described previously (33). Overnight cultures of PAO1 wild-type or the ΔtonB1 mutant were diluted 1:100 into the assay medium and incubated at 37°C with aeration. Bacterial growth was monitored by measuring optical density at regular 2-h intervals throughout the time course. All experiments were carried out with a minimum of three biological replicates and independently repeated three times to ensure reproducibility.
Oxidative stress assays
P. aeruginosa strains were cultured in 5 mL of LB liquid medium with 400 μM FeCl_3_ at 37°C for 8 h and with shaking at 200 rpm. Cells were subcultured in 5 mL of fresh LB medium with 400 μM FeCl_3_ using a 1% inoculum until the logarithmic growth phase. The cultures were then subcultured into MFe medium supplemented with 50 μM TPEN using 1% inocula. A control group without any treatment and an experimental group treated with 1 mM H_2_O_2_ were used. Both groups were incubated at 37°C for 1 h with shaking at 220 rpm. Each treatment was performed in triplicate. After treatment, bacterial suspensions were serially diluted 10-fold, and 3 µL of each dilution was spotted onto LB plates containing 400 μM FeCl_3_. The plates were then incubated overnight at 37°C, followed by colony counting to calculate the survival rate as follows:
Motility assays
Swarming, swimming, and twitching motility assays were performed as previously described (43, 59). Swarming motility plates comprised 0.8% nutrient broth, 0.5% glucose, and 0.5% agar, while swimming motility plates comprised LB broth with 0.3% agar, and twitching motility plates comprised 1% peptone, 0.5% yeast extract, 0.5% NaCl, and 1% agar. To address the iron-dependent growth deficiency of the ΔtonB1 mutant, all motility assays were performed using media supplemented with 400 μM FeCl_3_, which we defined as MoFe medium. This modification ensured optimal growth conditions for the ΔtonB1 mutant and was consistently applied in all subsequent experiments. For swarming and swimming assays, test strains were grown in 5 mL of liquid LBΒ containing 400 μM FeCl_3_ at 37°C with shaking at 220 rpm until the culture reached an OD_600_ of 0.80. Subsequently, 3 µL of each bacterial suspension was inoculated into the center of the corresponding agar plate; plates were incubated upright at 30°C for 24 h to assess the radial expansion of bacterial migration. For twitching motility assays, single colonies were transferred using a sterile toothpick and stab-inoculated into the bottom surface of the agar in twitching plates. Plates were then incubated inverted at 37°C for 24 h. After incubation, plates were stained with a staining solution (0.05% Coomassie Brilliant Blue R250, 40% methanol, and 10% acetic acid) for 3 h. The plates were then rinsed with industrial alcohol until the diffusion zones of bacterial movement at the bottom of the plates became visible.
Chinese cabbage infection assays
Chinese cabbage infection assays were performed as previously described (60). Test strains were inoculated into 5 mL of liquid LB medium with 400 μM FeCl_3_ and cultured at 37°C with shaking at 220 rpm until reaching the stationary phase. A 1 mL aliquot of bacterial culture was centrifuged at 5,500 rpm for 5 min to collect bacterial cells. The cells were then washed twice and resuspended in 10 mmol/L MgSO_4_, followed by adjustment of the OD_600_ to 2.00 for subsequent use. Fresh Chinese cabbage leaves with consistent growth conditions were selected for inoculation and surface-sterilized with 0.1% H_2_O_2_. A microsyringe was used to inoculate 10 μL of the pretreated bacterial suspension onto the abaxial sides of the leaves. The inoculated leaves were then incubated at 30°C under humid conditions for 6 days. Infections at the inoculation sites were observed and analyzed using the ImageJ software program.
G. mellonella killing assays
The G. mellonella infection assays were conducted as previously described, with some modifications (61). Briefly, P. aeruginosa strains were cultured in 5 mL of LB liquid medium with 400 μM FeCl_3_ at 37°C overnight with shaking at 200 rpm. Subculturing was conducted in 5 mL of fresh LB with 400 μM FeCl_3_ at a 1:100 ratio until the OD_600_ value reached 0.5. Bacterial cells were then collected via centrifugation at 4°C and 5,500 rpm for 5 min. Cells were suspended in 0.85% NaCl solution, centrifuged at 4°C and 5,500 rpm for 5 min, and then collected, with the entire procedure repeated twice. Cells were then suspended and diluted with 0.85% NaCl solution to an abundance of 2 × 10^7^ CFU/mL. G. mellonella larvae were placed on ice for 5 min initiate anesthesia. A microsyringe was used to inject 10^5^ cells into the hemocoel of 3-day-old, fifth-instar G. mellonella larvae, with 0.85% NaCl solution used as the control. Fifty G. mellonella larvae were injected for each group and cultured at 25°C in the dark. The procedure was repeated in triplicate for each strain. Survival data were recorded every 12 h and then analyzed using Kaplan–Meier survival curves. Statistical significance in survival difference was assessed using a Mantel–Cox log-rank test and then applying Bonferroni’s correction for multiple comparisons.
Bacterial cellular abundance in the hemolymph of G. mellonella after infection was enumerated as previously described, but with some modifications (55). Following the above procedure, a microsyringe was used to inject 10^5^ cells into the hemocoel of each G. mellonella larva (fifth instar, day 3). The larvae were incubated in the dark at 25°C for 36 h. After infection, larvae were placed on ice for 5 min to anesthetize them. Pre-chilled 1.5 mL centrifuge tubes were also placed on ice, and 2 µL of 1% phenylthiourea (PTU) solution was added to each tube. Hemolymph from five larvae per group was collected into the tubes, with three replicates used per group. The collected hemolymph was serially diluted 10-fold using PBS buffer, followed by bacterial cultivation and colony counting.
Bacterial growth in G. mellonella hemolymph
The assay to measure bacterial growth in G. mellonella hemolymph was adapted from a previous study (62) with modifications. Briefly, hemolymph was collected from G. mellonella larvae (fifth instar, day 3). Larvae were placed on ice for 5 min to anesthetize them, and pre-chilled 1.5 mL centrifuge tubes were placed on ice, followed by the addition of 10 µL of 1% PTU solution to each tube. A total of 500 µL of hemolymph was collected from the larvae into the tubes, with three replicates used per group. The hemolymph was centrifuged at 20,630 × g for 15 min at 25°C, and the supernatant was then filter-sterilized (0.45 μm). Each bacterial strain was inoculated into liquid LB medium with 400 μM FeCl_3_ and cultured until reaching the stationary phase, followed by transfer to fresh liquid LB medium supplemented with 400 μM FeCl_3_. A 1 mL aliquot of each bacterial culture grown to an OD_600_ of 1.00 was centrifuged at 5,500 rpm for 5 min to collect the bacterial cells. Cells were washed twice with PBS and resuspended in PBS to a concentration of 1 × 10^5^ CFU/mL for subsequent use. A 5 µL aliquot of the bacterial suspension was inoculated into 500 µL of the hemolymph supplemented with appropriate antibiotics, TPEN, ZnSO_4_, or FeCl_3_ at specified concentrations. The cultures were then incubated at 25°C and 70 rpm, with colony counting performed every 2 h.
Statistical analyses
All of the experiments were performed in triplicate and repeated on two different occasions. Data are expressed as means ± S.D. Differences between frequencies were assessed using Student’s t-tests (two-tailed and unpaired), and a P value <0.05 was considered statistically significant. Shapiro–Wilk and one-way analysis of variance tests were performed using the GraphPad Prism version 7.00 software program (GraphPad Software Inc., San Diego, CA, USA) to examine the normality of the data and the homogeneity of variances, respectively. GraphPad Prism 7 and Adobe Illustrator 2023 (CS6; Adobe, Mountain View, CA, USA) were used to generate figures.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Coverdale JPC, Khazaipoul S, Arya S, Stewart AJ, Blindauer CA. 2019. Crosstalk between zinc and free fatty acids in plasma. Biochim Biophys Acta Mol Cell Biol Lipids 1864:532–542. doi:10.1016/j.bbalip.2018.09.00730266430 PMC 6372834 · doi ↗ · pubmed ↗
- 2Andreini C, Banci L, Bertini I, Rosato A. 2006. Zinc through the three domains of life. J Proteome Res 5:3173–3178. doi:10.1021/pr 060369917081069 · doi ↗ · pubmed ↗
- 3Andreini C, Bertini I, Rosato A. 2009. Metalloproteomes: a bioinformatic approach. Acc Chem Res 42:1471–1479. doi:10.1021/ar 900015 x 19697929 · doi ↗ · pubmed ↗
- 4Andreini C, Bertini I, Cavallaro G, Holliday GL, Thornton JM. 2008. Metal ions in biological catalysis: from enzyme databases to general principles. J Biol Inorg Chem 13:1205–1218. doi:10.1007/s 00775-008-0404-518604568 · doi ↗ · pubmed ↗
- 5Rasmussen RA, Wang S, Camarillo JM, Sosnowski V, Cho B-K, Goo YA, Lucks JB, O’Halloran TV. 2022. Zur and zinc increase expression of E. coli ribosomal protein L 31 through RNA-mediated repression of the repressor L 31p. Nucleic Acids Res 50:12739–12753. doi:10.1093/nar/gkac 108636533433 PMC 9825181 · doi ↗ · pubmed ↗
- 6Scrutton MC, Wu CW, Goldthwait DA. 1971. The presence and possible role of zinc in RNA polymerase obtained from Escherichia coli. Proc Natl Acad Sci USA 68:2497–2501. doi:10.1073/pnas.68.10.24974944629 PMC 389452 · doi ↗ · pubmed ↗
- 7Potter AJ, Kidd SP, Mc Ewan AG, Paton JC. 2010. The Mer R/Nml R family transcription factor of Streptococcus pneumoniae responds to carbonyl stress and modulates hydrogen peroxide production. J Bacteriol 192:4063–4066. doi:10.1128/JB.00383-1020525825 PMC 2916378 · doi ↗ · pubmed ↗
- 8Carrigan CN, Poulter CD. 2003. Zinc is an essential cofactor for type I isopentenyl diphosphate:dimethylallyl diphosphate isomerase. J Am Chem Soc 125:9008–9009. doi:10.1021/ja 035038115369345 · doi ↗ · pubmed ↗