Catalytic Tunnel Dynamics Explain Functional Divergence of FAD2 Homologs Underlying the Oleic-to-Linoleic Ratio in Cornus wilsoniana
Wenyan Yuan, Wei Wu, Shushan Jia, Yunzhu Chen, Peiwang Li, Yan Yang, Lijuan Jiang, Qiang Liu, Li Li, Wenbin Zeng, Xiao Zhou, Jingzhen Chen

TL;DR
Researchers studied how different versions of an enzyme in a woody oil tree affect the ratio of two fatty acids in its oil, using simulations to track how the enzyme's structure changes during binding.
Contribution
The study introduces catalytic tunnel tracking to explain functional differences among FAD2 homologs in Cornus wilsoniana.
Findings
One FAD2 version lacks a key protein segment and has a damaged binding pocket, likely rendering it nonfunctional.
Two FAD2 versions form stable, open tunnels for efficient substrate positioning, while a third shows tunnel narrowing and substrate readjustments.
CW02G01750 is identified as the leading structural candidate for regulating the oleic-to-linoleic acid ratio.
Abstract
High-quality edible oils usually contain more oleic acid and less linoleic acid, but the molecular reasons for this balance remain unclear in the woody oil tree Cornus wilsoniana. We measured the main fatty acids in its fruit oil and then compared four closely related versions of an enzyme called fatty acid desaturase 2, which converts oleic acid into linoleic acid. One version lacked a large protein segment and showed a damaged binding pocket, suggesting it is unlikely to function properly. For the remaining three versions, we used molecular dynamics simulations together with catalytic tunnel tracking, a way to follow the passageway that guides the fatty acid to the reactive center, to watch how the protein shape changes during binding. Two versions formed a stable, more open access tunnel that kept the substrate well positioned, whereas the third showed higher internal flexibility and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
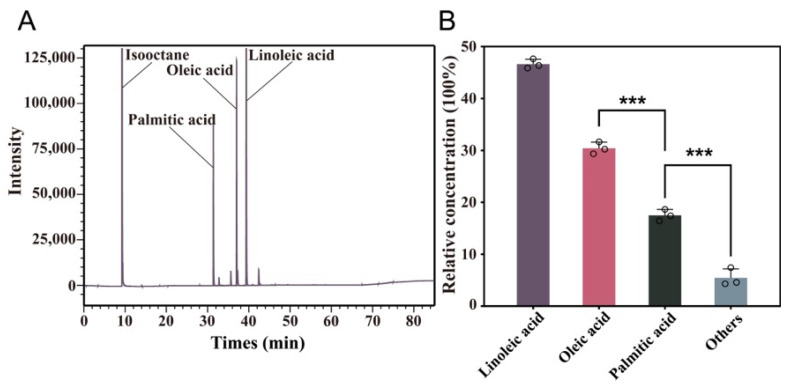 Figure 1
Figure 1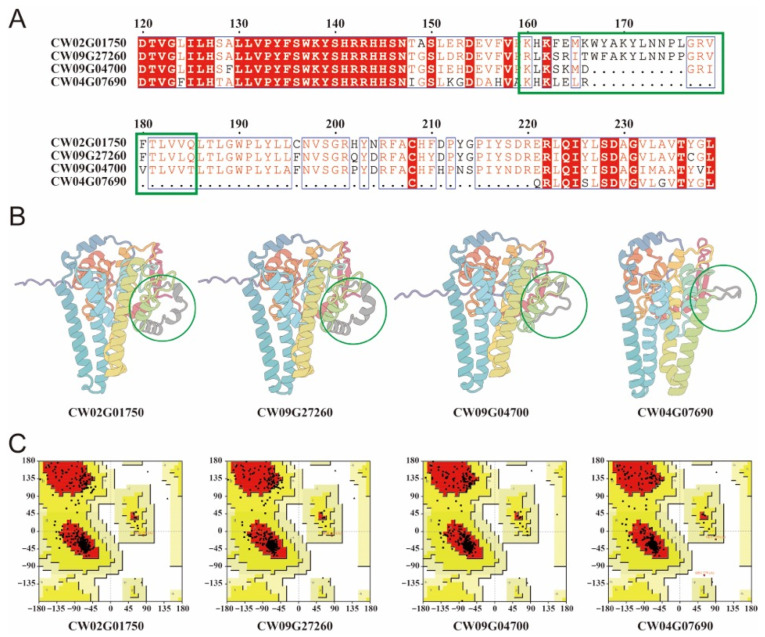 Figure 2
Figure 2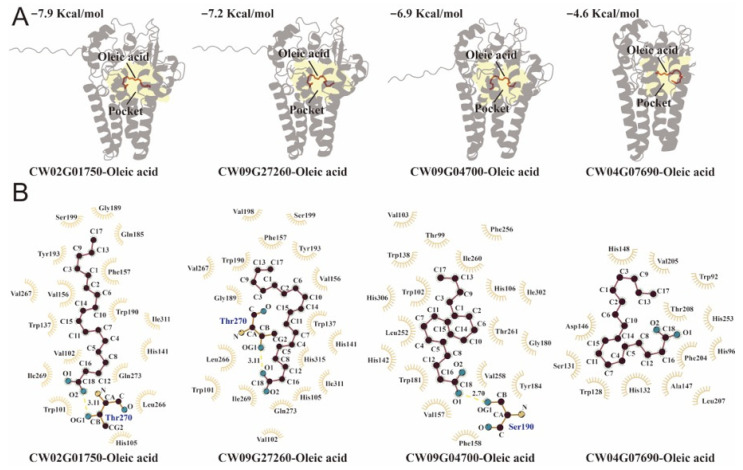 Figure 3
Figure 3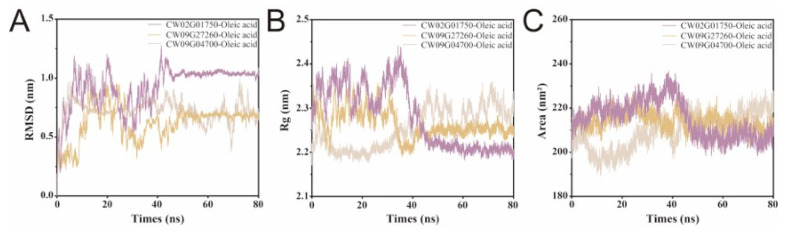 Figure 4
Figure 4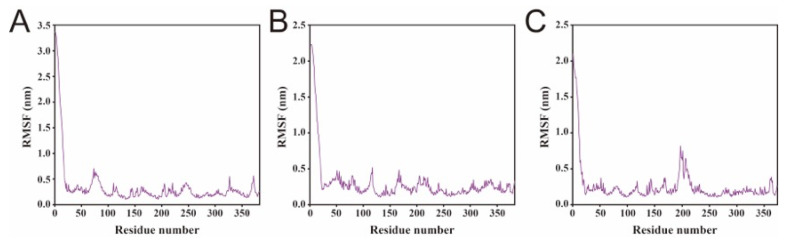 Figure 5
Figure 5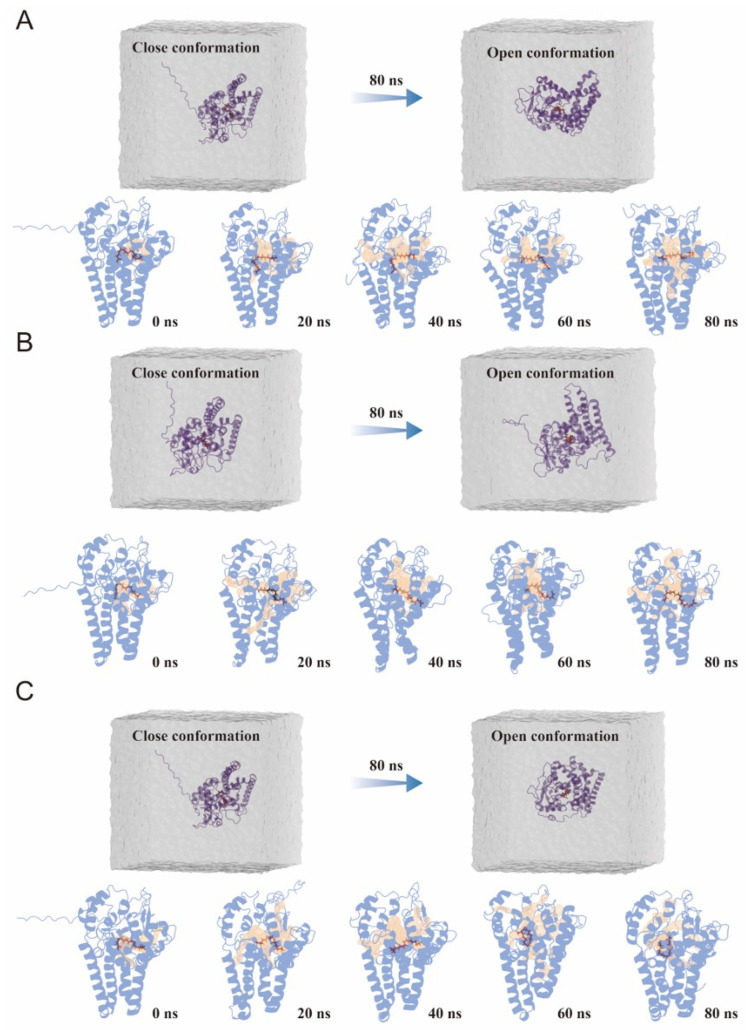 Figure 6
Figure 6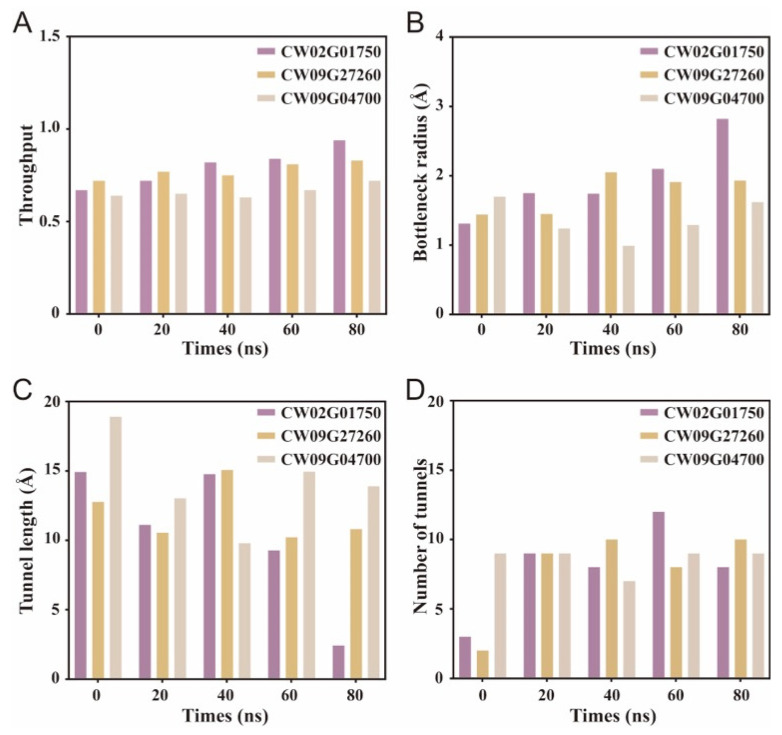 Figure 7
Figure 7- —Science and Technology Innovation Program of Hunan Province
- —National Natural Science Foundation of China
- —Central Government Guidance Funds for Local Science and Technology Development
- —Hunan Provincial Innovation Platforms & Talent Program, Major S&T Innovation Platforms
- —Changsha Science & Technology Program—‘Unveiling and Leading’ Major S&T Projects
- —Huxiang High-Level Talent Aggregation Initiative
- —Research Project of Changsha University of Science & Technology, Hunan Agricultural University—Changsha Innovation Institute of Modern Food
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLipid metabolism and biosynthesis · Food Chemistry and Fat Analysis · Edible Oils Quality and Analysis
1. Introduction
Cornus wilsoniana is an emerging woody oil crop with substantial ecological and economic value [1]. The fruit oil is characterized by a high proportion of C18 unsaturated fatty acids, particularly oleic acid (C18:1) and linoleic acid (C18:2), which together largely determine nutritional quality, oxidative stability, and the storage performance of plant oils [2,3,4]. Among these traits, the oleic-to-linoleic acid ratio (O/L ratio) is widely recognized as a pivotal indicator linking lipid composition to both health-related attributes and technological properties [5,6]. Therefore, elucidating the molecular basis governing the O/L ratio in C. wilsoniana is not only important for understanding lipid metabolism in woody oil species but also valuable for breeding and targeted improvement of oil quality.
Fatty acid desaturation constitutes the key biochemical route shaping the C18 fatty-acid profile [7]. In higher plants, the conversion of oleic acid to linoleic acid is predominantly catalyzed by microsomal ω-6 fatty acid desaturase (FAD2), which operates as a rate-limiting step that directly sets the O/L ratio [8,9,10]. In our previous chromosome-level genome study of C. wilsoniana, we systematically annotated genes related to fatty acid desaturation and identified four candidate FAD2 homologs (CW02G01750, CW09G27260, CW09G04700, and CW04G07690), providing a solid genomic foundation for functional dissection [11]. However, the relative contribution of these FAD2 homologs to oil formation—particularly during the rapid oil-accumulation stage—remains unclear, and the mechanistic basis underlying potential functional divergence among the four FAD2 proteins has not yet been resolved.
In addition to enzyme identity, the O/L ratio can also be influenced by the expression level and developmental timing of individual FAD2 homologs, substrate availability in cellular acyl-lipid pools, and the endoplasmic-reticulum membrane environment that constrains substrate access and tunnel geometry. Although these factors were outside the scope of the present work, we acknowledge their importance when interpreting simulation-based prioritization of candidate FAD2s.
Although multiple enzymes participate in the C18 desaturation network, not all steps contribute equally to variation in the O/L ratio of C. wilsoniana oil [12]. In contrast to the direct role of FAD2 in controlling the flux from C18:1 to C18:2, upstream and downstream steps are less informative for explaining O/L differences when their corresponding fatty-acid pools remain relatively low or stable in mature oil [13,14]. Consequently, focusing on FAD2 represents the most parsimonious and mechanistically grounded strategy to interpret and regulate the O/L ratio in C. wilsoniana, while simultaneously enabling a clear linkage from gene identity to biochemical outcome.
Accordingly, this study aims to prioritize candidate FAD2 homologs that may contribute to the oleate-to-linoleate conversion step relevant to the O/L ratio in C. wilsoniana oil by using a comparative structural and dynamic workflow. First, we quantified the fatty-acid composition of C. wilsoniana oil by gas chromatography, thereby defining the relative abundance of oleic and linoleic acids and the characteristic O/L ratio. Building on this phenotypic profile, we performed homology modeling and multiple sequence alignment of the four candidate FAD2 proteins to pinpoint divergent segments and putative key residues. We then conducted molecular docking to compare substrate-binding affinities among FAD2 homologs and examined differences in the geometry of the catalytic pocket to structurally assess substrate recognition and positioning. Finally, molecular dynamics simulations were employed to characterize the dynamic binding behavior and stability of the protein-substrate complexes, with additional analyses focusing on potential catalytic substrate-access tunnels. By integrating fatty-acid phenotype, sequence divergence, three-dimensional structure, and substrate binding dynamics, our work links FAD2 sequence variation with substrate accommodation and access-pathway behavior, thereby providing a structure-informed basis for prioritizing targets for future functional validation and oil-quality improvement.
2. Materials and Methods
2.1. Materials
Fruits of Cornus wilsoniana were collected from the National Tree Breeding Base at the Experimental Forest Farm of the Hunan Academy of Forestry (Changsha, China). The harvested samples were rapidly frozen in liquid nitrogen and stored at −80 °C prior to subsequent analyses.
Fatty-acid standards were obtained from Sigma-Aldrich (St. Louis, MO, USA). Solvents of chromatographic grade and other chemicals of analytical grade were used for fatty-acid methyl ester (FAME) preparation and GC analysis. Fatty-acid profiling was conducted using a Nexis GC-2030 gas chromatograph equipped with a flame ionization detector (Shimadzu, Kyoto, Japan); other routine laboratory instruments were used as needed.
2.2. Extraction and Methylation of Fatty Acids
Fatty acid methyl esters (FAMEs) were prepared from C. wilsoniana fruit oil using an alkaline methylation approach. Briefly, 0.06 g of oil was dissolved in 4.0 mL isooctane in a 10 mL centrifuge tube, followed by the addition of 2.0 mL KOH–methanol solution (2 mol/L). The mixture was vigorously shaken for 60 s to allow methylation.
To terminate the reaction and facilitate phase separation, 1.0 g anhydrous NaHSO_4_ was added, and the tube was shaken for another 30 s and left standing for 10 min. The upper organic layer was collected, passed through a 0.22 μm organic membrane, and used for GC analysis.
2.3. Gas Chromatography Analysis (GC–FID)
FAMEs were analyzed on a Nexis GC-2030 gas chromatograph (Shimadzu Corporation, Kyoto, Japan) equipped with a flame ionization detector and a fused-silica capillary column (0.25 mm × 100 m, 0.25 μm film thickness). Nitrogen served as the carrier gas at 1.1 mL/min, and samples were introduced in split mode (split ratio 1:100). The injector temperature was maintained at 270 °C, and 1.0 μL was injected for each run. GC–FID analyses were conducted using three independent biological replicates, each prepared from fruits collected from different individual trees.
The oven program was as follows: 100 °C for 13 min; ramp to 180 °C at 10 °C/min and hold for 6 min; ramp to 200 °C at 1 °C/min and hold for 20 min; then ramp to 230 °C at 4 °C/min and hold for 10.5 min. Fatty acids were identified by matching retention times against the Supelco 37 Component FAME Mix (Supelco, Bellefonte, PA, USA). Relative fatty-acid contents were calculated by normalizing individual peak areas to the total integrated peak area.
2.4. Homology Modeling and Multiple Sequence Alignment
Homology models of the four candidate FAD2 proteins were generated using the SWISS-MODEL server (https://swissmodel.expasy.org/) accessed on 25 November 2025. Model quality was primarily evaluated using the Global Model Quality Estimation (GMQE) score provided by SWISS-MODEL, and stereochemical reasonability was further assessed using Ramachandran plots generated via the SAVES v6.1 server (https://saves.mbi.ucla.edu/) accessed on 25 November 2025. Multiple sequence alignment of the four FAD2 proteins was performed, and the alignment was visualized using ESPript 3.2 (https://espript.ibcp.fr/ESPript/ESPript/) accessed on 26 November 2025 with emphasis on the divergent regions among the four sequences.
For homology modeling, SWISS-MODEL selected AlphaFold DB templates as follows: CW02G01750 and CW09G27260 were modeled using template I1N982.1.A (sequence identity 81.98% and 80.16%, respectively; coverage 100%); CW09G04700 used template Q8GZC3.1.A (sequence identity 79.89%; coverage 100%); CW04G07690 used template A0A5J4ZHK7.1.A (sequence identity 80.31%; coverage 99.7%).
2.5. Molecular Docking
Molecular docking was performed to compare the binding affinity of the four modeled FAD2 proteins toward the representative ligand (oleic acid, C18:1) and to characterize the catalytic pocket involved in substrate binding. The 3D structure of the ligand was obtained from the PubChem database and prepared prior to docking.
Docking calculations were carried out using AutoDock Vina (v1.1.2) [15]. Receptor and ligand structures were processed with AutoDockTools (e.g., hydrogen addition, Gasteiger charge assignment, and conversion to PDBQT format). A cubic grid box (50 × 50 × 50 Å) was applied to cover the catalytic region, and the best-scoring pose (lowest binding energy) was retained for subsequent pocket comparison and structural interpretation. Docking poses were inspected and visualized using VMD (v1.9.2). Protein–ligand interaction diagrams were generated using LigPlot+ (v2.2.9) to summarize key contacts within the catalytic pocket [16].
The docking search space was defined by cavity prediction on each modeled FAD2 structure. The grid box was centered on the centroid of the predicted substrate-binding cavity and sized to fully encompass the catalytic pocket and accommodate oleic acid.
2.6. Molecular Dynamics Simulation and Catalytic Tunnel Prediction
Molecular dynamics (MD) simulations were carried out with GROMACS 2020.6 [17]. The CHARMM36 force field was used for the FAD2 proteins, and ligand topology/parameters were generated using the CGenFF 3.0.1 webserver (https://cgenff.com) accessed on 7 December 2025. CGenFF penalty scores were inspected to assess parameter transferability; all penalties were below 10, with a maximum penalty score of 3.083, indicating a fair analogy for oleic acid. The CGenFF stream file is provided in the Supplementary Materials. Each FAD2–ligand complex was solvated in an orthorhombic box with TIP3P water, followed by the addition of Na^+^/Cl^−^ ions to neutralize the system. Energy minimization was performed using the steepest-descent algorithm (5000 steps), and the system was subsequently equilibrated under NVT and NPT ensembles (100 ps each) at 300 K and 1 atm. Production simulations were run for 80 ns with a 2 fs time step. For each complex, one 80 ns trajectory was generated; therefore, the MD-based comparisons are intended to be qualitative and trend-focused. Convergence was assessed by monitoring the stabilization of RMSD, Rg, and SASA profiles. The LINCS algorithm was applied to constrain bond lengths, and long-range electrostatics were treated using the particle-mesh Ewald (PME) method.
Trajectory convergence was evaluated by monitoring three global descriptors: backbone RMSD (nm), radius of gyration (Rg, nm), and solvent-accessible surface area (SASA, nm^2^). A quasi-equilibrated regime was defined as a continuous time window showing no systematic drift in these descriptors and only fluctuations around stable mean values.
Catalytic tunnel prediction was performed using the CAVER 3.0.3 plugin, enabling comparative analysis of putative substrate-access channels among different FAD2 variants. To improve transparency and reproducibility, identical CAVER settings were applied to all snapshots: minimum probe radius 0.9 Å, shell depth 4 Å, shell radius 3 Å, clustering threshold 3.5 Å, and 12 approximating balls. The starting point was defined as the centroid of a predefined atom selection near the catalytic pocket center; for each snapshot, the centroid coordinates were recalculated using the same selection criteria. Starting-point optimization used a maximum distance of 3 Å and a desired radius of 5 Å. Tunnels were ranked by throughput, and the dominant tunnel was defined as the path with the highest throughput in each snapshot.
3. Results
3.1. Fatty-Acid Composition of Cornus Wilsoniana Oil
Gas chromatography profiling revealed a simple fatty-acid pattern in Cornus wilsoniana oil, with several dominant peaks corresponding to major components (Figure 1A). The chromatogram showed clearly resolved signals for linoleic acid, oleic acid, and palmitic acid, with isooctane appearing as the solvent peak.
Quantitative analysis indicated that linoleic acid was the most abundant fatty acid (46.03%), followed by oleic acid (30.25%) and palmitic acid (17.43%), whereas the remaining fatty acids collectively accounted for less than 7% of the total (Figure 1B). Notably, linoleic acid and oleic acid were present at significantly higher levels than palmitic acid and the minor components, highlighting a C18 unsaturated fatty-acid-enriched profile in C. wilsoniana oil.
3.2. Sequence Divergence and Structural Differences Among Four FAD2 Homologs
Multiple sequence alignment of the four C. wilsoniana FAD2 homologs revealed that sequence divergence was mainly confined to a localized segment, with the most prominent variations occurring within the region spanning approximately residues 160–185 (Figure 2A). Within this divergent segment, CW09G04700 exhibited a small deletion, whereas CW04G07690 contained an extensive deletion, suggesting that the integrity of this region may differ substantially among the four homologs and potentially contribute to functional variation. The amino-acid sequences of the four FAD2 proteins are provided in the Supplementary Materials.
Consistent with the alignment results, homology models showed that CW02G01750 and CW09G27260 adopted highly similar overall conformations, while structural deviations were primarily observed at the divergent region highlighted in Figure 2B. In CW09G04700, the corresponding α-helical element in this region appeared to transition into a more flexible coil, whereas CW04G07690 displayed partial loss of secondary-structure elements, in agreement with the larger deletion detected by sequence alignment. Model quality assessment using Ramachandran plots indicated that all four models had more than 90% of residues located in favored or allowed regions (Figure 2C), supporting the reliability of the modeled structures for subsequent comparative analyses [18].
3.3. Molecular Docking Reveals Differential Binding of Oleic Acid to Four FAD2 Homologs
Docking analysis showed that oleic acid could be accommodated within the catalytic pocket of each modeled FAD2 protein (Figure 3A). The predicted binding affinities were −7.9 kcal/mol for CW02G01750, −7.2 kcal/mol for CW09G27260, −6.9 kcal/mol for CW09G04700, and −4.6 kcal/mol for CW04G07690. These results indicate the strongest interaction for CW02G01750 and the weakest for CW04G07690. In CW04G07690, the pocket region appeared markedly reduced in volume, consistent with its extensive deletion in the divergent segment, suggesting that the catalytic environment required for stable substrate binding may be compromised. Here, −5 kcal/mol was treated solely as a practical, AutoDock Vina scoring-specific reference to flag low-ranking poses within our dataset; it is not a universal biophysical threshold for binding, and its interpretation is protein- and protocol-dependent [19]. Accordingly, together with the markedly reduced pocket volume and the absence of stabilizing hydrogen bonds, CW04G07690 was deprioritized and not carried forward to downstream MD analyses.
The 2D interaction maps further supported these differences (Figure 3B). In both CW02G01750 and CW09G27260, Thr270 formed a hydrogen bond with oleic acid with a bond length of 3.11 Å. In CW09G04700, Ser190 formed a hydrogen bond with oleic acid with a bond length of 2.70 Å. In contrast, no hydrogen bond was detected between CW04G07690 and oleic acid, consistent with its weak docking affinity.
3.4. Molecular Dynamics Simulations Reveal Distinct Stability and Flexibility Among FAD2–Oleic Acid Complexes
Molecular dynamics simulations were conducted to evaluate the conformational stability and dynamic behavior of the three FAD2–oleic acid complexes retained after docking. Because one trajectory was analyzed for each complex, the results below are reported as trajectory-level comparisons that emphasize consistent trends across multiple observables. Time-dependent RMSD trajectories showed that CW02G01750 and CW09G27260 approached a relatively stable regime at around 50 ns, with CW09G27260 maintaining a lower post-equilibration RMSD than CW02G01750, indicating smaller overall structural deviation during the late stage of the simulation (Figure 4A). In contrast, CW09G04700 exhibited persistent mild fluctuations beyond 50 ns, suggesting increased conformational flexibility [20]. In the quasi-equilibrated segment (50–80 ns), CW02G01750 and CW09G27260 fluctuated within approximately 1.0–1.1 nm and 0.6–0.7 nm RMSD, respectively, whereas CW09G04700 remained more variable at about 0.6–0.9 nm. Consistently, Rg stabilized at approximately 2.20–2.23 nm for CW02G01750, 2.23–2.26 nm for CW09G27260, and 2.27–2.33 nm for CW09G04700, with corresponding SASA plateaus of approximately 205–212, 212–220, and 215–230 nm^2^.
Consistent with RMSD, the radius of gyration and solvent-accessible surface area converged for all three complexes at approximately 45 ns, while clear differences remained in their equilibrated levels (Figure 4B,C). CW02G01750 displayed a lower Rg than CW09G27260 after equilibration, indicating a comparatively more compact conformation, whereas CW09G04700 showed the highest Rg and continued to fluctuate slightly in the later phase, accompanied by a similar trend in surface area [21,22]. Residue-level RMSF profiles further highlighted distinct flexibility patterns among the three proteins (Figure 5) [23,24]. CW02G01750 and CW09G27260 exhibited generally low fluctuations across most residues, with higher mobility mainly confined to the N-terminal region. In contrast, CW09G04700 showed elevated RMSF in an internal segment centered around residues 180–220, consistent with the major divergent region identified by sequence/structure comparison. Notably, increased local flexibility within the protein core is more likely to affect functional architecture than terminal motions, as it may perturb the geometry and stability of the catalytic pocket and its surrounding access pathway [25,26,27]. Together, these results indicate that CW09G04700 samples a more dynamic conformational ensemble, whereas CW02G01750 and CW09G27260 maintain comparatively higher structural stability during the simulation.
3.5. Conformational Transitions and Catalytic Tunnel Dynamics During MD Simulations
To visualize how substrate binding is coupled to structural rearrangements, we tracked the conformational evolution of the three FAD2–oleic acid complexes and the corresponding catalytic tunnels predicted along the MD trajectories (Figure 6). Based on the 80 ns end-point structures, the divergent segment spanning residues 160–185, highlighted in red in Figure S1, is positioned at the rim of the catalytic pocket and adjacent to the entrance of the dominant access tunnel, providing a residue-level structural context for the access-pathway behaviors observed during the trajectories. At the system level, the water-box views indicate a clear transition from a relatively closed state to a more open conformation over the 80 ns simulation, accompanied by increased exposure of the catalytic pocket and stabilization of oleic acid within the active-site region.
Snapshots extracted every 20 ns further highlight distinct dynamic patterns among the three homologs. For CW02G01750 and CW09G27260, oleic acid gradually adopted a more extended conformation and became progressively accommodated in the catalytic pocket as the simulation proceeded. In parallel, the predicted tunnel network expanded and remained more pronounced toward the later stage of the trajectories, consistent with a structural shift toward an open state that facilitates ligand access and positioning. In contrast, CW09G04700 displayed a less consistent binding trajectory, in which oleic acid underwent a sequence of conformational rearrangements from a folded state to a more extended state and then returned to a more compact conformation at the end of the simulation. This behavior coincided with a tunnel pattern that increased during the middle stage but decreased in the late stage, suggesting that transient narrowing of the access pathway may impose additional constraints on ligand accommodation and contribute to a less stable binding mode [28]. These qualitative patterns are further supported by the snapshot-based tunnel descriptors summarized in Figure 7.
To enable objective comparisons, we extracted the CAVER output for each sampled structure and summarized the dominant-tunnel throughput, bottleneck radius, and tunnel length, together with the total number of detected tunnels (Figure 7A–D). Across the sampled snapshots, CW02G01750 exhibited an overall increase in throughput and a marked widening of the bottleneck radius toward the late stage, whereas CW09G27260 remained comparatively high and stable. In contrast, CW09G04700 showed lower throughput with a narrower bottleneck at mid-to-late snapshots, despite a relatively high number of detected tunnels, indicating a less permissive access architecture for sustained ligand accommodation. The dominant tunnel was defined as the highest-throughput tunnel identified by CAVER in each snapshot.
4. Discussion
This study provides a structure-informed framework to prioritize candidate FAD2 enzymes potentially underlying the oleic-to-linoleic conversion step associated with the O/L ratio in Cornus wilsoniana oil by focusing on FAD2, the microsomal ω-6 desaturase that directly catalyzes oleate-to-linoleate conversion. The fatty-acid profile showed that C18 unsaturated fatty acids dominated the oil composition, with linoleic and oleic acids constituting the principal components. Such a composition implies a strong flux through the C18:1-to-C18:2 conversion step, thereby supporting FAD2 as a logical and mechanistically proximal target for explaining and potentially manipulating the O/L ratio in C. wilsoniana.
A key finding is that sequence divergence among the four annotated FAD2 homologs is not diffuse across the entire protein but rather concentrates within a short segment around residues 160–185. As shown in Figure S1, this segment lies in close proximity to the tunnel mouth and pocket boundary, providing a structural rationale for how local sequence changes in this region can propagate to differences in access-pathway behavior observed during simulations. This region exhibited limited deletion in CW09G04700 and an extensive deletion in CW04G07690, and these differences translated into clear structural consequences in the corresponding homology models. The observation that CW04G07690 loses secondary-structure elements in the divergent segment suggests that this region contributes to maintaining the integrity of the catalytic architecture. Importantly, Ramachandran validation indicated that the models are of acceptable stereochemical quality, supporting their use as comparative templates for subsequent docking and dynamics analyses while acknowledging that membrane proteins remain challenging targets for high-accuracy modeling [29,30].
Docking results further suggest that the divergent segment can propagate to functional readouts at the level of substrate accommodation. The markedly weaker binding predicted for CW04G07690 was accompanied by a visibly reduced catalytic pocket, consistent with the hypothesis that large deletions disrupt the pocket environment required for stable oleate positioning. Excluding CW04G07690 from downstream simulations was therefore a pragmatic step to avoid over-interpreting a complex that already showed weak docking performance and lacked stabilizing hydrogen bonds. Among the remaining three homologs, CW02G01750 exhibited the strongest docking affinity, whereas CW09G04700 ranked lowest, a pattern consistent with their subsequent dynamic behaviors.
Molecular dynamics simulations provided a dynamic complement to static docking scores by capturing stability, compactness, residue flexibility, and access-pathway behavior over time [31,32]. CW02G01750 and CW09G27260 converged to relatively stable RMSD regimes and displayed lower, more uniform residue fluctuations across the protein core, with mobility largely confined to terminal regions. In contrast, CW09G04700 showed elevated RMSF within an internal region around residues 180–220 and sustained mild fluctuations in global metrics, which is consistent with the replacement of an α-helical element by a more flexible coil-like segment in the divergent region. Notably, increased local flexibility within the protein core is more likely to affect functional architecture than terminal motions, as it may perturb the geometry and stability of the catalytic pocket and its surrounding access pathway [33]. Consistently, tunnel analysis with quantitative descriptors showed that CW02G01750 and CW09G27260 tended to maintain higher throughput scores and wider bottleneck radii across the sampled snapshots, whereas CW09G04700 displayed greater variability in bottleneck size and access-route geometry, accompanied by repeated ligand readjustments. These results support the view that catalytic competence depends not only on initial binding but also on maintaining an accommodating access pathway that stabilizes substrate positioning during the catalytic cycle.
Importantly, the functional contribution of a given FAD2 homolog to the mature-oil O/L ratio in vivo is determined not only by structural competence inferred from simulations but also by expression level, developmental timing, ER localization, and regulatory context. Therefore, the present results should be interpreted as a structure- and dynamics-based prioritization of candidate FAD2 enzymes, rather than definitive evidence of physiological dominance.
A key limitation is that FAD2 is an endoplasmic-reticulum membrane protein, whereas our simulations were conducted in aqueous solvent without a lipid bilayer. In a native membrane context, lipid-mediated constraints may reshape the tunnel entrance and modulate transient opening events; thus, the tunnel geometries reported here should be viewed as comparative indicators under a simplified environment rather than quantitatively physiological descriptors. Another limitation is that the MD-based inferences were derived from single 80 ns trajectories for each complex; accordingly, the MD outcomes were interpreted as comparative, trend-level evidence supported by concordant patterns across multiple observables. Future work should include membrane-embedded simulations, replicate trajectories, and activity assays to test the proposed access-pathway mechanism and the predicted functional hierarchy.
From an applied perspective, the combined evidence supports CW02G01750 as a strong candidate for a major functional FAD2 in C. wilsoniana and suggests that CW09G27260 may serve as a secondary contributor with comparable structural robustness. CW09G04700 appears capable of binding oleic acid but may do so with reduced stability due to increased internal flexibility and a less persistent access pathway. CW04G07690, given its extensive deletion and weak docking performance, may represent a low-activity or conditionally active homolog, or could be a pseudogene-like variant, although functional validation would be required to distinguish these possibilities. Future work integrating expression profiling during oil accumulation, membrane-mimetic simulations, and enzymatic assays will be important to test the predicted functional hierarchy and refine the mechanistic link between access-pathway dynamics and oleate desaturation efficiency.
5. Conclusions
In this study, we profiled the fatty-acid composition of Cornus wilsoniana oil and compared four annotated FAD2 homologs to identify structural features potentially governing the oleic-to-linoleic conversion step relevant to the O/L ratio. GC-FID confirmed that C18 unsaturated fatty acids dominated the oil, supporting FAD2 as a proximal mechanistic target. A short variable segment accounted for most sequence divergence and produced distinct local structural effects. Docking indicated that CW04G07690 carries a compromised catalytic pocket and weak oleic-acid binding, and it was therefore excluded from downstream analyses. For the remaining homologs, 80 ns molecular dynamics simulations coupled with tunnel analysis revealed that CW02G01750 and CW09G27260 maintain greater conformational stability and increasingly accessible catalytic architectures, whereas CW09G04700 shows elevated internal flexibility and less persistent access pathways with repeated ligand conformational adjustments. Collectively, these findings rank CW02G01750 as a leading structural and dynamic candidate among the annotated FAD2 homologs in C. wilsoniana, with CW09G27260 likely contributing in a complementary manner. Importantly, the present study is limited by the lack of experimental validation at the expression and enzyme-activity levels; therefore, our conclusions represent a structure- and dynamics-based prioritization rather than definitive evidence of physiological dominance or in vivo flux control. Future work integrating expression profiling during oil accumulation and heterologous activity assays, ideally in membrane-mimetic contexts, will be necessary to validate the predicted functional hierarchy and establish a direct link between access-pathway dynamics and oleate desaturation efficiency. This work provides a structure-informed foundation for future efforts aimed at validating and improving the O/L ratio and oil quality in woody oil crops.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chen Y.Z. Gao Z.Z. Yang Y. Liu Q. Jiang L.J. Chen J.Z. Zhou X. Zhang L.H. Ji Y.N. Tu J. Revealing the Existence of Diverse Strategies for Phosphorus Solubilization and Acquisition in Plant-Growth Promoting Streptomyces misionensis Sw B 1Microorganisms 20251337810.3390/microorganisms 1302037840005744 PMC 11858620 · doi ↗ · pubmed ↗
- 2Liu R.T. Yi J. Li S.H. Xia J.L. Li M. Ding H.Y. Xu L.N. Yang X.H. Yao N. Design and Preparation Technology of Single/Dual-Cross-Linking Copolymers Based on Swida wilsoniana Oil ACS Omega 20216219152192410.1021/acsomega.1c 0219934497887 PMC 8412939 · doi ↗ · pubmed ↗
- 3Halimi R.A. Raymond C.A. Barkla B.J. Mayes S. King G.J. Development of Selection Indices for Improvement of Seed Yield and Lipid Composition in Bambara Groundnut (Vigna subterranea (L.) Verdc.)Foods 2022118610.3390/foods 11010086 PMC 875073035010212 · doi ↗ · pubmed ↗
- 4Sabbahi R. Azzaoui K. Rhazi L. Ayerdi-Gotor A. Aussenac T. Depeint F. Taleb M. Hammouti B. Factors Affecting the Quality of Canola Grains and Their Implications for Grain-Based Foods Foods 202312221910.3390/foods 1211221937297464 PMC 10252454 · doi ↗ · pubmed ↗
- 5Rashid M.T. Liu K.L. Han S.M. Jatoi M.A. The Effects of Thermal Treatment on Lipid Oxidation, Protein Changes, and Storage Stabilization of Rice Bran Foods 202211400110.3390/foods 1124400136553743 PMC 9778295 · doi ↗ · pubmed ↗
- 6Shen Y. Shen Y. Liu Y.H. Bai Y. Liang M. Zhang X.Y. Chen Z.D. Characterization and functional analysis of Ah GPAT 9 gene involved in lipid synthesis in peanut (Arachis hypogaea L.)Front. Plant Sci.202314114430610.3389/fpls.2023.114430636844041 PMC 9950565 · doi ↗ · pubmed ↗
- 7Qin J.S. Kurt E. Lbassi T. Sa L. Xie D.M. Biotechnological production of omega-3 fatty acids: Current status and future perspectives Front. Microbiol.202314128029610.3389/fmicb.2023.128029638029217 PMC 10662050 · doi ↗ · pubmed ↗
- 8Hajiahmadi Z. Abedi A. Wei H. Sun W.B. Ruan H.H. Qiang Z.G. Movahedi A. Identification, evolution, expression, and docking studies of fatty acid desaturase genes in wheat (Triticum aestivum L.)BMC Genom.20202177810.1186/s 12864-020-07199-1PMC 765369233167859 · doi ↗ · pubmed ↗