Targeting of Bacteria Using Amylase-Degradable, Copper-Loaded Starch Nanoparticles
Nathan A. Jones, Usha Kadiyala, Benjamin Serratos, J. Scott VanEpps, Joerg Lahann

TL;DR
This paper introduces a new type of nanoparticle that targets bacteria by responding to their environment and nearby microbes, offering a potential solution to antibiotic resistance.
Contribution
The novelty lies in using amylase-degradable, copper-loaded starch nanoparticles that target bacteria based on microbial interactions and environmental cues.
Findings
Copper-loaded starch nanoparticles showed tenfold higher antibacterial activity against Staphylococcus aureus compared to free copper nanoparticles.
In co-cultures with Bacillus subtilis, antibacterial activity increased by 44% due to enzymatic degradation of the nanoparticles.
Abstract
Background/Objectives: The treatment of bacterial infections is complicated by emerging antibiotic resistance. This paper identifies a novel approach with a nanoparticle that targets bacterial surface charge and is responsive to the nutrient environment (i.e., glucose) and presence of metabolically active bystander species (i.e., amylase secretion) within microbial communities. Methods: Thus, metabolically responsive composite nanoparticles (440 ± 58 nm) were fabricated via electrohydrodynamic jetting of a cationic starch polymer incorporating 5–7 nm copper nanoparticles (0.3 wt%). Starch was selected as the base polymer, as it is a common carbon source for amylase-producing bacterial communities, in particular under glucose-limited growth conditions. Results: The resulting positively charged particles effectively associated with Gram-positive Staphylococcus aureus, forming…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
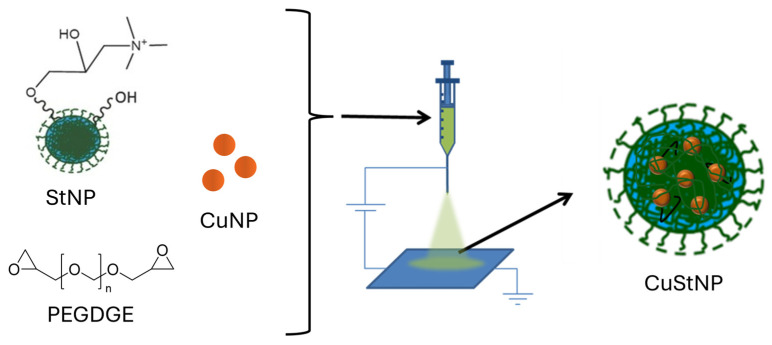 Figure 1
Figure 1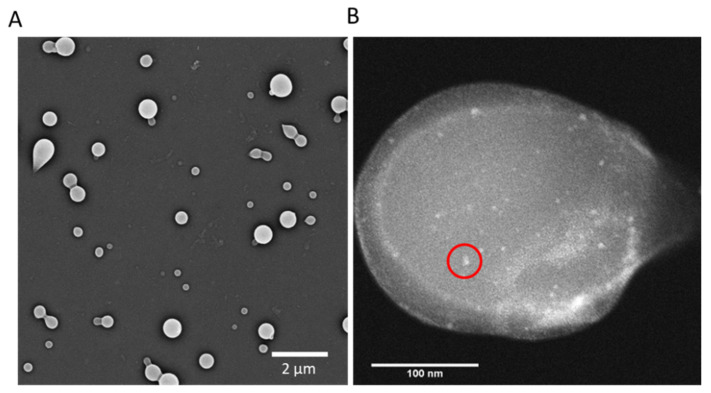 Figure 2
Figure 2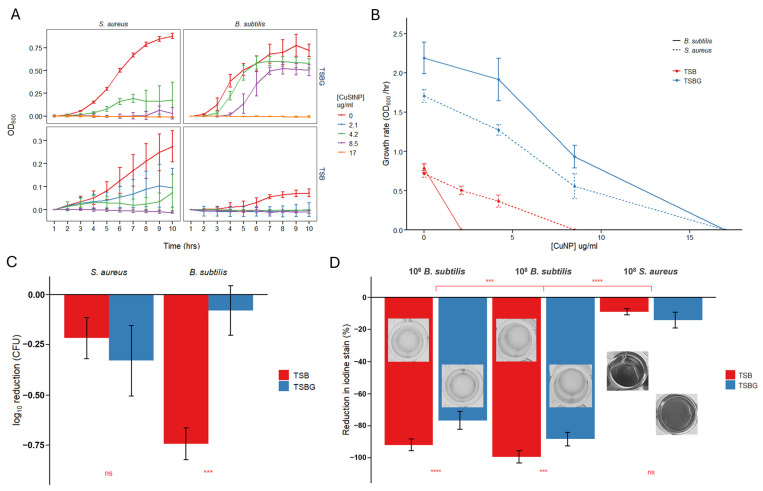 Figure 3
Figure 3 Figure 4
Figure 4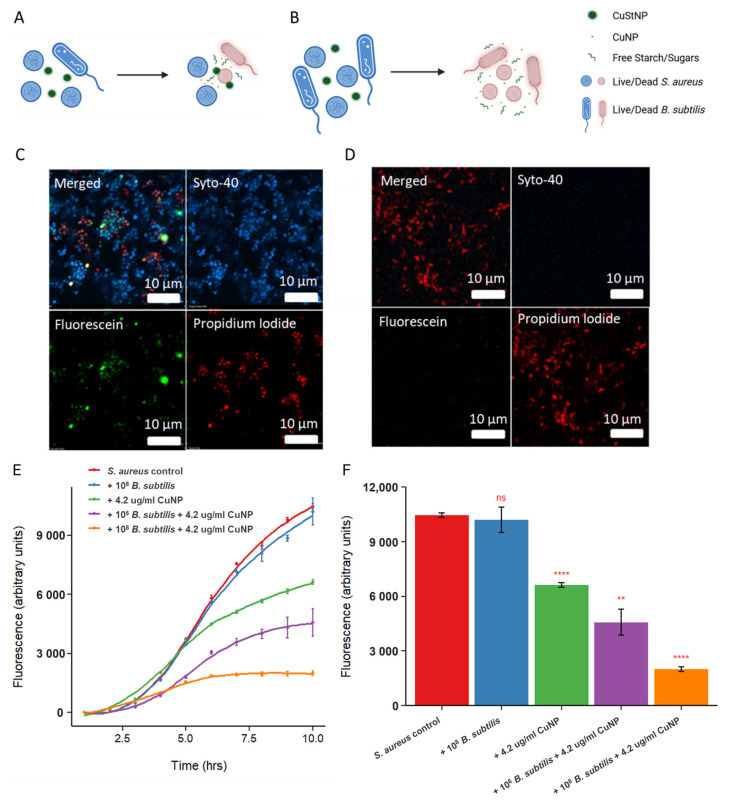 Figure 5
Figure 5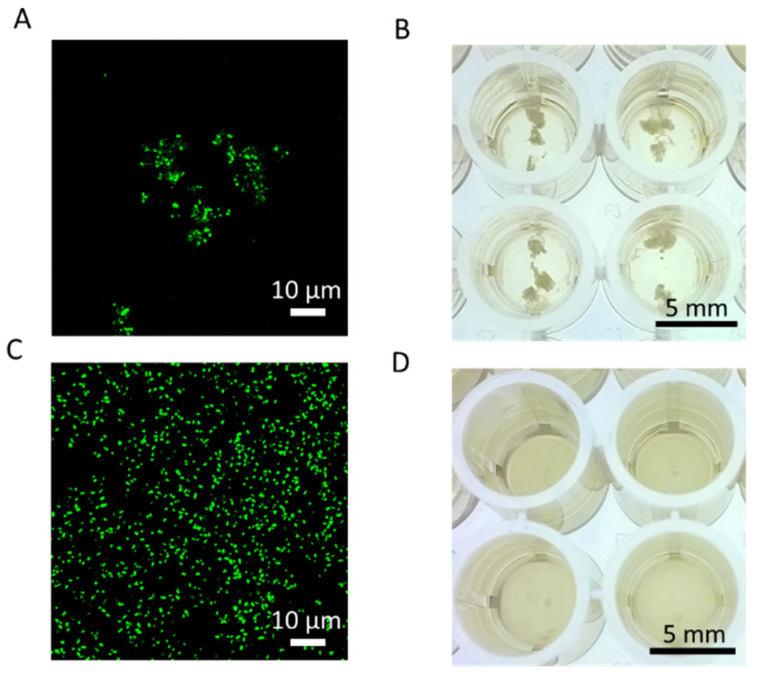 Figure 6
Figure 6- —National Institutes of Health (NIH)
- —National Institute for Allergy and Infectious Diseases (NIAID)
- —National Science Foundation (NSF)
- —Center for Complex Particle Systems (COMPASS)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMembrane Separation Technologies · Bacterial biofilms and quorum sensing · Nanoparticles: synthesis and applications
1. Introduction
Despite the widespread availability of small molecule antibiotics, bacterial infections remain a major cause of morbidity, mortality, and healthcare costs within hospital settings [1,2]. Approximately 3–13% of patients acquire an infection while being managed for a different condition, and incidence increases to nearly 50% for high-risk groups including newborns, immunocompromised patients, and those in intensive care units [3,4,5,6,7]. In the United States alone, antibiotic-resistant bacteria are involved in more than 2 million infections, with approximately 23,000 deaths annually [7].
Although the drivers for emerging antibiotic resistance are complex and multifactorial [8], they can be summarized as resulting from evolutionary pressure on bacteria outside the intended target or environment. Consequently, new options are needed to prevent and treat infections while limiting emerging resistance. To this end, recent efforts have explored antibacterial polymers [9] and metal nanoparticles, such as those made of silver [10,11,12], copper [12,13,14], or zinc oxide [15,16]. Metal nanoparticles exhibit broad-spectrum antibacterial activity with lower potential for resistance due to multiple simultaneous mechanisms of action, including oxidative stress, membrane disruption, metabolic disruption, and mismetallation [10,11,16].
However, the clinical translation of these antibacterial technologies is limited due to challenges such as poor aqueous solubility, scalability of production, and concerns over colloidal stability. Continued advances in medical nanotechnology are being developed to address these barriers, enabling improved targeted, responsive, and effective nanoparticle systems for bacterial infection control [17].
Targeted antibacterial approaches can be broadly categorized into passive and active schemes. Passive targeting methods rely on non-specific material properties to increase antibacterial dosage and efficacy. Bacterial infections, like cancerous tumors, are associated with increases in vascular permeability, allowing the Enhanced Permeation and Retention (EPR) effect for appropriately sized nanoparticles delivered intravenously [18]. Electrostatic interactions offer an effective method for targeting pathogens. Most bacteria exhibit a negative surface charge under physiological conditions, enabling cationic nanoparticles and polymers to bind through multivalent electrostatic interactions [9,13,19]. These passive targeting approaches provide broad utility, including the ability to target polymicrobial infections [19].
Active targeting methods, in contrast, employ specific ligands, such as vancomycin [20], lectins [21], antibodies [22], or aptamers [23], which are conjugated to antimicrobial agents or nanoparticle carriers to selectively recognize and bind to pathogenic bacteria [17,24]. A related approach, termed secondary targeting, involves directing nanoparticles to immune cells such as macrophages or other leukocytes, which are recruited to the infection site [25,26].
Beyond targeting specificity, nanoparticles have been engineered to respond to physicochemical cues within the infection microenvironment. Local acidity, for example, can trigger the degradation of acid-sensitive polymers, such as poly(lactic-co-glycolic acid) (PLGA) [19,27], poly(caprolactone) (PCL) [28], or chitosan [13]. Some polymers become cationic in acidic environments, enhancing bacterial binding [9,19]. More recently, enzyme-sensitive nanoparticles have been prepared that decompose in the presence of bacterial phosphatase or phospholipase [29].
Starch has attracted research interest as a biocompatible, renewable matrix in composite antibacterial materials, where it serves as both a stabilizer and a carrier for metal nanoparticles such as copper, enabling controlled release and enhanced contact with microbial cells [30,31,32,33]. In starch–copper nanocomposites, the polysaccharide framework helps disperse and protect small CuNPs, leading to strong bactericidal effects against both Gram-positive (e.g., Staphylococcus aureus, Bacillus subtilis) and Gram-negative (e.g., Escherichia coli) bacteria, often at low (microgram per milliliter) concentrations. These systems typically show concentration-dependent growth inhibition, with more potent activity against staphylococci and B. subtilis. Their efficacy is frequently attributed to copper ion release, ROS generation, and direct nanoparticle–cell membrane interactions, making starch–copper composites promising candidates for antimicrobial coatings, wound dressings, and biomedical devices. Villanueva et al. developed multi-use antibacterial composite gels with high loading (2.5–7.5% w/w) of silica-capped CuNPs (125–938 nm) embedded in a starch matrix [33]. Dinda et al. prepared starch–copper nanocomposites (~4 nm, unknown loading) that demonstrated antibacterial efficacy against S. aureus and Pseudomonas aeruginosa with minimum inhibitory concentrations (MICs) of 60 µg/mL and 75 µg/mL, respectively [30]. Hasanin et al. developed myco-synthesized starch–copper nanocomposites (200 nm) doped with 9 nm copper nanoparticles at 0.24% w/w, with highly negative surface charge (−150 mV), and more limited efficacy (125 µg/mL MIC) against Gram-negative bacteria like E. coli, with higher efficacy against Gram-positive bacteria including B. subtilis (7.81 µg/mL MIC) [31]. More recently, Hou et al. evaluated the impact of starch modifications, including acetylation, esterification, and amination on the morphology and qualitative antibacterial efficacy of prepared starch–copper composites against S. aureus and E. coli, finding that aminated starch improved antibacterial efficacy presumably due to incorporation of cationic charges [32]. Starch can be digested rapidly (20–30 min) by amylase enzymes, which can be present in biological environments, such as in the mouth and small intestines, or secreted by certain types of bacteria. Starch degradation by environmental amylase has been used for metabolically responsive payload release [33,34], and bacterial amylase secretion can be similarly leveraged [35,36].
In an attempt to develop a targeted nanomedicine, we decided to design, synthesize, and validate an amylase-responsive, cationic, starch-based nanoparticle platform (CuStNPs) that electrostatically associates with bacteria, and rapidly releases antibacterial copper nanoparticles (CuNPs) in the presence of amylase, an enzyme released by certain bacterial species (target or bystander) in ecological neighborhoods under specific nutrient conditions [37,38]. Potential species include Bacillus spp., Lactobacillus spp., Pseudomonas spp., Vibrio spp., and E. coli [39]. The composite CuStNPs allow for dispersion of hydrophobic CuNPs in aqueous media by providing colloidal stability within a cationic starch matrix, which may facilitate translation of CuNPs to clinical applications. This work introduces a distinct and novel paradigm of targeted antibacterial therapy that exploits vulnerabilities related to the metabolic ecology of microbial communities via secreted enzymes like amylase, which could be modified for alternative payloads or enzyme-substrate pairs.
2. Results & Discussion
2.1. Particle Preparation and Characterization
Electrohydrodynamic (EHD) jetting of particles and fibers has been extensively studied by our group [19,40,41,42,43,44], and others [45,46,47], employing various polymer compositions such as poly(lactic-co-glycolic) acid [42], acetal dextran [43,44], and stimuli-responsive polymers [40,41]. Starch is less commonly used in the fabrication of nanoparticles due to the inherent crystallinity of amylose, which promotes uncontrolled aggregation of prepared nanostructures [48]. However, successful approaches have been achieved through chemical modification and cross-linking of starch [49,50,51,52,53]. Starch is an interesting low-cost biocompatible material for metabolically responsive antibacterial applications because it is a common metabolite that can be degraded by amylase enzymes, which are produced by certain bacterial species [37,38,39].
In this work, we use research-grade starch nanoparticles (StNPs), which were further modified to introduce cationic functionality [34]. These cationic StNPs were cross-linked following EHD jetting. Incorporation of other components into the jetting solution allows their encapsulation within the starch matrix. Specifically, 5–7 nm CuNPs were dispersed into the jetting solution, forming a semi-stable Pickering-type emulsion upon vortexing. This dispersion remained stable throughout jetting and deposition onto metal collection plates, prior to thermal curing and final CuStNP collection (Figure 1).
Representative SEM images (Figure 2A) show the spherical morphology and uniform size distribution of the resulting composite CuStNPs, while TEM (Figure 2B) demonstrates the homogeneous distribution of CuNPs within the larger starch matrix. Dynamic light scattering measurements indicated an average hydrodynamic diameter of 440 ± 58 nm. Comparison with size analysis by ImageJ v.2.0.0 from SEM [54], which showed a dry particle size of 193 ± 91 nm, suggests a swell ratio of approximately 2.3 ± 0.5. Particle surface charge was confirmed as cationic with a zeta potential of +22.7 ± 1.3 mV. Swelling of the particles is indicative of their “softness”, which allows for highly effective multivalent binding.
The copper content of CuStNP samples was quantified by inductively coupled plasma optical emission spectroscopy (ICP-OES) using a calibration curve made with copper ICP standards (Supplementary Figure S1; R^2^ > 0.99). Samples prepared in tryptic soy broth (TSB) at concentrations of 1, 2, and 4 mg/mL yielded an average CuNP loading of 0.34 ± 0.05 wt% with a loading efficiency of 89.8 ± 7.5% (mean ± standard error, n = 3). This loading is comparable to the loading reported in the recent literature for similar systems [31].
2.2. Antibacterial Efficacy
Antibacterial activity was assessed against three Gram-positive species (S. aureus, Staphylococcus epidermidis, and B. subtilis) and two Gram-negative species (E. coli and Klebsiella pneumoniae). S. aureus and S. epidermidis were included as common human skin and wound pathogens frequently used in antibacterial screening. E. coli and K. pneumoniae were selected as clinically important Gram-negative species associated with gastrointestinal and opportunistic infections. B. subtilis, although not typically a human pathogen, was included due to its well-characterized physiology and its endogenous amylase production, which allowed us to evaluate the stability and antibacterial performance of the starch-based nanoparticles in the presence of amylolytic activity. Bacteria were exposed to serial dilutions of CuStNP dispersions corresponding to 2.1–34 µg/mL CuNP (0.5–8.0 mg/mL CuStNP). Control experiments using unloaded StNPs were performed with S. aureus and E. coli and showed minimal effects on bacterial growth, even at the highest tested concentrations (Supplementary Figure S2).
Consistent with prior reports on copper nanoparticle activity [12,14,30,31,33], CuStNPs exhibited limited efficacy against Gram-negative bacteria (Supplementary Figure S3A,B), which has been hypothesized due to cell well composition resulting in stronger resilience to copper nanoparticles [30]. In contrast, strong antibacterial activity was observed against Gram-positive bacteria even at effective copper doses of 4.2 µg/mL for S. aureus and 8.5 µg/mL for B. subtilis and S. epidermidis (Figure 3A,C and Supplementary Figure S3C). This is an order of magnitude lower effective dose than previously reported for free CuNPs and other copper–starch composite systems [12,14,30,31,33]. Further mechanistic evaluation of potential reasons for this improved efficacy is explored below (Section 2.3). Dose–response curves were generated to compare the dose–response of each species’ growth for different media (Figure 3B). These results highlight a more significant reduction in growth rate constant for amylase-releasing B. subtilis absent supplemental glucose, with null growth at just 2.1 µg/mL CuNP vs. a dosage of 8.5 µg/mL CuNP for S. aureus, and further demonstrate complete eradication of bacterial growth with supplemental glucose at CuNP doses of 17 µg/mL.
We hypothesized that CuStNPs would exhibit greater antibacterial activity against B. subtilis than S. aureus due to the ability of B. subtilis to secrete amylase and enzymatically digest starch [39], thereby accelerating release of CuNPs. However, in TSB with supplemental glucose (TSBG), this trend was not observed in either growth curve analysis (Figure 3A) or quantitative culture (Figure 3C). There was a more potent response for S. aureus compared to B. subtilis, particularly at the 4.2 µg/mL CuStNP dose. However, when tested in unsupplemented TSB, the opposite trend emerged; B. subtilis growth was more strongly inhibited than S. aureus. This may partly reflect the reduced baseline growth of B. subtilis under glucose-limited conditions.
Nevertheless, the glucose-dependent differences suggest that B. subtilis metabolism and growth are sensitive to nutrient availability. Specifically, in carbohydrate-limited media, starch-degrading bacteria are expected to upregulate amylase expression to digest starch into monomer sugars for energy metabolism. This is analogous to the regulation of the lac operon in many bacterial species [38]. In this specific case, glucose limitation results in upregulation of amylase, which promotes enhanced nanoparticle degradation and CuNP release. Consistent with this hypothesis, CuStNPs exhibited increased antibacterial efficacy against B. subtilis in TSB (glucose limited) relative to TSBG (Figure 3B). Although a plausible explanation for the data, we cannot rule out the physical and chemical effects of glucose in the media, including osmolality and oxidative potential, as other contributors to this observation.
To complement the turbidometric growth curve analysis, quantitative culture studies were performed to directly measure bacterial killing (Figure 3C, Supplemental Figure S4). These results confirmed that B. subtilis killing is significantly greater (p = 0.00036) in unsupplemented TSB, whereas S. aureus showed no media-dependent difference (p = 0.213). At 4.2 µg/mL CuStNP, viable bacterial counts showed a maximum 1-log reduction with the following rank order of efficacy: B. subtilis in TSB > S. aureus in TSBG ≥ S. aureus in TSB > B. subtilis in TSBG (p < 0.001 by ANOVA with post hoc Tukey’s test, except between S. aureus media conditions).
Iodine staining confirmed starch degradation by B. subtilis but not S. aureus in both TSB and TSBG media following 8 h of incubation (Figure 3D). Starch degradation was significantly greater for B. subtilis in unsupplemented media (p = 0.000018 and p < 0.0001 for 10^8^ and 10^6^ cell counts, respectively), though more than 80% degradation occurred even in TSBG. At this late time point, apparent differences in degradation rate are not evident as increased cell density in TSBG cultures likely compensated for lower per-cell amylase expression.
2.3. Strong Interactions Between CuStNPs and S. aureus
Given the unexpectedly high antibacterial efficacy observed with CuStNPs relative to their copper content, we hypothesized that the nanoparticles may interact directly with bacterial cells, facilitating localized copper delivery and increasing the effective local dose. Therefore, to test this, S. aureus cultures were incubated with 4.2 µg/mL CuStNP for 8 h, then prepared for imaging by SEM, confocal microscopy, and digital imaging (Figure 4).
Because these CuStNPs comprise hydrophilic polymers, they display nanogel-like properties and swell to more than twice their size after suspension in water. We propose that the cationic charge of the particles promotes strong multivalent interactions with the negatively charged bacteria, leading to the formation of agglomerates comprising bacteria and starch hydrogel nanoparticles. In SEM images, the dehydrated particles appear as a smooth coating enveloping the bacterial cells (Figure 4A,B), consistent with the adhesion hypothesis.
This association likely accounts for the enhanced antibacterial activity of the CuNPs. Although the overall copper dose (~4.2 µg/mL) is approximately an order of magnitude lower than in comparable studies [12,14], the CuNPs are effectively concentrated with the bacteria by the cationic hydrogel carrier. Elemental mapping by EDX confirmed the presence of copper localized to bacterial aggregates (Figure 4C). These findings support a mechanism where electrostatic association between CuStNPs and bacterial cells increases the effective local CuNP concentration, resulting in significantly enhanced antimicrobial potency relative to free CuNP dispersions.
2.4. Co-Culture of Amylase and Non-Amylase Producing Species
Bacteria-mediated starch digestion, via amylase production, provides a way to confirm our hypotheses of the amylase-responsive antibacterial effect (Figure 3) and starch mediated aggregation with S. aureus (Figure 4). Unlike S. aureus, B. subtilis enzymatically digests starch through the synthesis and excretion of amylase. Bacterial co-cultures containing 10^8^ S. aureus and either 10^6^ or 10^8^ B. subtilis were exposed to CuStNPs (4.2 µg/mL CuNPs). It was hypothesized that increasing numbers of B. subtilis would increase the availability of the CuNPs, resulting in higher overall antibacterial activity. Confocal microscopy with Live/Dead™ staining of bacterial cells was used to observe and determine antibacterial activity trends for various co-culture conditions. Expected results based on this hypothesis are illustrated in Figure 5A,B.
Experimentally, after 8 h of incubation, the co-culture seeded with low relative count (10^6^) B. subtilis showed a mixture of live and dead cells, with green fluorescently labeled CuStNPs interspersed (Figure 5C). In contrast, the co-culture seeded with 10^8^ B. subtilis was completely dead with green fluorescence dispersed, likely due to the degradation of the particles (Figure 5D). To support these observations, growth curves were charted for a GFP-producing S. aureus strain as a function of various co-culture conditions (Figure 5E), which indicate that the presence of B. subtilis had minimal effect on the growth of S. aureus, and increasing the amount of starch-digesting B. subtilis synergistically improved the antibacterial efficacy of the CuStNPs against S. aureus. Quantitatively, at 10 h, the number of S. aureus cells was reduced by 37% by treatment with 4.2 µg/mL CuNPs (p < 0.0001). The addition of an equivalent number (10^6^) of amylase-producing bystander B. subtilis lead to an additional 19% or total 56% reduction (p = 0.0037), while the addition of 10^8^ B. subtilis cells had an additional 44% or total 81% reduction (p < 0.0001).
As noted, the cationic starch facilitated aggregation with negatively charged S. aureus. In co-culture samples with equivalent numbers of S. aureus and B. subtilis, this aggregation was observed microscopically using fluorescently labeled CuStNPs (Figure 6A), and macroscopically in the well plate (Figure 6B). In contrast, the co-culture sample with 100-fold excess of starch-digesting B. subtilis showed no evidence of aggregation (Figure 6C,D), presumably because the starch was digested by amylase.
2.5. Study Limitations and Future Directions
Our study demonstrates strong antibacterial activity of electrohydrodynamic jetted starch–copper nanocomposites against Gram-positive bacteria, but several limitations should be acknowledged. The current work focuses on planktonic growth inhibition in standard and glucose-free media, and future studies should extend to biofilm eradication, long-term stability of the composite under storage, and potential copper leaching under physiological conditions, as excessive copper ion release can raise biocompatibility concerns in biomedical applications [31]. Mechanical and barrier properties of the starch material (e.g., moisture sensitivity, mechanical strength) were not fully characterized here; evaluating these in future work would better inform the suitability of this material for wound dressings or food-contact surfaces. In addition, while efficacy against S. aureus, S. epidermidis, and B. subtilis is promising, broader testing against multidrug-resistant clinical isolates and in vivo models with amylase secretion (e.g., skin infection or wound healing models) will be essential to assess translational potential and safety profile.
3. Conclusions
In this study, we report the development of a novel composite starch nanoparticle platform prepared by reactive EHD jetting. The resulting particles, loaded with CuNPs, exhibited potent antibacterial activity against Gram-positive bacteria, including S. aureus, S. epidermidis, and B. subtilis. Antibacterial efficacy was improved under conditions favoring starch degradation, specifically in the presence of starch-digesting microbial communities (via amylase) and in media lacking glucose supplementation, demonstrating that nanoparticle composition can be leveraged to target therapeutic activity to specific metabolic environments.
The cationic charge of high-swelling CuStNPs facilitates strong electrostatic association with bacterial cells, localizing CuNPs at the bacterial surface and thereby increasing their effective potency. Co-culture experiments further revealed the potential for synergistic antibacterial effects in mixed bacterial populations containing starch-digesting species, as is relevant to natural microbiomes such as in the lower digestive tract.
Future work will investigate detailed mechanisms underlying antibacterial activity and evaluate the incorporation of other therapeutics into the StNP platform, potentially enabling targeted and metabolically regulated drug delivery within the gastrointestinal environment, wound dressings, or medical contact surfaces.
4. Materials and Methods
4.1. Materials
All chemicals were reagent-grade and purchased from Sigma Aldrich (St. Louis, MO, USA) except as noted. These include tryptic soy broth (TSB), glucose, glycidyltrimethylammonium chloride (GTAC), poly(ethylene glycol) diglycidyl ether (MW = 5k) (PEGDGE), sodium hydroxide, 2-propanol, and ethanol. Copper nanoparticles dispersed in organic phosphate ester, sized 5–7 nm (Product # 0851HN), were purchased from Sky Spring Nanomaterials (Houston, TX, USA). Research-grade starch nanoparticles (StNP) were provided by EcoSynthetix Inc. (Burlington, ON, Canada) [34].
4.2. Bacterial Strains, Media, & Growth Conditions
The bacterial strains used in this study were methicillin-resistant S. aureus COL [55], a green fluorescing protein (GFP) producing S. aureus strain (SH1000/pCM29) [56], B. subtilis PC194 [57], S. epidermidis 1457 [58], E. coli UTI89 [59], and K. pneumoniae LM21 [60]. Glycerol stocks of all strains maintained at −80 °C were plated on tryptic soy agar, cultured overnight at 37 °C and stored at 4 °C. Single colony inoculates were grown in tryptic soy broth with or without 1% glucose w/v (TSBG or TSB, respectively) under shaking conditions until optical density at 600 nm (OD_600_) reached 0.4–0.6 (mid-log), prior to dilution to OD_600_ of 0.01 for planktonic growth curves and quantitative culture studies.
4.3. Preparation & Characterization of Copper–Starch Nanoparticles
4.3.1. Electrohydrodynamic Jetting of Copper Starch Nanoparticles
Electrohydrodynamic (EHD) jetting was performed using StNPs to control particle size, morphology, drug loading, and incorporate additional functionalities. Initial tests using StNP dispersions in water were unsuccessful as the solution would drip; substitution of up to 20% ethanol (v/v) lowered solution surface tension, improving jetting stability and yielding nanoparticles.
Copper–starch composite nanoparticles (CuStNPs) were prepared using EHD jetting as illustrated by the schematic in Figure 1. Cationic StNPs were prepared by chemically modifying research-grade StNPs, as reported previously [34]. The jetting formulation contained 10% (m/v) cationic StNP, 3% (m/v) PEGDGE cross-linker, and ~0.5% (v/v) of a 5–7 nm CuNP solution, dispersed in a solution of 80:20 deionized water/ethanol. Samples were vortexed to disperse hydrophobic CuNPs, creating a Pickering emulsion that remained visually stable over 24 h. EHD jetting was performed immediately after preparation of the jetting solution, using a voltage of ~12 kV, a pump speed of 0.05 mL/hr, and a jetting needle to collection plate distance of ~30 cm. Collected plates were subsequently cured in a 50 °C oven for 72 h and prepared CuStNPs were collected as a dry aggregate powder. Fluorescent CuStNPs were prepared by the same protocol, with substitution of 10% (w/w) of the cationic StNPs with particles additionally modified with covalently linked fluorescein (i.e., 1% m/v fluorescent cationic StNPs, 9% cationic StNPs m/v in the jetting formulation) [34]. To our knowledge, EHD jetting has found limited application in the preparation of starch particles with research focus on fibers and films [61], and just one recent example that evaluated methods for preparation of crystalline starch particles [48].
4.3.2. Particle Size and Charge Characterization
Nanoparticle samples were dispersed in 0.01 M phosphate-buffered saline (PBS) solution at a concentration of 2.5 × 10^−4^ g/mL and characterized by dynamic light scattering (DLS) and electrophoretic light scattering (ELS) for zeta potential determination using a Malvern ZetaSizer (Malvern Panalytical, Malvern, UK). DLS measurements were performed at 25 °C with using automatic attenuation and measurement position selection. Zeta potential was determined by measuring electrophoretic mobility in folded capillary cells (Malvern Panalytical, Malvern, UK) via the Smoluchowski approximation. All samples were adjusted to pH 7.4 and measured in triplicate, allowing for calculation of average particle size and charge and estimates of error [62]. Particle size was also quantified from five representative scanning electron micrographs, each containing 50–100 particles per image, using the ImageJ v.2.0.0 particle analysis tool. To account for measurement variability, images were processed using three different sensitivity thresholds, and the resulting datasets were combined. All measurements were manually reviewed to correct for instances in which particle aggregates appeared, ensuring that only individual particles were included in the final size distribution.
4.3.3. Electron Microscopy
Scanning electron microscopy (SEM) imaging was performed using a Quanta SEM (FEI, Hillsboro, OR, USA) with beam voltage of 3 kV and a magnification of 10,000× and a NOVA SEM (FEI, Hillsboro, OR, USA) with beam voltage of 5 kV, 0.4 nA, and at a magnification of 10,000×. Energy-Dispersive X-ray Spectroscopy (EDX) analysis was conducted to determine atomic mapping of copper to correspond with captured SEM images. Samples were mounted onto aluminum Scanning Electron Microscopy (SEM) stubs using carbon adhesive tabs and allowed to dry completely. The samples were then sputter-coated with a ~10–20 nm gold layer using an SPI-Module Carbon/Sputter Coater (West Chester, PA, USA) operated under argon at 18 mA for 120 s. Gold coating was used to improve sample conductivity and minimize charging during electron beam exposure. Transmission electron microscopy (TEM) imaging was performed using a JEOL 2100 (JEOL, Tokyo, Japan) probe-corrected analytical microscope with an accelerating voltage of 100 kV, a zirconated tungsten (100) thermal field emission tip filament, with gun vacuum at ~10^−9^ torr and column vacuum at ~10^−7^ torr (Michigan Center for Materials Characterization). For these images, nanoparticle powder was applied directly onto carbon-coated copper TEM grids by gently tapping a small amount of the dry powder over the grid surface. Excess material was removed by tapping the grid before loading into the microscope.
4.3.4. Inductively Coupled Plasma—Orbital Emission Spectroscopy (ICP-OES) Evaluation of Copper Loading
ICP-OES measurements were performed using a Perkin-Elmer Optima 2000 DV (Waltham, MA, USA) with Winlab 6.4 software (MA, USA). Samples were doped with 1 ppm yttrium as a standard prior to degradation by addition of 5% (w/w) nitric acid. All CuStNP samples were prepared at mass concentration of 2 mg/mL in TSB and calibrated against a copper ICP standard (Sigma Aldrich, St. Louis, MO, USA). Three independent replicates were measured for each sample.
4.4. Bacterial Culture Studies
4.4.1. Bacterial Growth Curves
CuStNP suspensions were prepared by vortexing 4.0 mg/mL CuStNPs into TSB or TSBG for 5 min. The final CuStNP concentrations were serial dilutions of the original stock solution. To compare to previously reported data on copper nanoparticles, concentrations are shown in terms of the CuNP concentration, as determined by the ICP loading study. The maximum concentration used in this study (25 µg/mL CuNPs, equivalent to ~4.0 mg/mL CuStNP, based on calculated loading of approximately 0.37% (w/w) CuNPs) was comparable to previously reported CuNP data [12,14]. Bacterial growth was assessed by measuring OD_600_ (Perkin Elmer Enspire multimode plate reader, Waltham, MA, USA) hourly over 10 h in the presence of CuStNPs. Dose–response curves were generated from the averages of three independent replicates. To compare individual growth curves we calculated the growth rate constant as the slope of the initial linear portion (i.e., exponential phase) of the log_2_(OD_600_) vs. time data determined by linear regression [15]. For co-culture experiments, S. aureus SH1000, transformed with a GFP-producing plasmid (pCM29), was used with B. subtilis. S. aureus growth was independently assessed by tracking green fluorescence (excitation 480 nm/emission 520 nm). Error estimates were calculated as the standard deviation from three independent replicates.
4.4.2. Quantitative Culture
Bacterial cultures were exposed to 4.2 µg/mL CuNP dosage of CuStNPs (0.8 mg/mL) for 8 h, wherein cultures were serially diluted 10-fold in DI-H_2_O and plated on tryptic soy agar and allowed to grow for an additional 8–12 h for B. subtilis and 18–20 h for S. aureus prior to colony enumeration. The experiment was run in triplicate to allow for calculation of average logarithmic reductions and standard deviations. Statistical comparisons were evaluated using a two-tailed unpaired Student’s t-test.
4.4.3. Iodine Test for Starch Degradation
Bacterial cultures were exposed to 4.2 µg/mL CuNP dosage of CuStNPs (0.8 mg/mL) for 8 h, mixed 50:50 with 1 M iodine solution to evaluate the starch concentration in solution. Sample absorbance readings were taken at 580 nm [63].
4.4.4. Confocal Microscopy
Confocal microscopy was used to visualize bacteria-CuStNP interactions and evaluate bacterial viability in co-culture experiments. For bacteria-CuStNP interactions, fluorescein-labeled CuStNPs (dose 4.2 µg/mL CuNPs) were incubated with 10^8^ S. aureus or B. subtilis cells for 8 h. Samples were then transferred to confocal grade 8-well chambered coverglass wells (ThermoFisher 155411PK, Waltham, MA, USA) and viewed using a Nikon (Tokyo, Japan) A1RSi confocal laser scanning microscope, equipped with a CFI Plan Apo Lambda 100X oil objective with NA = 1.45 (Nikon, Tokyo, Japan). The excitation wavelength for fluorescein was 488 nm and imaging was captured with a fluorescein isothiocyanate (FITC) filter with emission 490–525 nm. For bacterial viability assessment of co-cultures, 10^8^ S. aureus cells were incubated with either 10^6^ or 10^8^ B. subtilis cells and fluorescein-labeled CuStNPs (dose 4.2 µg/mL CuNPs) for 8 h and stained using the LIVE/DEAD BacLight bacterial viability kit, (Molecular Probes Inc, Eugene, OR, USA). Samples (20 µL) were spotted onto confocal grade 8-well chambered coverglass wells (ThermoFisher 155411PK, Waltham, MA, USA) for imaging. The excitation wavelengths of the LIVE (SYTO 40) and DEAD (propidium iodide) dyes are 420 nm and 561 nm, respectively. Visualization was made by overlaid images with emission spectra for the live cells, dead cells, and CuStNPs of 435–485 nm, 570–620 nm, and 490–525 nm, respectively.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Russo C.A. Andrews R.M. The National Hospital Bill: The Most Expensive Conditions, by Payer, 2004 Healthcare Cost and Utilization Project (HCUP) Statistical Briefs Agency for Healthcare Research and Quality (US)Rockville MD, USA 200621938850 · pubmed ↗
- 2Rhee C. Dantes R. Epstein L. Murphy D.J. Seymour C.W. Iwashyna T.J. Kadri S.S. Angus D.C. Danner R.L. Fiore A.E. Incidence and Trends of Sepsis in US Hospitals Using Clinical vs Claims Data, 2009–2014 JAMA 20173181241124910.1001/jama.2017.1383628903154 PMC 5710396 · doi ↗ · pubmed ↗
- 3Current HAI Progress Report: 2023 National and State Healthcare-Associated Infections Progress Report 2024 Available online: https://arpsp.cdc.gov/profile/national-progress-2023/united-states(accessed on 1 October 2025)
- 4Abban M.K. Ayerakwa E.A. Mosi L. Isawumi A. The burden of hospital acquired infections and antimicrobial resistance Heliyon 20239 e 2056110.1016/j.heliyon.2023.e 2056137818001 PMC 10560788 · doi ↗ · pubmed ↗
- 5Odoom A. Donkor E.S. Prevalence of Healthcare-Acquired Infections Among Adults in Intensive Care Units: A Systematic Review and Meta-Analysis Health Sci. Rep.20258 e 7093910.1002/hsr 2.7093940636544 PMC 12239516 · doi ↗ · pubmed ↗
- 6Rogers E. Alderdice F. Mc Call E. Jenkins J. Craig S. Reducing nosocomial infections in neonatal intensive care J. Matern.-Fetal Neonatal Med.2010231039104610.3109/1476705090338702920718580 · doi ↗ · pubmed ↗
- 7Spellberg B. Guidos R. Gilbert D. Bradley J. Boucher H.W. Scheld W.M. Bartlett J.G. Edwards J.Jr. Infectious Diseases Society of America The epidemic of antibiotic-resistant infections: A call to action for the medical community from the Infectious Diseases Society of America Clin. Infect. Dis.20084615516410.1086/52489118171244 · doi ↗ · pubmed ↗
- 8Irfan M. Almotiri A. Al Zeyadi Z.A. Antimicrobial Resistance and Its Drivers—A Review Antibiotics 202211136210.3390/antibiotics 1110136236290020 PMC 9598832 · doi ↗ · pubmed ↗