Genomic and phenotypic analysis of ST25 A. baumannii identifies virulence-associated clades and capsular/outer core locus types
Antonella Migliaccio, Thibault Destanque, Marisa Haenni, Jean-Yves Madec, Keith A. Jolley, Maria Stabile, Eliana De Gregorio, Agnese Lupo, Raffaele Zarrilli

TL;DR
This study identifies specific virulence traits in ST25 Acinetobacter baumannii strains, showing that certain clades are more persistent and pathogenic.
Contribution
The study reveals novel virulence-associated clades and capsular/outer core locus types in ST25 A. baumannii.
Findings
CIVb and CIVd ST25 isolates showed higher virulence and resistance to environmental stressors.
KL14 and OCL6 genotypes correlate with increased resistance to oxidative stress and desiccation.
Phenotypic differences among isolates are linked to clade identity rather than origin.
Abstract
The increase in the infection caused by Acinetobacter baumannii is sustained by the selection of distinct epidemic clonal lineages, which are frequently resistant to a broad range of antimicrobials and possess virulence traits responsible for their persistence in the contaminated environment and spread among patients. The present study aimed to perform an integrated genomic and phenotypic analysis to assess the virulence features of ST25 isolates. A. baumannii isolates assigned to the ST25 epidemic clonal lineage shared high genomic similarity and clustered in four clades (I, II, III, and IV), with clade IV further subdivided into CIVa, CIVb, CIVc, and CIVd. Capsular locus (KL) KL14 was the predominant KL type (47%). Accessory genome analysis showed the presence of tartrate metabolism genes only in CII genomes. CIVb and CIVd ST25 A. baumannii isolates showed higher ability to infect…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
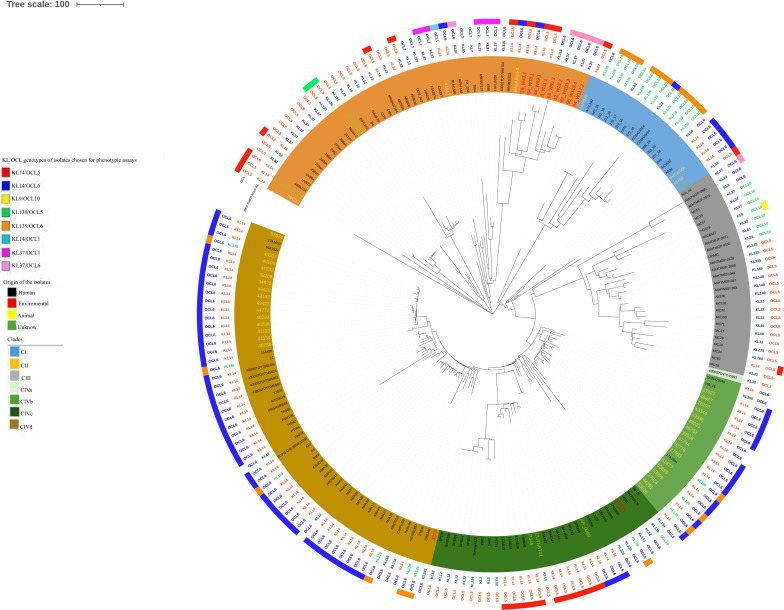 Fig 1
Fig 1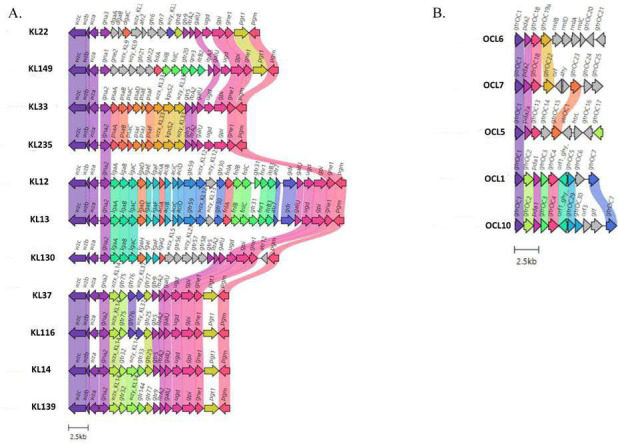 Fig 2
Fig 2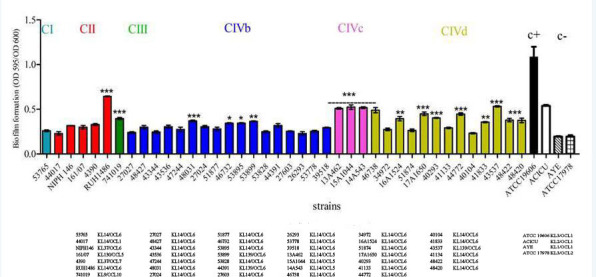 Fig 3
Fig 3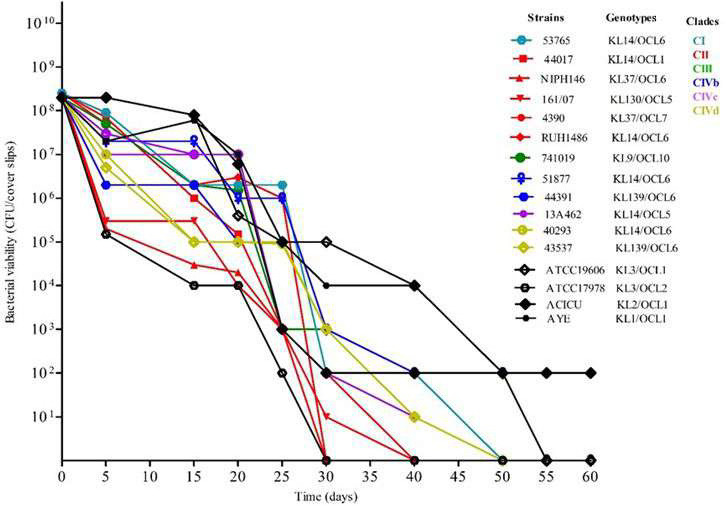 Fig 4
Fig 4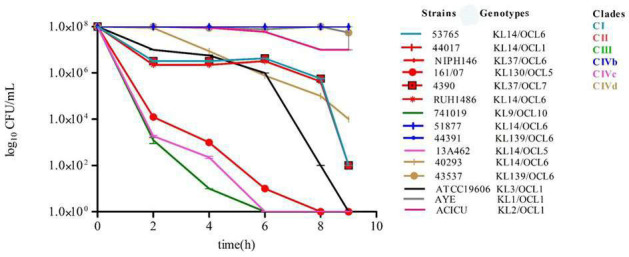 Fig 5
Fig 5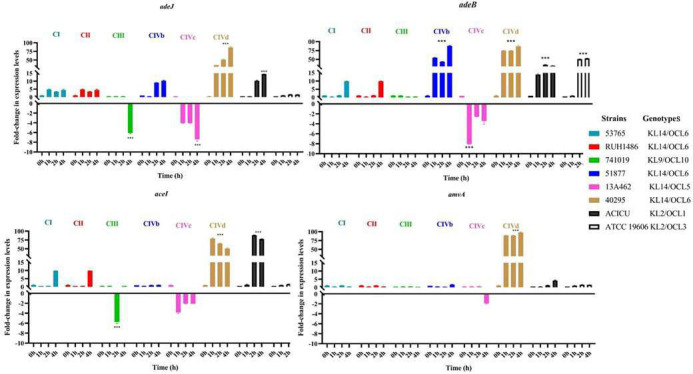 Fig 6
Fig 6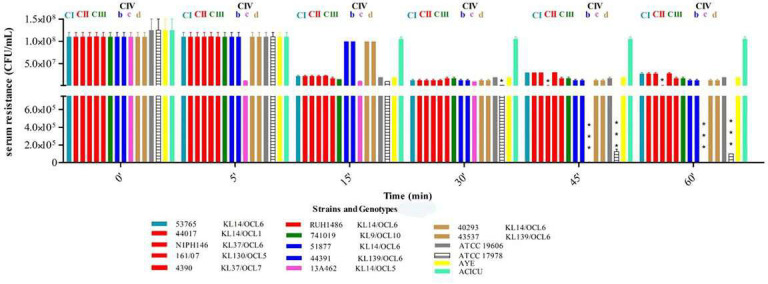 Fig 7
Fig 7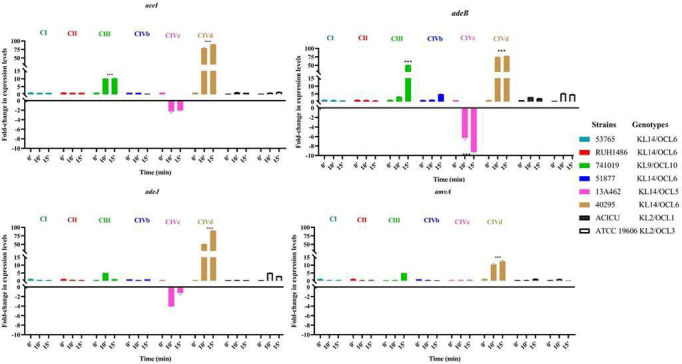 Fig 8
Fig 8| Strain | LD90 | LD50 | R^2 | Genotype | |
|---|---|---|---|---|---|
| 53765 | 1,9x10^6 | 1x10^7 | 0,877 | CI ST25 | KL14/OCL6 |
| 4390 | 1,5x10^8 | 2,6x10^7 | 0,785 | CII ST25 | KL37/OCL7 |
| 44017 | 1,9x10^6 | 1x10^7 | 0,777 | CII ST25 | KL14/OCL1 |
| 161/07 | 2x10^8 | 1x10^7 | 0,845 | CII ST25 | KL130/OCL5 |
| NIPH146 | 4x10^8 | 1x10^8 | 0,775 | CII ST25 | KL37/OCL6 |
| RUH1486 | 1x10^7 | 5x10^6 | 0,821 | CII ST25 | KL14/OCL6 |
| 741019 | 1x10^8 | 1,2x10^7 | 0,789 | CIII ST25 | KL9/OCL10 |
| 26293 | 1x10^7 | 9x10^6 | 0.825 | CIVb ST25 | KL14/OCL6 |
| 27024 | 2,2x10^6 | 1x10^6 | 0,881 | CIVb ST25 | KL14/OCL6 |
| 27027 | 2,1x10^6 | 1x10^6 | 0,777 | CIVb ST25 | KL14/OCL6 |
| 27603 | 4x10^7 | 1x10^7 | 0,888 | CIVb ST25 | KL14/OCL6 |
| 39518 | 6x10^5 | 1x10^6 | 0,744 | CIVb ST25 | KL14/OCL6 |
| 43344 | 2x10^6 | 1x10^6 | 0,912 | CIVb ST25 | KL14/OCL6 |
| 43536 | 1,5x10^6 | 9x10^6 | 0,778 | CIVb ST25 | KL14/OCL6 |
| 44391 | 9x10^7 | 2x10^7 | 0,699 | CIVb ST25 | KL139/OCL6 |
| 46732 | 1,5x10^6 | 1x10^7 | 0,811 | CIVb ST25 | KL14/OCL6 |
| 47244 | 4x10^6 | 1x10^6 | 0,712 | CIVb ST25 | KL14/OCL6 |
| 48031 | 1x10^8 | 6x10^7 | 0,846 | CIVb ST25 | KL14/OCL6 |
| 48427 | 1x10^7 | 8x10^6 | 0,835 | CIVb ST25 | KL14/OCL6 |
| 51877 | 1x10^7 | 1x10^6 | 0,745 | CIVb ST25 | KL14/OCL6 |
| 53778 | 2x10^6 | 1x10^6 | 0,755 | CIVb ST25 | KL14/OCL6 |
| 53828 | 1x10^7 | 5x10^6 | 0,788 | CIVb ST25 | KL14/OCL6 |
| 53895 | 9x10^7 | 4x10^7 | 0,689 | CIVb ST25 | KL14/OCL6 |
| 53899 | 1,7x10^6 | 1x10^7 | 0,899 | CIVb ST25 | KL139/OCL6 |
| 14A543 | 1x10^8 | 2x10^7 | 0,854 | CIVc ST25 | KL14/OCL5 |
| 13A462 | 1x10^8 | 3x10^7 | 0,875 | CIVc ST25 | KL14/OCL5 |
| 15A1044 | 1x10^8 | 1x10^7 | 0,799 | CIVc ST25 | KL14/OCL5 |
| 34792 | 1x10^8 | 9x10^7 | 0,775 | CIVd ST25 | KL14/OCL6 |
| 40104 | 5x10^6 | 1x10^6 | 0,888 | CIVd ST25 | KL14/OCL6 |
| 40293 | 4x10^7 | 1x10^7 | 0,754 | CIVd ST25 | KL14/OCL6 |
| 41133 | 2x10^6 | 1x10^6 | 0,887 | CIVd ST25 | KL14/OCL6 |
| 41833 | 4x10^6 | 1x10^6 | 0,789 | CIVd ST25 | KL14/OCL6 |
| 43537 | 9x10^6 | 6x10^6 | 0,845 | CIVd ST25 | KL139/OCL6 |
| 46738 | 1x10^7 | 1x10^6 | 0,689 | CIVd ST25 | KL14/OCL6 |
| 44772 | 6x10^6 | 1x10^6 | 0,778 | CIVd ST25 | KL14/OCL6 |
| 48420 | 1x10^7 | 9x10^6 | 0,884 | CIVd ST25 | KL14/OCL6 |
| 48422 | 6x10^7 | 1x10^7 | 0,845 | CIVd ST25 | KL14/OCL6 |
| 51874 | 2x10^6 | 1x10^6 | 0,788 | CIVd ST25 | KL14/OCL6 |
| 16A1524 | 1x10^7 | 6x10^6 | 0,811 | CIVd ST25 | KL14/OCL6 |
| 17A1650 | 1x10^7 | 8x10^6 | 0,847 | CIVd ST25 | KL14/OCL6 |
| AYE | 4x10^7 | 2x10^7 | 0,789 | ST1 | KL1/OCL1 |
| ACICU | 9x10^5 | 1x10^5 | 0,845 | ST2 | KL2/OCL1 |
| ATCC 19606 | 3x10^7 | 1x10^7 | 0,874 | ST52 | KL3/OCL1 |
- —Ministero dell'Università e della Ricercahttp://dx.doi.org/10.13039/501100021856
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInvertebrate Immune Response Mechanisms · Coccidia and coccidiosis research · Malaria Research and Control
INTRODUCTION
Acinetobacter baumannii is an opportunistic pathogen responsible for healthcare-associated infections occurring worldwide. Less frequently, A. baumannii has been associated with community-acquired infection in fragile patients (1). These infections are difficult to treat because the majority of A. baumannii isolates are resistant to at least three different classes of antimicrobials, including carbapenems, and retain susceptibility to colistin only (1). Because of this, the World Health Organization classifies carbapenem-resistant A. baumannii as a “critical” pathogen among the antibiotic-resistant bacteria of global priority (2).
While the hospital environment has been considered the primary reservoir of A. baumannii (1, 3), additional sources such as animals, food, poultry livestock, soil, plants, freshwater, sludge, and wastewater were identified (4–10).
The global epidemiology of A. baumannii isolates has shown that the population structure of the species was characterized by the selection of genetically distinct clonal lineages, three international clonal lineages (ICL) I, II, and III, and up to eight additional epidemic clonal lineages (11–13). The ST25 A. baumannii clone, according to Pasteur’s MLST scheme (14), emerged as a globally distributed multidrug-resistant lineage responsible for epidemics in Europe, South America, Africa, and Asia (15–17). Isolates belonging to the ST25 lineage have also been reported from infections in animals, in particular urinary tract infections, and in veterinary clinics (5, 8, 10). The phylogenetic analysis of ST25 A. baumannii lineage identified four clades (CI–CIV) with different geographical distribution. In particular, isolates belonging to CI and CIII clades originated from South America, whereas those included in CII and CIV clades have global geographical origin (5). ST25 A. baumannii was also isolated from chick-box papers of turkey chicks (6). Further studies demonstrated that ST25 A. baumannii isolates possess specific capsular polysaccharide loci (KL) and lipooligosaccharide outer core locus (OCL) types and show a high variability in these virulence determinants (18, 19).
In addition to resistance to a broad range of antimicrobials, A. baumannii epidemic clonal lineages possessed virulence-related traits, such as biofilm growth, adherence to host epithelial cells, resistance to desiccation, resistance to oxidative stress, and the ability to kill Galleria mellonella larvae, which contributed to replication into infected hosts and persistence in contaminated environments (3, 20). Since knowledge on specific virulence traits of the ST25 lineage is lacking, the present study aimed to (i) update the phylogenetic analysis and KL and OCL typing of ST25 A. baumannii; (ii) identify the genotypic virulence profiles of 203 ST25 A. baumannii genomes from human, animal, and environmental isolates; (iii) analyze phenotypic virulence-related and stress-related traits of 40 representative ST25 A. baumannii isolates; and (iv) correlate genotypic virulence profiles with phenotypic virulence-related and stress-related traits of ST25 A. baumannii isolates.
MATERIALS AND METHODS
Database of genomes and phylogeny
A collection of 203 A. baumannii genomes, all assigned to ST25 sequence type according to the Pasteur MLST scheme (14), was assembled from (i) the data set (n = 148) previously described by Lupo et al. (5) and (ii) genomes (n = 55) manually selected from the PubMLST database (21) up to 1 May 2024. The characteristics of the genomes were included in Table S1 and are available as a public project: pubmlst.org/bigsdb?db=pubmlst_abaumannii_isolates&page=project&project_id=23. The reference strains A. baumannii AYE (ST1), ATCC 19606 (ST52), ATCC 17978 (ST437), and ACICU (ST2) were also included in the study (22–25). The core-genome analysis was undertaken based on 2,386 conserved genes defined within the A. baumannii cgMLST scheme (https://github.com/bvalot/pyMLST). Phylogenetic reconstruction was carried out using MrBayes version 3.2.7, applying the GTR + G + I nucleotide substitution model (26). The resulting phylogenetic tree was midpoint-rooted and visualized using Interactive Tree of Life (iTOL) version 6 (27).
In silico typing of virulence factors associated with ST25 isolates
A comprehensive screening of virulence-associated genes was performed across all 203 ST25 A. baumannii genomes. We used the pubMLST software (21) to develop A. baumannii-specific virulence genes database. Our curated list comprised 127 genes, selected from three main sources: the ResFinder database (six genes belonging to effector delivery system or enzymes categories), the PubMLST virulence gene collection (111 genes belonging to biofilm, adherence, EPs, quorum sensing, metabolism/nutrition, or immune modulation categories), biofilm-associated proteins (Baps)1-2-3 as described by De Gregorio et al. (28), and genes involved in iron acquisition and metabolism previously characterized by Artuso et al. (29). The virulence genes were grouped and classified in functional categories and are available on pubMLST (https://pubmlst.org/bigsdb?db=pubmlst_abaumannii_isolates&page=plugin&name=GenePresence). A threshold identity of 80%, a minimum length of 80% matches, and a coverage value of more than 90% were the values considered for establishing the presence of each virulence gene.
To further characterize the surface polysaccharide loci of each ST25 A. baumannii genome, KL and OCL typing was performed using Kaptive 2.0.4 (18, 19) and the CAGECAT platform (https://cagecat.bioinformatics.nl/)
To assess the pangenome characteristics, PPanGGOLiN v1.1.136 (30) was used with default settings to classify gene families into persistent, shell, and cloud partitions. As input, genome annotations in GFF3 format were generated using Bakta v1.8.2 (31), also executed with default parameters.
Bacterial isolates and growth conditions
Forty ST25 representative isolates available from ANSES-Laboratory Lyon and Public Health-Laboratory Naples, and the reference strains A. baumannii ACICU, AYE, ATCC 19606, and ATCC 17978 (5, 12, 15, 22–24) were used in phenotypic assays to analyze their virulence ability. To investigate the role of EPs on virulence features of ST25 A. baumannii, a set of isogenic mutants derived from the A. baumannii ATCC 19606 wild-type strain was used. These included gene deletions targeting adeB and adeJ (belonging to the RND, Resistance-Nodulation-Division family), aceI (PACE, Proteobacterial Antimicrobial Compound family), and amvA (MFS, Major Facilitator Superfamily), as previously described by Migliaccio et al. (32). All isolates were cultured under aerobic conditions at 37°C in Luria-Bertani (LB) broth/agar. LB broth/agar, phosphate-buffered saline (PBS) 1×, and tryptic soy broth (TSB) were used to perform biofilm formation, oxidative stress resistance, serum resistance, desiccation resistance, and Galleria mellonella larvae virulence assays. The chemical reagents were purchased from Sigma-Aldrich (Sigma, Milan, Italy).
Biofilm assays
Biofilm formation was assessed using the crystal violet (CV) staining assay as previously described (33, 34) and estimated according to the European Committee for Antimicrobial Susceptibility Testing (35). Bacterial cell suspensions were adjusted to 0.5 McFarland using a BD PhoenixSpec nephelometer after an overnight (O/N) growth in TSB. Subsequently, the bacterial cells were diluted to a final culture density of approximately 1 × 10^6^ colony-forming units (CFU)/mL in TSB and were transferred into a 96-well flat-bottomed polystyrene microtiter plate. The microplates were incubated at 37°C for 24 h. The culture supernatants were gently discarded, the wells were washed twice with PBS pH 7.4, and the biofilms were stained with 200 μL of 0.1% CV for 20 min. The wells were washed twice with PBS, and the linked CV was eluted with ethanol. The absorbance was measured at 595 nm using a microplate reader (Bio-Rad Laboratories S.r.l.). The OD_595_/OD_600_ ratio was used to normalize the amount of biofilm formed versus growth.
G. mellonella larvae virulence assays
A. baumannii ST25*,* ATCC 19606, ATCC 17978, ACICU, and AYE strains were grown in LB until late exponential phase (0.4–0.5 OD_600_). Cells were collected by centrifugation and suspended in PBS. G. mellonella larvae were purchased by Insect Novel Ecologic Food S.r.l, Padua, Italy. Serial 10-fold dilutions of bacterial cell suspensions in PBS 1× were injected into G. mellonella larvae as described previously (36). Ten larvae were infected with each infecting dose (from ~1 × 10^5^ to ~1 × 10^8^ CFU/mL), and 10 larvae were injected with PBS as a negative control. Larvae were incubated at 37°C for 96 h to monitor the survival. Each isolate was tested in three independent experiments. Dose-dependent survival curves and 50% (LD_50_) and 90% (LD_90_) lethal doses were determined using the GraphPad Prism software as previously described (36). The results obtained were plotted to generate Kaplan-Meier survival curves (GraphPad Prism version 10.0).
Resistance to desiccation assays
The desiccation assay was performed as previously described (20). Overnight LB cultures were centrifuged at 12,000 × g for 5 min in a microcentrifuge. The cell pellet was washed twice with PBS and suspended to 1 OD_600_. Twenty microliters of each suspension was deposited onto a glass cover slip to produce an inoculum of 1 × 10^7^ CFU/mL. The coverslip was kept at 30% + 5% relative humidity by the presence of a saturated CaCl_2_ in an uncovered Petri dish and stored at room temperature in an air-tight transparent plastic box (17 × 11 × 5.5 cm) for up to 60 days. Viable cell counts were determined by detaching and seeding on LB agar and further incubation at 37°C O/N.
Serum resistance assays
Serum resistance assays were performed as described previously by Shin et al. (37) with minor modifications. Overnight bacterial cultures were diluted 1:100 into 10 mL of fresh LB medium and incubated until the bacterial suspension reached 0.5 OD_600_. Then, 1 mL aliquot of the culture was washed with PBS and resuspended in the same quantity. Subsequently, 100 μL of the bacterial suspension was added in 96-multiwell and mixed with bovine serum diluted to 20%; the inactivated serum at 56°C for 45 min was also used. After mixing, the serum-bacteria suspensions were incubated at 37°C for 60 min. To calculate the serum bactericidal effect, a 100-µL aliquot was taken from each suspension, serially diluted, and plated on LB agar. The serum bactericidal effect was expressed as CFU/mL of viable cells.
Oxidative stress tolerance
Oxidative stress tolerance was tested by exposing 0.1 OD_600_ of O/N growth of representative clades A. baumannii ST25 strains, ATCC19606, ATCC17978, AYE, ACICU, and EPs mutant strains to 150 µM H_2_O_2_ for 9 h at 37°C under shaking. Bacteria were exposed to 50–200 µM H_2_O_2_. These concentrations were obtained by diluting in PBS from the stock solution (9.8 × 10^6^ µM at pH 4.0). To calculate the H_2_O_2_ effect, a 100 µL aliquot was taken each hour, serially diluted, and plated on LB agar, incubated at 37 °C O/N (38).
RNA purification and real-time quantitative PCR
A. baumannii strains were grown as for the serum assay or oxidative stress assays. The bacterial cells were washed with PBS, and total RNA was isolated via TRIzol reagent (Qiagen, Milan, Italy). The cDNAs were synthesized using QuantiTect Reverse Transcription Kit (Qiagen, Milan, Italy), according to the manufacturer’s protocol. Real-time quantitative PCR (RT-qPCR) assays were performed using SYBR Green master mix (Applied Biosystems) (36). The rpoB housekeeping gene was used to normalize the expression of target genes (39), and the fold changes of gene expression levels were calculated using the 2^–ΔΔct^ method (40). All experiments were performed three times in triplicate. The primers used in the RT-qPCR experiments were previously reported (32).
Statistical analysis
All statistical analyses were conducted using GraphPad Prism version 10.0 (GraphPad Software, San Diego, CA, USA). Differences among multiple groups were assessed using two-way analysis of variance (ANOVA), followed by the Bonferroni post hoc test to correct for multiple comparisons. Statistical significance was defined as P < 0.05, P<0.01, and P<0.001. Two-way ANOVA and Tukey test P-values were utilized for statistical analysis of biofilm experiments. Spearman’s rank correlation coefficient (ρ) was used to assess the relationship between the expression levels of efflux pump (EP) genes and the corresponding phenotypes of EP mutants, in comparison to the ST25 isolates. This non-parametric test was selected due to the ordinal nature of the expression data and the potential lack of normal distribution. Correlation analyses were performed to investigate potential associations between KL/OCL genotypes and phenotypic traits of virulence and stress resistance. Specifically, the contingency tables were constructed for each phenotype, and chi-square (χ²) tests were performed. Cramer’s V coefficient was calculated to quantify the strength of the associations based on degrees of freedom (df), and P-values were used to assess statistical significance (P < 0.05) (41). All data were presented as means ± standard deviation. The number of replicates was two or three times.
RESULTS
Phylogenomic structure and KL and OCL typing of ST25 A. baumannii clonal lineage
The core-genome MLST analysis of 203 A. baumannii ST25 genomes distinct clades: CI (20 genomes), CII (48 genomes), CIII (27 genomes), and CIV (108 genomes) (Fig. 1). Clade IV was further subdivided into subclades IVa (1 genome), IVb (20 genomes), IVc (31 genomes), and IVd (56 genomes) (Fig. 1).
cgMLST-based phylogeny of 203 A. baumannii ST25 genomes. The phylogenetic tree is annotated with concentric colored rings as follows: the outermost ring indicates the KL types, the second ring from the outside represents OCL types, and the innermost ring indicates clades and subclades. Distribution of KL/OCL types corresponding to the KL/OCL genotypes of isolates selected for phenotypic assays, indicated by colored rectangles outside the tree. The colors of the isolates specify the origin of isolation: 150 isolates from humans in black, 36 from animals in yellow, and 10 from environmental sources in red, 7 isolates of unknown origin in green.
In addition to the population structure analysis, this study also assessed the capsular KL and OCL profiles of ST25 A. baumannii genomes. Each genome was characterized by a specific KL and OCL combination, adding resolution to the genomic diversity within the ST25 lineage (Table S1). KL14 was the predominant capsular locus, found in 47% of genomes assigned to clades CIV (80%) and CII (20%) (Fig. 1); KL139 and KL37 were found in 10.3% and 9.8% of genomes, respectively. OCL6 was the most frequent OCL type (50%), followed by OCL5 (35%) and OCL10 (9.8%). These isolates originated from diverse sources, including humans (150 genomes), animals (36 genomes), the environment (10 genomes), and unknown (7 genomes). The human-derived isolates were distributed across all clades and displayed considerable heterogeneity in KL and OCL types (Table S1; Fig. 1). Instead, animal-derived isolates belonged to clades CII, CIVb, and CIVd and carried the KL14/OCL6 combination (33 out of 36 genomes). This KL14/OCL6 combination was especially predominant in CIVb (n=16/20) and CIVd (n=44/56) sub-clades independently of the host (Fig. 1). Environmental isolates belonged to clades CII (9 genomes) and CIVc (1 genome) and were associated with KL14/OCL5 (4 out of 9 genomes), KL37/OCL6 (4 out of 9 genomes), and KL10/OCL6 (1 out of 1 genome) (Table S1; Fig. 1). The most common KL types shared identical genes at the 5′ and the 3′ of the locus (Fig. 2A). In detail, capsular export genes wzc and wzb, but not wza, showed 100% nucleotide identity and were found at the 5′ of KL22, KL129, KL33, KL116, KL37, KL14, and KL139, while identical pgm and gne1 genes were found at the 3′ of all prevalent loci (Fig. 2A).
Comparative analysis of representative KLs (A) and OCLs (B). The colors indicating the alignment of the KL/OCL loci use an identity threshold of 0.98.
Contrarily, the OCL loci showed relatively higher structural conservation, particularly among OCL10, OCL5, and OCL6 (Fig. 2B). The glycosyltransferases encoding gene, gtrOC1, was found identical in all OCL types, while gtrOC2, gtrOC3, and gtrcOC4 were identical in OCL1 and OCL10. Also, the identical polysaccharide deacetylase pda2 gene was found in OCL6 and OCL7, a similar polysaccharide deacetylase pda2a gene was found to be associated with OCL7 and OCL5, and an identical pda1 gene was found in OCL1 and OCL10 (Fig. 2B).
In silico analysis of virulence genes in A. baumannii ST25 genomes
Among the adherence category, distinct pil- and fim- genes were present in the 203 ST25 A. baumannii genomes analyzed, while ata, pilA, and pilC were absent (Table S2). In the effector delivery system category, all genes of T2SS and some genes of the T6SS subcategories were present in ST25 genomes. In the biofilm category, the genes belonging to Poly-β−1,6-N-acetyl-D-glucosamine, quorum sensing, csu fimbriae, and bap-3 (type-3 biofilm-associated protein) encoding genes were present in all ST25 genomes. In the metabolism/nutrition category, the eight genes belonging to the hemO cluster subcategory were present in all ST25 genomes, while the bauA gene of acinetobactin subcategory was present in 7% of ST25 genomes. The adeA and adeB genes encoding the AdeABC EP system and adeI, adeJ, and adeK of the AdeIJK efflux system, along with adeN, adeL, adeR, and adeS transcriptional regulators, were present in all ST25 genomes. Among the regulation categories, the Bfmrs regulatory system is present in all ST25 genomes. Among the exoenzyme category, plcD, plc2, plc1 exotoxin genes, and cpaA gene are present in all ST25 genomes (Table S2).
ST25 A. baumannii pangenome analysis
Pangenome analysis of the 203 ST25 A. baumannii genomes generated a K3 gene family partition into persistent, cloud, and shell, based on their frequency across genomes (Fig. S1). The persistent group, comprising 3,084 genes present in all 203 genomes, was considered the core genome. The cloud (3,597) and shell (2,118) genes were identified in 1–8 and 8–198 genomes, respectively, and were considered the accessory genome (Table S3).
We carefully analyzed the shell partition, which includes moderately conserved genes across the genomes and could potentially reflect clade-specific traits. Among shell gene families, the genes encoding Fe-S hydro-lyase tartrate dehydratase beta-type catalytic domain-containing protein, tartrate dehydratase, a putative tartrate transporter, and an HTH-type transcriptional regulator DmlR were found only in CII clade genomes (or CII clade-specific) (Table S3). Also, gene families associated with phage-related functions and hypothetical proteins, including DUF domain-containing proteins and various bacteriophage tail and head components, were identified in the 129 genomes belonging to CIV (108/108 genomes) (or CIV semi-clade-specific), CII (10/53 genomes), and CIII (11/27 genomes) (Fig. S1; Table S3).
Virulence-related traits analyses of ST25 A. baumannii
Based on the in silico results, a subset of 40 ST25 A. baumannii isolates belonging to different KL/OCL genotypes was included for in vivo and in vitro analyses of the most common virulence traits in A. baumannii. The selected ST25 A. baumannii isolates were assigned to the following genotypic capsular profiles: KL14/OCL6 (one isolate) assigned to clade CI, KL14/OCL6 (two isolates), KL14/OCL1 (1), KL37/OCL6 (1), KL37/OCL7 (1), and KL130/OCL5 (1) for isolates assigned to clade CII; KL9/OCL10 for clade CIII (one isolate); KL14/OCL6 (16) and KL139/OCL6 (1) for subclade CIVb; KL14/OCL5 (3) for subclade CIVc; and KL14/OCL6 (14) for subclade CIVd (Tables S1 and S4).
Virulence profiles of ST25 A. baumannii strains in G. mellonella larvae
The virulence potential of ST25 A. baumannii selected isolates was analyzed using the Galleria mellonella infection model and results compared with those of AYE, ACICU, and ATCC 19606 A. baumannii strains (Table 1; Fig. S2). Dose-dependent LD_50_ and LD_90_ values in G. mellonella larvae showed distinct virulence profiles among ST25 A. baumannii strains belonging to the different clades and KL/OCL types. In particular, CIVd and CIVb strains showed a high infectivity with LD_50_ and LD_90_ values ranging from 1 × 10^6^ to 4 × 10^7^ CFU/mL and 2 × 10^6^ to 6 × 10^7^ CFU/mL, respectively (Table 1). In contrast, LD_50_ and LD_90_ of CI, CII, CIII, and CIVc strains were 3.33- and 5-fold higher than LD_50_ and LD_90_ of CIVd and CIVb strains (P < 0.01). The virulence profiles of CIVd and CIVb strains were similar to those of AYE and ATCC 19606 strains, which displayed LD_50_ and LD_90_ values ranging from 1 × 10^7^ CFU/mL to 4 × 10^7^ CFU/mL, while they were lower than that of the ACICU strain, which showed LD_50_ and LD_90_ values of 1 × 10^5^ CFU/mL and 9 × 10^5^ CFU/mL, respectively. Moreover, a significant association was found between KL/OCL genotypes and virulence in G. mellonella, with nearly all (n = 22/29) KL14/OCL6 genotypes being highly virulent (Cramer’s V = 0.94, χ² = 35.4, df = 7, P = 9.4 × 10⁻⁶).
Biofilm formation ability of ST25 A. baumannii
The biofilm growth of ST25 A. baumannii strains was analyzed using the ATCC 19606 strain as a positive control (42), and AYE and ATCC 17978 as negative controls (15, 43) (Fig. 3). Among the 40 ST25 isolates, 19 (1/1 CI, 1/5 CII, 1/1 CIII, 4/17 CIVb, 3/3 CIVc, and 9/14 CIVd isolates) showed 1.5- to 2.5-fold higher biofilm formation than AYE and ATCC17978 controls, but less biofilm formation than ATCC19606 (Fig. 3) and were classified as intermediate biofilm producers (43).
*Biofilm formation of ST25 A. baumannii strains. The data were obtained from three independent experiments in which each isolate was tested in triplicate. The significance of the differences was calculated using two-way ANOVA (*P-values < 0.5 , **P-values <0.01, and **P-values <0.001). The isolates’ names and their respective KL/OCL genotypes are shown at the bottom of the graph.
Our data showed a high degree of heterogeneity in biofilm formation of ST25 A. baumannii isolates assigned to different clades, while identifying a correlation between specific KL/OCL genotypes and biofilm formation among the 40 ST25 A. baumannii isolates. In particular, biofilm production was strongly associated with KL14/OCL5 and KL139/OCL6 genotypes, while KL14/OCL6 isolates were biofilm-non-producer (Cramer’s V = 0.85, χ² = 28.7, df = 7, P = 0.00016) (Fig. 3; Table S4).
Resistance to desiccation of ST25 A. baumannii strains
The ability of ST25 A. baumannii strains to survive under desiccating conditions on abiotic surfaces was evaluated and compared with that of ATCC 19606, AYE, ACICU, ATCC 17978 A. baumannii strains, and ATCC 19606 EPs mutant strains. Strains from clades CI, CII, and CIII exhibited notable variability in survival, with several CII isolates, such as 44017 and 4390, retaining viability up to 30 days, while others declined more rapidly (Fig. 4). Interestingly, the CIII isolate 741019 demonstrated higher resistance to desiccation than CI and CII strains and maintained viability beyond 50 days (Fig. 4).
Desiccation resistance of ST25 A. baumannii strains. The viable cells (CFU/mL) were determined for each isolate after incubation on glass slips in a 30% ± 5% humidity environment. The data were obtained from three independent experiments in which each isolate was tested in triplicate.
Clade CIVb strains showed more consistent and prolonged survival, with many isolates retaining detectable CFUs up to 50–60 days. Similarly, isolates from clade CIVd showed resistance to desiccation, with several strains, including 16A1524 and 40293, maintaining viability well beyond 40 days. Similarly, clade CIVc strains displayed survival until 50 days (Fig. 4). Collectively, these findings highlighted considerable variability in desiccation tolerance across A. baumannii clades and strains. ST25 A. baumannii strains showed similar desiccation resistance compared with ATCC 19606, AYE, ACICU, and higher desiccation resistance compared with ATCC 17978 (Fig. 4). In particular, AYE and ACICU strains showed the highest survival at 60 days, with 10^4^ and 10^2^ CFU/mL, respectively. In contrast, ATCC 17978 showed a rapid decline, with no viable cells detected beyond day 20 (Fig. 4). Also, ST25 strains assigned to KL14/OCL6, KL139/OCL6, and genotypes consistently displayed long survival to desiccation, while KL9/OCL10 was susceptible (χ² = 40.0, df = 7, P = 1.26 × 10⁻⁶; V = 1.00) (Fig. 4; Table S4).
To investigate the contribution of EPs to desiccation survival in A. baumannii, the ability of ATCC 19606 A. baumannii and ATCC 19606 EPs mutant strains to survive under desiccating conditions was investigated. The parental strain ATCC 19606 survived up to 60 days, whereas EP mutants displayed differential desiccation tolerance (Fig. S3). The ΔadeB strain notably showed impaired long-term survival, with CFUs dropping sharply after day 50. The Δamva, Δacel, and ΔadeJ strains exhibited markedly reduced survival in CFU/mL terms, compared with wild type or ΔadeB, becoming undetectable around day 50 (Fig. S3).
Oxidative stress tolerance and expression of EP genes under oxidative stress in ST25 A. baumannii
We evaluated the ability of ST25 A. baumannii strains to tolerate oxidative stress following exposure to 150 µM H_2_O_2_. Strains 51877 and 44391, belonging to CIVb, displayed strong resistance to 150 µM H_2_O_2_, maintaining high CFU levels (10^8^ CFU/mL) over 8 h. In contrast, 741019 of CIII, 13A462 strains belonging to CIVc showed higher sensitivity than CIVb strains and rapidly declined below the detection limit within 2–6 h. Furthermore, CI, CII, and CIVd strains showed 10^7^ CFU/mL until 6 h, and then the viability decreased to 10^4^–10^2^ CFU/mL (Fig. 5). These data indicated substantial heterogeneity among ST25 A. baumannii isolates in oxidative stress response. Similarly to desiccation resistance, ST25 A. baumannii strains assigned to KL14/OCL6 and KL139/OCL6 genotypes consistently displayed high resistance to H₂O₂, while those assigned to KL14/OCL5, KL130/OCL5, and KL9/OCL10 genotypes were generally susceptible (χ² = 40.0, df = 7, P = 1.26 × 10⁻⁶; V = 1.00) (Fig. 5; Table S4).
Oxidative stress tolerance of ST25 A. baumannii strains. The viable cells (CFU/mL) were determined for each isolate after 0–9 h of 150 µM H₂O₂ exposure. The data were obtained from three independent experiments in which each isolate was tested in triplicate.
To study the role of EPs in oxidative stress tolerance in ST25 A. baumannii, ATCC 19606 A. baumannii, and ATCC 19606, EP mutant strains were included. The targeted deletions in EP genes (∆aceI, ∆amvA, ∆adeJ, and ∆adeB) impaired H₂O₂ survival compared to the wild-type ATCC19606. Notably, ∆adeJ and ∆adeB strains were highly susceptible, counting 10^4^ or 10^2^ CFU/mL cells viable after 2 h of exposure (Fig. S4). These results suggested that these efflux systems contributed directly or indirectly to oxidative stress protection, potentially by exporting toxic compounds or maintaining redox balance. The expression of EPs genes in ST25 A. baumannii isolates was thus assessed under exposure to 150 mM H₂O₂. All Ep gene expressions were significantly upregulated at 1, 2, and 4 h in isolate 40293 belonging to CIVd; adeJ and adeB genes were significantly upregulated at 2 and 4 h in isolate 51788 belonging to CIVb. In contrast, aceI and amvA were downregulated but not significantly in 13A462 assigned to CIVc; adeB and adeJ genes were significantly downregulated, and aceI gene expression was downregulated but not significantly in 13A462 strain belonging to CIVc (Fig. 6).
*Fold change in expression levels of adeJ, adeB, aceI, and amvA EPS genes in ST25 A. baumannii after 0, 1, 2, and 4 h of H2O2 exposure. The significance of the differences was calculated using two-way ANOVA (**P-values <0.01, **P-values < 0.001).
The expression profiles and mutant phenotypes suggested a link between the upregulation of adeB, adeJ, and aceI gene expression and the ability of ST25 A. baumannii isolates to tolerate oxidative stress. Notably, the high tolerance to oxidative stress of CIVd and CIVb isolates correlated with adeB and adeJ overexpression (Spearman p-value corresponding to 0.0173 and 0.0166, respectively).
Serum resistance and expression of genes encoding efflux systems under serum exposure in ST25 A. baumannii
The majority of ST25 A. baumannii isolates within CI, CII, CIII, CIVb, and CIVd exhibited high levels of serum resistance, maintaining bacterial viability (10^7^–10^8^ CFU/mL) even after 60 min of exposure. In contrast, isolates belonging to CIVc showed a significant decline in viability over time, even after 45-min exposure to serum. Furthermore, ST25 A. baumannii isolates assigned to KL14/OCL6 and KL139/OCL6 consistently displayed high resistance to serum treatment, while KL14/OCL5 and KL9/OCL10 were more sensitive (χ² = 40.0, df = 7, P = 1.26 × 10⁻⁶; V = 1.00) (Fig. 7; Table S4).
*Serum resistance of ST25 A. baumannii, ST2 ACICU, ST1 AYE, and ST52 ATCC 19606. The viable cells (CFU/mL) were determined for each isolate following a 5- to 60-min incubation in 20% activated serum and normalized using values obtained from incubation with heat-inactivated. The data were obtained from three independent experiments in which each isolate was tested in triplicate. **P-values <0.001.
Considering that ATCC 19606 isogenic deletion mutants of adeB, adeJ, aceI, and amvA EPs genes displayed significantly reduced serum resistance compared to the wild-type strain (Fig. S5), we investigated whether EPs played a role in serum resistance of ST25 A. baumannii isolates and whether expression of EP genes was modulated during serum exposure. In CIVd isolates, transcripts of adeB, adeJ, and aceI genes were 80-fold higher (P < 0.001) compared to ATCC 19606 (Fig. 8). Furthermore, the amvA gene was upregulated up to 15-fold compared with ATCC 19606. Similarly, Clade III isolate 741019 exhibited 4- and 10-fold upregulation of aceI and adeB genes, respectively. In contrast, the expression of aceI and adeB genes in CIVc isolates was downregulated by 2- and 6-folds, respectively, compared with ATCC19606 A. baumannii (Fig. 8). No statistical correlation was found between the expression of adeB, adeJ, aceI, and amvA genes and resistance to serum exposure in ST25 A. baumannii isolates (Sperman P-values corresponding to 0.667, 0.677, 0.755, and 0.371, respectively).
*Fold change in expression levels of adeJ, adeB, aceI, and amvA EPS genes in ST25 A. baumannii after 0, 10, or 15 min of serum exposure. The significance of the differences was calculated using two-way ANOVA (**P-values < 0.01, **P-values < 0.001).
DISCUSSION
A. baumannii strains assigned to ST25 epidemic clonal lineage have been increasingly isolated worldwide from humans (5, 7, 15, 44), animals (5, 8), and the environment (6, 7) and were selected because of a high number of acquired and/or chromosomally embedded antimicrobial resistance genes (4–8, 15). In accordance with previous studies (5, 15), the core-genome MLST phylogeny of 203 ST25 isolates from human, animal, and environmental sources analyzed herein showed genomic diversity and distribution of genomes in four clades (CI–CIV) and four sub-clades (CIVa–CIVd). Corroborating previous publication by Lupo et al. on 141 ST25 A. baumannii isolates (5), the data of the present study showed geographical clustering, with CI and CIII clades over-representing isolates from South America, while isolates from CII and CIV clades came from all over the world. Additional epidemiological information was provided by KL and OCL. The capsular polysaccharides KL14, KL139, KL37, and KL116, which are a group of related structures found in ST25 A. baumannii isolates (45, 46), were identified in 47%, 10%, 7.4%, and 2% of ST25 A. baumannii isolates, respectively. The oligosaccharides OCL6, OCL5, and OCL10 were found in 50%, 35%, and 9.8% of ST25 A. baumannii isolates, respectively. In accordance with previous studies (18, 19), ST25 A. baumannii isolates included in this study showed an elevated variability of KL and OCL types. Nevertheless, KL14/OCL6 and KL14/OCL5 were the most prevalent KL and OCL types and were found in 61 and 11 out of 203 genomes, respectively. In keeping with our data, KL116/OCL6 and KL14/OCL6 genotypes are found in ST25 A. baumannii clinical isolates from Nigerian hospitals (16) and ST25 A. baumannii isolates from French companion animals (10), respectively.
Increasing evidence indicates that A. baumannii strains belonging to the ICLs I–III and to additional epidemic clonal lineages, including the ST25 lineage, possess peculiar virulence features, which sustain infection of animated hosts and survival in the environment (3, 20). The analysis of virulence genes identified in the core and shell genome of A. baumannii shows that all 203 ST25 A. baumannii genomes possess an extended panel of virulence genes belonging to biofilm, adherence, EPs, quorum sensing, metabolism/nutrition, immune modulation, and exotoxin categories. The analysis of the accessory genome of ST25 A. baumannii isolates showed the presence of tartrate metabolism genes only in CII genomes and gene families associated with phage-related functions in CIV genomes, which are absent or rare in other clades. Clade IV semi-specific genes were mostly phage derived and may influence surface structures, prophage dynamics, and host interactions, supporting the hypothesis that accessory elements contribute to the evolutionary diversification of this clade. Interestingly, ST25 A. baumannii genomes constituting the CIV clade were also the most recent and carried multiple resistance genes, including carbapenemase-encoding genes (5).
Further information on the virulence features of ST25 A. baumannii isolates was obtained by the phenotypic virulence-related and stress-related traits of a representative data set of 40 ST25 A. baumannii strains selected among the most prevalent genotypic profiles of ST25 genomes (Table S4). The analysis of virulence profiles in the G. mellonella model demonstrated that ST25 A. baumannii strains possess an intermediate/high infectivity, with CIVb and CVd strains being more virulent than strains assigned to other clades. The ability of ST25 CIVb and CIVd strains to kill G. mellonella larvae is similar to that of AYE A. baumannii strain assigned to ICL-I and to ATCC19606 (ST52) A. baumannii reference strains, while it is lower than that of ACICU strain A. baumannii assigned to ICL-II. Moreover, ST25 A. baumannii strains with KL14/OCL6 genotype showed higher ability to infect G. mellonella larvae than strains with other capsular types.
Virulence-related traits such as biofilm formation on abiotic surfaces, tolerance to oxidative stress, and resistance to desiccation contribute to the survival and spread of A. baumannii in the contaminated environment (3). In accordance with previous data (15, 43), ST25 A. baumannii isolates were intermediate biofilm producers but showed higher biofilm formation than the AYE strain and the ATCC17978 biofilm non-producer reference strain. A high biofilm production was observed in ST25 A. baumannii isolates with KL14/OCL5 and KL139/OCL6 genotypes, suggesting that the above KL/OCL loci may play a role in biofilm formation.
Our study also showed that ST25 A. baumannii strains, similarly to A. baumannii strains AYE (ICL-I), ACICU (ICL-II), and ATCC 19606 reference strains, possessed an elevated ability to survive desiccation on abiotic surfaces. While strains assigned to clades CI and CII exhibited variability in desiccation survival from 30 to 50 days, the single CIII isolate, and all isolates assigned to clades CIVb, CIVc, and CIVd showed desiccation survival up to 50–60 days. This enhanced desiccation resistance suggests that isolates assigned to clades CIVb-c-d possess adaptive mechanisms that promote long-term persistence on dry surfaces. The above findings are in agreement with previous data showing that ST25 A. baumannii strains possess elevated but variable resistance to desiccation (20), showing a similar phenotype to that found for AYE and ACICU strains (20, 47). Our data also demonstrate that ST25 A. baumannii isolates with KL14 and KL139 capsular polysaccharides have elevated resistance to desiccation compared to those displaying KL37, KL9, and KL130. This suggests that distinct capsule structures play a role in resistance to desiccation, as indicated by previous publications (48, 49).
In agreement with a previous study (38), A. baumannii ST25 strains possessed the ability to resist oxidative stress. The data of the present study demonstrated that tolerance to H_2_O_2_ differed according to the genotype of strains, CIVb and CIVd strains showing higher levels of resistance than those assigned to other clades and subclades. Furthermore, ST25 strains with KL14 and KL9 capsular loci possessed higher resistance to oxidative stress compared to those presenting the KL139, KL37, and KL130 capsular loci. ST25 A. baumannii isolates assigned to different clades were similarly resistant to serum exposure, except for isolates assigned to clade CIVc, which displayed significantly reduced serum resistance. On the other hand, ST25 A. baumannii strains with KL14/OCL6 and KL139/OCL6 genotype showed higher resistance to serum exposure than strains with KL14/OCL5 and KL9/OCL10 capsular type.
Overall, our data demonstrated that phenotypic virulence-related and stress-related traits of ST25 A. baumannii isolates depend on their KL and OCL genotypic profiles. In detail, the KL14/OCL6 profile emerged as a dominant, highly virulent, and multi-stress resistant genotype, KL14/OCL5 and KL9/OCL10 were specialized in biofilm formation, and KL139/OCL6 combined biofilm formation with environmental resilience (Table S4).
Previous studies demonstrated that the activation of EPs belonging to PACE, RND, and MFS efflux systems in A. baumannii modulates susceptibility to antimicrobials and biocides (50–52) as well as stress response, biofilm formation, and virulence (50–52). In support of this, the data shown herein demonstrated that ΔadeB and ΔadeJ (RND), Δamva (MFS), and Δacel (PACE) ATCC 19606 mutants possess reduced long-term survival to desiccation compared with the ATCC19606 wild-type strain, the maximum effect being observed in the ΔadeB mutant. Similarly, ∆adeB, ∆adeJ, ∆aceI, and ∆amvA ATCC 19606 mutants showed reduced tolerance to H₂O₂ treatment, with ∆adeJ and ∆adeB strains being highly susceptible to oxidative stress. The data presented herein are consistent with the study of Srinivasan et al. (53), which demonstrates that inactivation of AbaO TolC-like protein (AbaO), an outer membrane protein often associated with RND-type efflux families in Acinetobacter spp. (50), increases sensitivity to oxidative stress challenge in A. baumannii strain AYE (53). The role of EPs in response to H₂O₂ treatment of ST25 A. baumannii strains was corroborated by the finding that gene expression of all EPs is upregulated in CIVb and CIVd isolates, which were ST25 A. baumannii strains highly tolerant to oxidative stress. Furthermore, a statistically significant correlation was found between adeB and adeJ expression and tolerance to oxidative stress in ST25 A. baumannii strains. We speculate that these efflux systems contribute directly or indirectly to oxidative stress protection, potentially by exporting toxic compounds or maintaining redox balance.
Conclusions
A. baumannii epidemic clonal lineages are selected because of multidrug resistance and virulence and stress tolerance mechanisms, which contribute to infection in human and animal hosts and environmental persistence (3, 20). A. baumannii strains belonging to ST25 epidemic clonal lineage possess similar virulence profiles in G. mellonella infection model compared with A. baumannii strain AYE assigned to international clone I, but lower virulence profiles compared with A. baumannii strain ACICU assigned to international clone II. On the other hand, ST25 A. baumannii shows high resistance to desiccation, oxidative stress, and serum treatment, similarly to A. baumannii AYE and ACICU strains assigned to international clones I and II, respectively. ST25 A. baumannii strains showed an elevated heterogeneity in phenotypic profiles of virulence and stress resistance among CI–CIV and within the same clade. Also, virulence-related and stress-related traits of ST25 A. baumannii correlated with KL14, KL9, and KL139 types, OCL5, OCL6, and OCL10 types. Furthermore, the increased expression of AdeB and AdeJ RND EPs correlated with resistance to oxidative stress and serum in KL14/OCL6 ST25 CIVb and KL14/OCL6 ST25 CIVd A. baumannii isolates, suggesting that EPs regulate tolerance to stresses and survival in the contaminated environment.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wong D, Nielsen TB, Bonomo RA, Pantapalangkoor P, Luna BM, Spellberg B. 2017. Clinical and pathophysiological overview of Acinetobacter infections: a century of challenges. Clin Microbiol Rev 30:409–447. doi:10.1128/CMR.00058-1627974412 PMC 5217799 · doi ↗ · pubmed ↗
- 2World Health Organization. 2024. Bacterial priority pathogens list, 2024. Bacterial Pathogens of Public Health Importance to Guide Research, Development and Strategies to Prevent and Control Antimicrobial Resistance. WHO, Geneva.
- 3Lucidi M, Visaggio D, Migliaccio A, Capecchi G, Visca P, Imperi F, Zarrilli R. 2024. Pathogenicity and virulence of Acinetobacter baumannii: factors contributing to the fitness in healthcare settings and the infected host. Virulence 15:2289769. doi:10.1080/21505594.2023.228976938054753 PMC 10732645 · doi ↗ · pubmed ↗
- 4Hamidian M, Maharjan RP, Farrugia DN, Delgado NN, Dinh H, Short FL, Kostoulias X, Peleg AY, Paulsen IT, Cain AK. 2022. Genomic and phenotypic analyses of diverse non-clinical Acinetobacter baumannii strains reveals strain-specific virulence and resistance capacity. Microb Genom 8:000765. doi:10.1099/mgen.0.00076535166651 PMC 8942024 · doi ↗ · pubmed ↗
- 5Lupo A, Valot B, Saras E, Drapeau A, Robert M, Bour M, Haenni M, Plésiat P, Madec J-Y, Potron A. 2023. Multiple host colonization and differential expansion of multidrug-resistant ST 25-Acinetobacter baumannii clades. Sci Rep 13:21854. doi:10.1038/s 41598-023-49268-x 38071225 PMC 10710421 · doi ↗ · pubmed ↗
- 6Schmitz A, Hanke D, Lüschow D, Schwarz S, Higgins PG, Feßler AT. 2023. Acinetobacter baumannii from samples of reared turkeys: genomic relationships, antimicrobial and biocide susceptibility. Microorganisms 11:759. doi:10.3390/microorganisms 1103075936985332 PMC 10052703 · doi ↗ · pubmed ↗
- 7Sykes EME, Mateo-Estrada V, Engelberg R, Muzaleva A, Zhanel G, Dettman J, Chapados J, Gerdis S, Akineden Ö, Khan IUH, Castillo-Ramírez S, Kumar A. 2024. Phylogenomic and phenotypic analyses highlight the diversity of antibiotic resistance and virulence in both human and non-human Acinetobacter baumannii. m Sphere 9:e 0074123. doi:10.1128/msphere.00741-2338440986 PMC 10964423 · doi ↗ · pubmed ↗
- 8Chanchaithong P, Leelapsawas C, Sroithongkham P, Yindee J, Thummeepak R, Collaud A, Perreten V. 2025. Localization of Aba R 4-type genomic islands and multidrug resistance plasmids in multiple Acinetobacter baumannii clones and Acinetobacter pittii from infections of dogs and cats. Infect Genet Evol 133:105782. doi:10.1016/j.meegid.2025.10578240490126 · doi ↗ · pubmed ↗