Microbiota-Driven Immune Dysregulation Along the Gut–Lung–Vascular Axis in Asthma and Atherosclerosis
Elena-Larisa Zimbru, Răzvan-Ionuț Zimbru, Florina-Maria Bojin, Sorin Dan Chiriac, Laura Haidar, Minodora Andor, Gabriela Tănasie, Carmen Tatu, Marius Georgescu, Cristina Uța, Camelia-Felicia Bănărescu, Sabine Groza, Carmen Panaitescu

TL;DR
This paper explores how gut microbes contribute to both asthma and atherosclerosis through shared immune pathways, suggesting microbiota-targeted therapies could help treat both conditions.
Contribution
The paper introduces the gut–lung–vascular axis as a novel framework to explain shared inflammatory mechanisms in asthma and atherosclerosis.
Findings
Gut dysbiosis with reduced SCFA-producing bacteria and increased pathobionts links to inflammation in asthma and atherosclerosis.
Metabolites like SCFAs, TMAO, and LPS connect gut, lung, and vascular inflammation across human and experimental studies.
Microbial profiling and drug interactions offer potential for new diagnostics and microbiota-targeted treatments.
Abstract
Background: Asthma and atherosclerosis frequently coexist in clinical populations and share convergent immunometabolic pathways amplified by gut microbial dysbiosis. We propose the gut–lung–vascular axis as a unifying mechanistic framework connecting epithelial and endothelial inflammation providing a foundation for understanding shared inflammatory mechanisms beyond tissue-specific disease boundaries. Methods: A targeted narrative review systematically appraised clinical, experimental and multi-omics studies published over the last five years to delineate microbiota-driven pathways relevant to asthma and atherosclerosis. Particular emphasis was placed on specific microbial taxa, metabolite profiles and immunometabolic networks that connect gut dysbiosis with respiratory and cardiovascular dysfunction. Results: Across human and experimental cohorts, dysbiosis marked by depletion of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
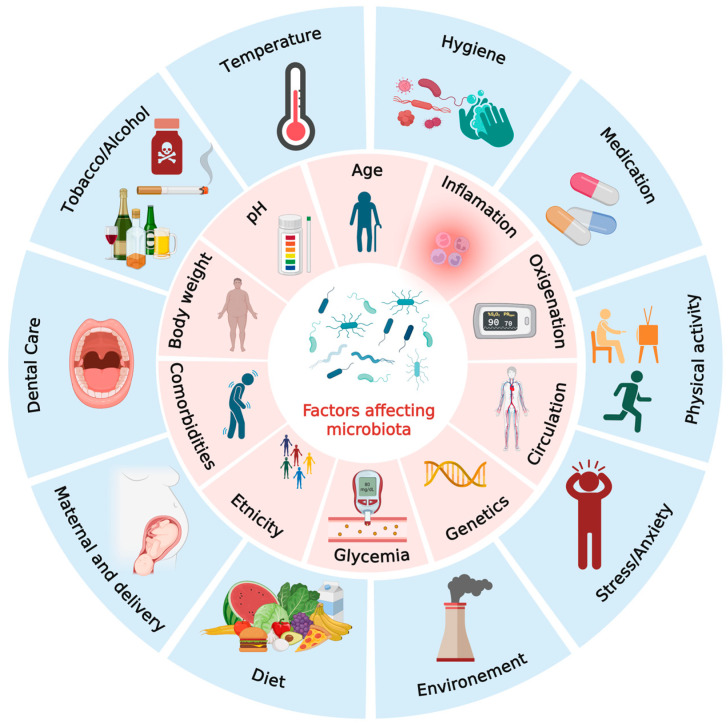 Figure 1
Figure 1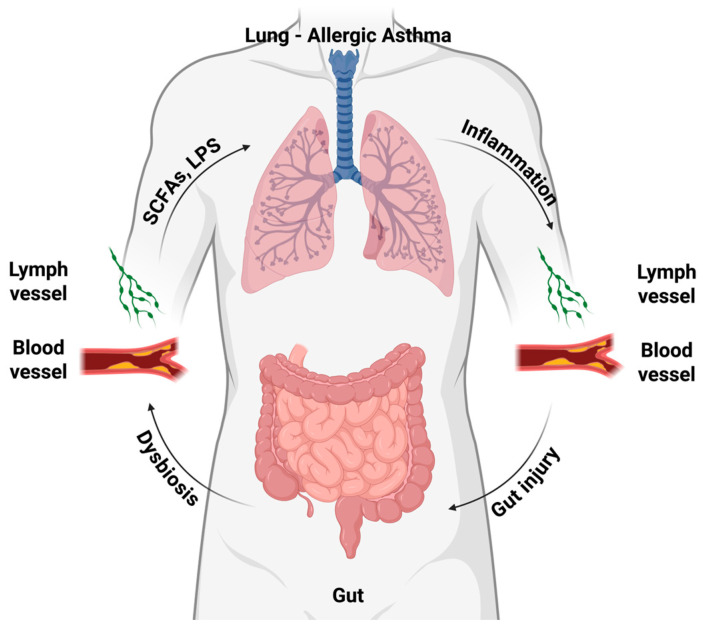 Figure 2
Figure 2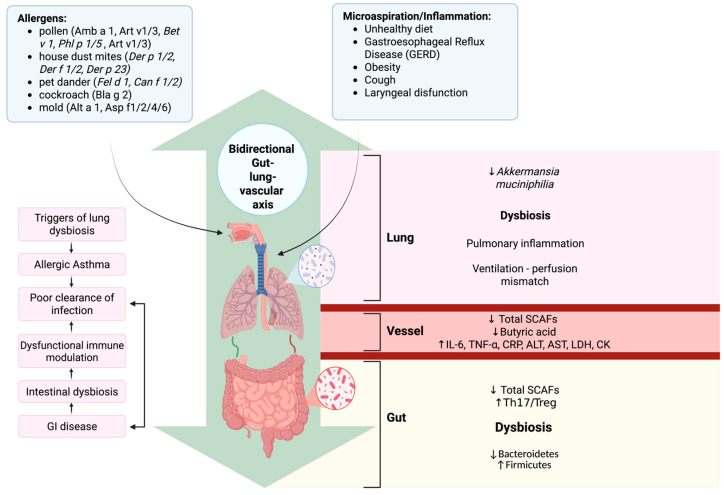 Figure 3
Figure 3| Condition | Key Microbial Taxa | Key Metabolites | Functional Role | References |
|---|---|---|---|---|
| Allergic Asthma |
| - | Enriched in airway microbiomes of children with asthma; associated with wheezing/exacerbations and disease severity. | [ |
| - | SCFAs | Linked with protection against allergic disease and dampen airway inflammation via Tregs/MDSCs. | [ | |
| Clostridia | SCFAs | Early-life depletion of SCFA-producing taxa is associated with atopy/asthma; restoration is protective. | [ | |
|
| SCFAs | Lower early-life abundance of these gut commensals predicts increased risk of wheezing and childhood asthma; their presence supports epithelial barrier integrity and tolerogenic immune programming. | [ | |
| - | Tryptophan metabolites (indole-3-carbinol, indole-3-acetic acid, kynurenine) | Activate AhR pathways to reduce airway inflammation and improve asthma outcomes. | [ | |
| - | Secondary bile acids | Bile acid-FXR/TGR5 signaling modulates lung immunity and suppresses type-2 inflammation. | [ | |
| Atherosclerosis | Firmicutes | SCFAs | SCFAs exert anti-inflammatory and vascular-protective effects; reduced SCFA tone linked to CVD risk. | [ |
| Enterobacteriaceae | TMAO | Microbial conversion of choline/carnitine to TMAO promotes atherosclerosis and adverse cardiovascular events. | [ | |
|
| LPS | Oral taxa present in plaques; endotoxin signaling (TLR4) drives vascular inflammation and lesion progression. | [ | |
| Enterobacteriaceae | Secondary bile acids | Microbiota-modified bile acids regulate lipid metabolism and vascular inflammation via FXR/TGR5 pathways. | [ | |
| Aromatic amino acid–metabolizing gut consortia | Phenylacetylglutamine (PAGln) | PAGln, derived from microbial phenylalanine metabolism, enhances platelet reactivity through adrenergic receptors and is associated with higher risk of major adverse cardiovascular events. | [ |
| Study (Year) | Model/ | Main Findings | Key Microbial/ | Proposed Mechanisms/ |
|---|---|---|---|---|
| Kim YC et al. (2024) | Human cohort review | Provides overview of clinical and experimental evidence demonstrating that gut microbial imbalance correlates with asthma severity and exacerbations. | Depletion of SCFA-producers ( | ↓ SCFAs → impaired Treg induction, enhancedTh2 inflammation and airway hyperresponsiveness. |
| Boulund U. et al. (2025) | Review | Microbial exposures during infancy and early gut colonization patterns are strong determinants of subsequent asthma susceptibility | ↓ | Early dysbiosis interferes with immune maturation → higher allergic sensitization and increased risk of childhood asthma. |
| Zheng XW et al. (2024) [ | Human cohort/genetics integration | Identified genetic links between gut microbial traits and asthma risk; complex taxa associations (some complex and sometimes inconsistent associations across taxa). | Context-dependent patterns, including reduced abundance of certain SCFA-producing genera but variable signals for taxa (e.g., | Indicates that host–microbe interactions in asthma are multifactorial and cannot be reduced to simple, linear taxon-to-effect relationships. |
| Ramar et al. (2025) | Mechanistic humanized/cell-report studies | Provide emerging causal evidence that defined gut microbial communities can shape susceptibility to asthma, particularly when hosts are exposed to environmental particulate triggers. | Specific live bacterial consortia were shown to modulate airway inflammatory responses in colonized hosts. | Supports causality: gut microbes can prime lung immunity through epigenetic pathways, such as DNA methylation changes in dendritic cells, thereby driving asthma-like phenotypes. |
| Study (Year) | Model/ | Main Findings | Key Microbial/ | Proposed Mechanisms/ |
|---|---|---|---|---|
| Zhu Y. et al. (2020) | Review integrating experimental and preclinical evidence | Proposed central role of TMA/TMAO pathway and other metabolites in promoting endothelial dysfunction and atherogenesis. | ↑ TMAO levels associated with increased activity of TMA-producing taxa such as Clostridia and | TMAO enhances foam cell formation, promotes vascular inflammation and contributes to pro-thrombotic remodeling of the vessel wall. |
| Li X. et al. (2021) | ApoE−/− mouse model | Antibiotic treatment or targeted modulation of the gut microbiota can alter atherosclerotic plaque burden by influencing TMA/TMAO production. | ↓ TMA-generating bacterial taxa accompanied by decreased circulating TMAO levels. | Lower TMAO limits macrophage foam cell formation and vascular inflammatory pathways → smaller atherosclerotic lesions. |
| Pala B. et al. (2024) | Human carotid intima–media thickness (IMT) cohort | Gut microbiome composition correlated with carotid IMT and plaque characteristics, linking microbial patterns to early vascular remodeling. | ↑ | Microbially derived LPS and related metabolites correlate with endothelial dysfunction and markers of subclinical atherosclerosis. |
| Mao Y. et al. (2024) | Review/multi-study synthesis | Integrative overview of how gut microbial alterations contribute to different stages of atherosclerosis, emphasizing the role of key microbial metabolites (TMAO, SCFAs, BA). | ↓ SCFAs (anti-inflammatory), ↑ TMAO and specific BA derivatives in patients with increased cardiovascular risk. | Metabolite shifts modulate vascular inflammation, lipid metabolism and immune cell recruitment, thereby shaping atherosclerotic disease progression. |
| Jarmukhanov Z. et al. (2024) | Systematic review/meta-analysis | ↑ TMAO consistently associated with higher CVD and atherosclerosis risk across multiple human cohorts. | ↑ circulating TMAO across cohorts; microbial taxa linked to TMA production (e.g., | Supports the concept of a microbe–metabolite–vascular risk pathway, with TMAO emerging as a prognostic biomarker for adverse cardiovascular outcomes. |
| Zhou Y. et al. (2024) | Mechanistic review/multi-omics | Multi-omics evidence links gut microbial metabolome (TMAO, BA) to vascular NF-κB and inflammasome activation. | ↑ TMAO and pro-inflammatory BA; | Activation of endothelial inflammatory signaling (NF-κB, NLRP3) → plaque development, progression and destabilization. |
- —Victor Babeș University of Medicine and Pharmacy, Timisoara
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Immune responses and vaccinations · Asthma and respiratory diseases
1. Introduction
Allergic asthma and atherosclerosis stand among the most prevalent chronic inflammatory diseases, each contributing significantly to global morbidity, mortality and economic burden [1,2]. Although asthma and atherosclerosis impact different organ systems, they share important immunological and pathological features, including persistent low-grade inflammation, activation of immune cells (mast cells, T cells), oxidative stress and endothelial dysfunction or epithelial barriers impairment [2,3]. Recent data, including research on microRNAs, suggest overlapping pathways that may mediate the link between respiratory and cardiovascular diseases, emphasizing the importance of targeted preventive measures and the development of personalized therapeutic strategies [4].
Beyond their individual impact, asthma and atherosclerotic cardiovascular disease (ASCVD) have been documented to co-occur within the same individuals. Clinical studies indicate that adults with persistent asthma exhibit higher rates of coronary heart disease, stroke and carotid atherosclerosis compared with non-asthmatic controls [2,3,5,6]. A recent systematic review and meta-analysis reported that individuals with asthma have a significantly elevated risk of composite cardiovascular disease and cardiovascular mortality, with summary relative risks around 1.3 for overall CVD and higher risks for specific outcomes such as angina pectoris, myocardial infarction and heart failure compared with non-asthmatic controls [7]. This augmented cardiovascular risk is particularly pronounced in adult-onset asthma and in women, supporting the existence of shared inflammatory and metabolic susceptibilities across these conditions [8].
Trillions of microorganisms in the gut perform fundamental metabolic and immunological functions, while dysbiosis has been associated not only with ASCVD but also with allergic asthma, suggesting that perturbations in gut microbial communities may drive systemic inflammation affecting both vascular and airway health [9]. We propose the concept of a gut–lung–vascular axis as a mechanistic framework linking asthma and atherosclerosis, suggesting that microbiome-driven immune dysregulation along this axis may contribute to the emergence of a novel, nontraditional cardiovascular risk factor, not yet integrated into current clinical paradigms. Therefore, the gut–X axis perturbations may constitute the initial modulation of pulmonary and vascular pathways, providing the systemic inflammatory background that drives the gut–lung–vascular axis [10]. The gut–X axis is at the core of bidirectional communication networks linking the gut microbiota with distant organs, such as the gut–lung, gut–brain and gut–vascular axes. These axes operate through coordinated immune, metabolic and neuroendocrine signaling pathways, and their disruption has been increasingly recognized as a contributor to the development and progression of chronic inflammatory and cardiometabolic diseases [11,12]. This conceptual framework emphasizes how microbiota-derived metabolites and immune mediators may act systemically to synchronize inflammatory responses across distant organs.
The human gut microbiota (GM) comprises approximately 3 × 10^13^ bacterial cells, predominantly belonging to taxa such as Bacteroidetes, Firmicutes, Actinobacteria, Proteobacteria, Fusobacteria, and Verrucomicrobia. These bacterial communities coexist and interact closely with other components of the gut ecosystem, including the mycobiota (fungae) and the viromes (mainly bacteriophages) [13]. Building upon this foundation, recent studies have introduced the concept of a “gut–vascular axis,” describing how gut microbial communities and their metabolites influence the development and progression of atherosclerosis [9,14,15]. Through intricate host–microbe interactions, the GM and its metabolic derivatives modulate vascular inflammation, endothelial integrity and function, as well as lipid metabolism, thereby preserving vascular health or facilitating atherosclerotic disease when disrupted [15,16].
Microbial communities extend beyond the well-known GM, colonizing other regions of the body, such as the oral cavity, skin, reproductive tract and respiratory tract, contributing to diverse functions in host regulation. These can also influence distal organs through systemic circulation, highlighting the interconnectedness of microbial ecosystems [17]. Despite differences in function, the gut and lungs share common developmental origins and mucosal architectures, with epithelial barriers and associated immune networks that maintain tissue homeostasis. Perturbations in the gut–lung axis can disrupt these networks, potentially exacerbating respiratory disease severity. In chronic respiratory conditions such as allergic asthma, the composition and function of the bacteriome, virome and mycobiome, along with their metabolites, are markedly altered compared to healthy individuals, although the precise mechanistic pathways remain incompletely defined [18]. Emerging evidence indicates that gut microbial dysbiosis may enhance susceptibility to allergic airway diseases via the gut–lung axis. Characterizing these host–microbiota interactions offers opportunities for the identification of disease biomarkers and the development of microbiome-targeted therapies for allergic asthma [19,20].
Disturbances in microbial composition and function, collectively referred to as dysbiosis, have been implicated in a wide spectrum of chronic inflammatory and metabolic disorders [15,19,21,22,23]. Emerging evidence emphasizes the role of microbiota-derived metabolites, such as short-chain fatty acids (SCFAs) and trimethylamine N-oxide (TMAO), in modulating systemic inflammation, vascular integrity and immune regulation [24,25,26]. The concept of the gut–lung and gut–vascular axis has become increasingly widespread, suggesting that dysbiosis in one compartment can exert distant effects on other organ systems [10,15,24,27,28].
A comprehensive literature analysis was performed using PubMed. To ensure methodological rigor and reflect current conceptual frameworks, the search focused on studies published within the preceding five years. Eligible sources included clinical, experimental and omics studies addressing host–microbiota interactions and immune signaling networks in asthma and atherosclerosis. Particular emphasis was placed on dysbiosis-related inflammation, alterations in microbial metabolites such as SCFAs and TMAO, and their subsequent effects across the gut–lung–vascular crosstalk.
This review synthesizes recent evidence revealing the immunometabolic interplay between allergic asthma and atherosclerosis through microbiota-driven mechanisms. By integrating insights from metagenomic, metabolomic and immunologic studies, we propose the gut–lung–vascular axis as a novel conceptual framework explaining how dysbiosis contributes to systemic inflammation and multi-organ pathology. Therapeutic strategies aimed at modulating the intestinal microbiome, through diet and lifestyle changes, targeted microbial supplementation, reducing environmental stressors, may be promising approaches for improving systemic inflammation and enhance clinical outcomes in both cardiovascular and respiratory diseases. Understanding this interconnected axis may guide precision strategies for microbiome-based prevention and therapy.
2. Microbiota: An Overview
Interactions between the host and its microbiota are essential for maintaining physiological balance [29]. Also, microbial metabolites, particularly SCFAs, support regulatory T-cell differentiation and anti-inflammatory pathways, while commensal species reinforce epithelial and endothelial barrier function [30,31]. When this equilibrium is disrupted, a state of dysbiosis arises, fostering systemic inflammation that can drive the development of diseases such as allergic asthma and atherosclerosis [24,32,33]. In asthma, microbial imbalance contributes to airway hyperresponsiveness and enhanced immune activation, whereas in atherosclerosis, it promotes endothelial dysfunction, immune cell recruitment and increased production of pro-inflammatory cytokines [28,34,35]. The gut microbiota, representing about 70% of the human microbial population, shows marked variation along the digestive tract due to chemical and immunological gradients [17,36,37]. Colonization begins at birth, shaped by delivery mode, breastfeeding and early-life exposures and shifts from facultative to strict anaerobes as the host matures [38,39]. The human microbiota is primarily composed of five dominant bacterial phyla: Firmicutes, Bacteroidetes, Proteobacteria, Actinobacteria and Verrucomicrobia [29,40,41,42,43]. Within these groups, Firmicutes (approximately 60–80%) include the classes Clostridia, Bacilli and Negativicutes, while Bacteroidetes (roughly 20–40%) encompass Bacteroidia, Flavobacteria, Sphingobacteria and Cytophagia, consisting exclusively of Gram-negative taxa. Generally, strict anaerobes such as Bacteroides, Clostridioides, Eubacterium, Ruminococcus, Peptococcus, Fusobacterium and Bifidobacterium dominate over facultative anaerobes including Lactobacillus, Escherichia, Enterobacter, Enterococcus, Proteus and Klebsiella [44,45]. Other phyla, such as Cyanobacteria, Fusobacteria and Spirochaetaceae, are present at lower abundance within the human microbial ecosystem [45,46]. Gut microbiota composition is influenced by host genetics and physiology, environment, diet, chronic disorders, medication use, physical activity, stress, circadian rhythm, habits (alcohol or tobacco use), maternal microbiota, early-life microbial encounters, hygiene, urban versus rural living and exposure to animals, with diet exerting the strongest impact through its effects on microbial diversity and metabolite production (Figure 1) [44,47,48,49,50,51,52,53,54].
2.1. Diet
Dietary composition represents one of the principal determinants of gut microbiota architecture and metabolic function, acting as a major factor governing host–microbiome homeostasis. Sustained dietary patterns modulate microbial diversity, metabolic function and inflammatory balance, thereby exerting profound and long-term effects on host physiology and susceptibility to chronic conditions [47,54].
The Mediterranean diet, characterized by high consumption of fiber-rich plant foods, legumes, polyphenol-containing fruits, whole grains, fish and extra-virgin olive oil, consistently promotes microbial diversity and metabolic resilience. Individuals following a Mediterranean diet typically exhibit increased abundances of Bacteroides, Prevotella, Lactobacillus, Faecalibacterium, Clostridioides and Oscillospira. This microbial pattern is associated with a higher Bacteroidetes-to-Firmicutes ratio and greater alpha-diversity, reflecting a more balanced and resilient gut ecosystem. The beneficial effects are largely attributed to the diet’s high fiber and unsaturated fatty acid content, which favor saccharolytic and anti-inflammatory bacterial communities [55,56,57].
In contrast, Western-style diets, typically high in saturated fats, refined carbohydrates, animal protein and food additives, are associated with reduced microbial diversity, increased Firmicutes and a decline in Bacteroides and expansion of pathobionts such as Enterobacteriaceae, Bilophila wadsworthia and Alistipes spp. Such dysbiotic configurations favor the production of pro-inflammatory metabolites (e.g., TMAO, lipopolysaccharides (LPS)) that impair gut barrier function and activate toll-like receptor-mediated inflammatory signaling, promoting metabolic endotoxemia and insulin resistance [58,59]. Similarly, high-fat/high-fructose diets have been shown in animal and human studies to disrupt intestinal tight-junction integrity, suppress SCFA synthesis and elevate oxidative stress, collectively contributing to obesity-associated inflammation and vascular dysfunction [60,61].
Other dietary patterns also elicit distinct microbial responses. Plant-based diets, encompassing vegetarian and vegan regimens, increase the abundance of fiber-degrading and saccharolytic taxa such as Prevotella and Ruminococcus, thereby enhancing SCFA output and reducing intestinal pH. The resulting metabolic environment supports colonocyte health and anti-inflammatory immune regulation [62]. In contrast, ketogenic diets, while clinically beneficial for metabolic disorders, are linked to decreased microbial diversity and reduced levels of Bifidobacterium and Eubacterium rectale, accompanied by elevated bile-acid metabolism and inflammatory gene expression [63,64].
Low-fiber diets negatively affect gut microbial composition by depleting fermentable substrates for beneficial taxa such as Faecalibacterium prausnitzii and Roseburia. Sustained fiber deficiency fosters dysbiosis, increasing susceptibility to metabolic and inflammatory disorders, including obesity, diabetes and atherosclerosis [65,66,67].
Diets high in fermented foods enhance gut microbial diversity and promote beneficial taxa such as Lactobacillus and Bifidobacterium. Their bioactive metabolites support epithelial integrity, increase SCFA production and reduce systemic inflammation. Regular consumption of fermented foods like yogurt, kefir, sauerkraut, pickles, sourdough bread or fermented beverages has been linked to improved immune regulation and metabolic health [68,69].
2.2. Medication Use
Pharmaceutical agents profoundly influence gut microbial composition and function, often producing long-lasting effects on host metabolism and immunity [70].
Recent evidence indicates that pharmacological treatments, including corticosteroids and antibiotics (used in asthma exacerbation treatment), can notably alter GM composition and metabolic activity. Statins, such as atorvastatin and rosuvastatin, have been shown to reshape gut microbial profiles by increasing beneficial taxa such as Bifidobacterium longum, Akkermansia muciniphila and Ruminococcus obeum, while reducing potentially pathogenic species like Parabacteroides merdae [71,72]. These changes may enhance SCFA production and bile acid metabolism, contributing to the anti-inflammatory and lipid-lowering effects of statins, while interindividual differences in microbiota composition may explain variable therapeutic responses [72,73]. Such disruptions in microbial diversity and metabolite production (e.g., SCFAs, secondary bile acids, TMAO, LPS) have been linked to impaired gut-barrier integrity, immune dysregulation and altered drug metabolism, ultimately influencing both disease progression and therapeutic efficacy [25,31,74].
Many routinely prescribed medications induce distinct shifts in gut microbial ecology, with some taxa, such as Streptococcus salivarius, increasing across multiple drug classes, while others displaying drug-specific signatures, including Bifidobacterium dentium with PPIs, Clostridium leptum with tricyclic antidepressants, Eubacterium ramulus with selective serotonin reuptake inhibitors (SSRIs) and a decrease in Bacteroides fragilis with metformin [75]. Other agents, such as steroid inhalers, statins, methotrexate, laxatives and L-thyroxine, also remodel microbial composition and functional pathways in predictable patterns [75].
Antibiotics remain the most potent disruptors, reducing microbial diversity, depleting beneficial taxa such as Bifidobacterium and Faecalibacterium prausnitzii and promoting the overgrowth of opportunistic pathogens and antibiotic-resistant strains. Broad-spectrum antibiotics have a very potent iatrogenic disruptor effect of the gut ecosystem, producing acute and occasionally long-lasting reductions in microbial diversity, loss of key commensals and changes in microbiome functional capacity. By eliminating susceptible taxa, they create microbial micro-environments that favor the overgrowth of opportunistic pathogens such as Clostridioides difficile, with varied clinical consequences that may result in life-threatening colitis [76,77]. Moreover, even short antibiotic courses can lead to persistent alterations in microbial metabolic capacity and resilience [78,79].
More broadly, metagenomic studies demonstrate that almost one-quarter of non-antibiotic medications exert antibiotic-like effects, suppressing gut bacterial strains and reshaping microbial composition [70].
Corticosteroids also modify the gut ecosystem by suppressing immune-regulated microbial niches and favoring the proliferation of pathobionts, contributing to increased intestinal permeability and systemic inflammation [80]. Oral glucocorticoid therapy markedly induces alterations in the gut microbiome, characterized by increased richness but reduced overall diversity. It leads to a higher Firmicutes/Bacteroidetes ratio, a signature frequently associated with metabolic dysregulation and induced consistent shifts at the genus level, including reductions in beneficial taxa such as Bifidobacterium, Faecalibacterium, Akkermansia and Prevotella, alongside a proliferation of Streptococcus, Collinsella and Parabacteroides [81]. Functionally, oral glucocorticoids impaired considerably the gut’s capacity to generate short-chain fatty acids, as confirmed by a reduced expression of key SCFA-producing enzymes and increased gut serotonin levels, reflecting broad metabolic rewiring [81]. It remains uncertain to what extent the inhaled corticosteroids reshape the diversity of specific bacterial taxa in the airways or in the gut, in the latter likely via the swallowed drug fraction and its systemic effects [73]. In severe asthma, high-dose inhaled corticosteroids modulate the pathological immune activation and airway inflammation. Their pharmacologic effects are associated with an enrichment of Proteobacteria and a reduced abundance of Bacteroidetes, Prevotella and Fusobacteria in the gut [82]. In parallel, a balanced intestinal microbiota has a reciprocal influence upon the immune cell activation, as outer membrane proteins from gut microbiota promote the formation and functionality of Tregs while inhibiting excessive Th2 and Th17 cell activation [83,84]. Consequently, the gut microbiota is essential for preserving immune homeostasis primarily by counterbalancing the detrimental impact of Th2/Th17 hyperactivation and beneficial effects of Tregs responses [83].
Proton-pump inhibitors (PPIs) are recognized as major and usually under-appreciated promoters of gut dysbiosis. Their use is associated with reduced bacterial abundance and changes involving up to 20% of detectable taxa, resulted from altering gastric pH and facilitating the survival and translocation of oral bacteria to the gut and increasing the abundance of Enterococcus, Streptococcus and Escherichia coli, which have been linked to dysbiosis-related disorders [85,86,87]. Sometimes, the microbiome impact of PPIs could exceed that of antibiotics or other common drugs [85].
Additionally, cardiovascular drugs such as statins, beta-blockers, ACE inhibitors and platelet aggregation inhibitors can modulate gut microbial metabolism, influencing bile acid transformation, SCFA production, TMAO and microbial diversity [73]. Altered gut microbiota and their associated metabolites are closely connected to the pathogenesis of atherosclerosis, dyslipidaemia, hypertension and heart failure, indicating that gut bacteria could biotransform cardiovascular drugs, consequently modifying their pharmacodynamics, pharmacokinetics and toxicity. For statins, pretreatment levels of microbiota-derived bile acids reliably predict the extent of LDL-cholesterol lowering, reflecting a bidirectional relationship in which microbial communities both influence and are modulated by lipid-lowering therapy [73,88]. In atherosclerosis, gut dysbiosis is marked by depletion of SCFA-producing Faecalibacterium and Roseburia and increase in Streptococcus, Escherichia and other opportunistic pathogens, enhancing TMAO production, NLRP3–IL-1 activation and endothelial dysfunction [40]. Statins can remodel gut microbial communities, typically shifting towards more SCFA-producing and BA-transforming taxa, while also lowering pro-atherogenic TMAO generating bacteria and inflammatory markers. Conversely, baseline dysbiosis characterized by reduction of SCFA-producing genera and a proliferation of pathobionts has been linked to attenuated LDL-cholesterol reduction and sustained vascular inflammation under statin treatment. Further supporting this, higher levels of SCFA-producing gut microbiota were reported during effective statin therapy [40,72,89]. Therefore, lipid control alone might not be a fully effective preventive strategy in some cases of atherosclerosis [40]. Furthermore, statin treatment in mice led to increased blood glucose levels and body weight, accompanied by specific alterations in the metabolic profile of the gut microbiota [72,75,90]. This pattern would indicate that lipid-lowering medications are partly microbiota-dependent and that the gut microbiota is both a target and a determinant of statin efficacy.
2.3. Age
Across the human lifespan, the gut microbiota experiences continuous and finely regulated transitions that mirror developmental, metabolic and immunological changes. The gut microbiome undergoes predictable transitions, from rapid colonization and ecological instability in infancy, through relative equilibrium in adulthood and finally to compositional decline in elderly [38,91,92,93]. Advancing age, is associated with a progressive decline in community diversity, reduced abundance of short-chain fatty acid-producing taxa and an increase in pro-inflammatory bacteria, fostering low-grade systemic inflammation and metabolic dysfunction, characteristic of “inflammaging” [94,95]. Large-scale cohort analyses have further linked these compositional shifts with frailty, immune dysregulation and cardiometabolic disease [96]. Evidence from studies of healthy aging populations indicates that gut microbiome decline is not inevitable. A large Chinese cohort found that healthy older adults, even beyond 94 years, shared microbiome profiles similar to young adults [97]. Centenarians often display higher microbial diversity, enriched in beneficial taxa such as Akkermansia, Christensenellaceae, Lachnospiraceae and Ruminococcaceae, suggesting links between microbial richness, resilience and longevity [98,99]. In contrast, frailty and cognitive decline are associated with reduced diversity, loss of Faecalibacterium prausnitzii and increased abundance of proinflammatory taxa including Eggerthella and Ruminococcus gnavus [100,101]. Despite these trends, inconsistencies across studies emphasize the observational nature of most microbiome-aging research and the need for mechanistic insight into how specific microbial shifts influence health outcomes in older adults [94].
2.4. Host Genetics and Physiology
Host genetic variation influences gut microbiota via bile acid metabolism. A murine study identified the Slc10a2 transporter as a key link between bile acid regulation and microbial composition [102]. Gut microbes transform primary bile acids into diverse metabolites that influence host signaling and microbial ecology [103]. The gut microbiome is partly regulated by host genetics, with heritability estimates ranging from 5–45%, though more recent studies suggest that only 3–13% of microbial taxa are heritable, indicating that genetics accounts for only a small fraction of inter-individual microbiome variation [104,105]. Twin and genome-wide studies have identified numerous host loci and at least 30 genes have been experimentally validated to influence microbial composition. Genetic variants can affect disease either directly, by altering host physiology, or indirectly, through microbiome-mediated mechanisms, emphasizing the need for integrative research linking host genetics, microbiota and disease phenotypes [104]. Although genetic effects are modest compared to environmental influences, they define a physiological framework within which diet, immunity and microbial ecology interact to maintain host-microbe homeostasis [106]. Probiotics can regulate miRNA expression, influencing immune balance and inflammation. The proposed microbiota–miRNA–lung axis suggests that gut microbes may epigenetically affect lung health. Combining probiotics with miRNA-modulating therapies (agomiRs and antagomiRs) could enhance anti-inflammatory effects, but more studies are needed to confirm these mechanisms and therapeutic benefits [4,107].
2.5. Maternal Microbiota and Early-Life Microbial Exposures
The maternal microbiota profoundly modulates neonatal immune ontogeny, with microbial transfer during gestation contributing to the programming of immune tolerance and predisposing or protecting against allergic asthma in later life [39]. Early-life microbial exposures, including delivery mode, breastfeeding and antibiotic use, further modulate immune tolerance, with dysbiosis in infancy predisposing individuals to chronic inflammation and heightened cardiovascular risk [38,91]. During vaginal birth, infants acquire microbes primarily from the mother’s vaginal and intestinal flora, whereas those delivered by C-section are mainly colonized by skin-associated bacteria [108]. Moreover, C-section and subsequent colonization with Clostridioides difficile have been linked to increased asthma risk, whilst vaginally born children were only pre-disposed if a parent suffered from an atopic disease. Probiotic supplementation has shown potential as an adjunctive approach for asthma prevention and symptom control [109,110]. Although defining a universal core respiratory microbiota remains challenging due to substantial individual variability, several genera, including Staphylococcus, Streptococcus, Haemophilus, Moraxella and Veillonella, are consistently detected within the respiratory tract [111].
Early-life gut and airway microbiota shape immune development and influence susceptibility to allergic asthma or ACVDs later in life [38,40,112]. Infants with reduced Bifidobacterium, Akkermansia, Faecalibacterium or fungal genera such as Candida and Rhodotorula are at higher risk of developing allergic diseases, whereas the presence of Faecalibacterium, Lachnospira, Veillonella or Rothia supports immune tolerance and modulates asthma risk [34,112,113]. These findings emphasize the importance of early microbial exposures and suggest that targeted interventions, dietary, probiotic and/or prebiotic, beneficially reshape the GM, enhance SCFAs synthesis and ameliorate systemic inflammation, providing potential benefits for both asthma management and vascular health.
2.6. Environmental Exposures
Environmental exposures such as hygiene practices, urban versus rural living and interactions with animals strongly shape early microbial colonization. For example, infants raised in rural households or on farms show greater colonization by beneficial bacteria like Bifidobacterium, Lactobacillus and Bacteroides, as well as higher diversity of anaerobic commensals, which is associated with lower allergy risk [114]. In contrast, children growing up in urban environments exhibit less microbial richness, altered gut and airway microbiota and increased susceptibility to asthma and sensitization [115]. Moreover, household exposures to animals and animal fecal contamination further modify the gut microbiome and resistome in early childhood, especially in less-sanitized environments [116]. A prospective analysis of over 200,000 participants from the UK Biobank found that urban environment features (air pollution, complex street networks) were associated with greater ASCVD risk, partly mediated by adverse cardiometabolic status and mental health pathways [117]. Additionally, recent studies indicate that higher levels of ambient particulate matter (PM_2.5_), nitrogen dioxide (NO_2_) and road traffic noise are linked to increased incident CVD risk, emphasizing these as key environmental risk factors [118,119].
2.7. Chronic Conditions
Recent evidence highlights the microbiota contribution to the pathogenesis of numerous chronic diseases, including autoimmune disorders, inflammatory diseases, metabolic syndromes and cancer, through its intricate crosstalk with the host immune system [120]. Dysbiosis has been associated with increased inflammatory responses and impaired metabolic regulation, as observed in some diseases such as type 2 diabetes and obesity [121]. Moreover, microbiota composition can influence tumor initiation, progression and therapeutic outcomes [122]. Modulation of gut microbial communities, through probiotics, prebiotics or bioactive compounds like phenolics, represents an emerging therapeutic strategy aimed at restoring eubiosis and improving clinical outcomes in complex chronic diseases [123,124].
2.8. Lifestyle Factors
Lifestyle factors significantly influence the gut microbiota: for example, higher levels of physical activity have been associated consistently with increased microbial diversity and favorable changes in microbial composition in numerous human cohorts [125,126]. Disruption of circadian rhythms and sleep–wake cycles alters the diurnal dynamics of microbial communities and has been shown to impair gut barrier function and microbial-derived metabolite production [127]. Habitual behaviors such as excessive alcohol consumption and smoking are linked to decreased microbial diversity, increased conditionally pathogenic abundance and heightened intestinal permeability, thus promoting dysbiosis and inflammation [128,129]. Physiological stress exerts pronounced effects on the gut microbiota through activation of the hypothalamic–pituitary–adrenal (HPA) axis, sympathetic nervous system and associated hormonal mediators, which in turn alter gut motility, secretions and barrier integrity [130]. In humans, higher levels of psychological stress are associated with reductions in microbial abundance and diversity, characterized by a reduction in beneficial genera such as Lachnospira, Sutterella, Lachnospiraceae, Phascolarctobacterium and Veillonella, alongside an increased prevalence of taxa including Methanobrevibacter, Rhodococcus and Roseburia [130,131]. Animal models further reveal that stress-induced increases in glucocorticoids and catecholamines promote shifts toward potentially pathogenic taxa and increased intestinal permeability [132,133,134]. Emerging evidence shows that microbiota changes can influence stress responses by altering HPA axis activity, creating a bidirectional gut–brain feedback loop under prolonged stress conditions [135].
3. Microbiota in Allergic Asthma
3.1. Evidence for the Gut–Lung Axis in Asthma
In recent years, the hypothesis of the existence of a "gut–lung axis" has become increasingly relevant in explaining the pathogenic mechanisms of asthma, pointing to the complex and bidirectional interaction between intestinal and pulmonary immunity [19,35,42,83]. Recent data suggest that alterations in the composition of the intestinal microbiota can influence inflammatory processes in the airways through microbial metabolites, bacterial components (such as endotoxins) and systemic immune regulation [136,137]. Prospective studies in infants have shown that delayed or abnormal development of the intestinal microbiota in the first years of life correlates with an increased likelihood of childhood asthma [112,138].
Studies have shown that fermentation of dietary fiber by Lactobacillaceae and Bifidobacteriaceae increases SCFAs, which suppress Th2-type inflammation partly via Treg induction [139,140]. Lactobacillus and Bifidobacterium enhance IL-10 and inhibit IgE-dependent responses, while probiotics also modulate Th17 pathways. DCs conditioned with Lactobacillus reuteri or Lactobacillus casei promote Th1 polarization and Treg expansion [141,142]. Probiotic use has likewise been associated with increased IFN-γ [143].
At the same time, analysis of the respiratory microbiome has shown notable differences between asthmatic patients and healthy individuals, indicating that dysbiosis in the respiratory tract may contribute to the perpetuation of chronic lung inflammation [43,144]. Taken together, this evidence supports the existence of a functional connection in the gut–lung axis, in which microbial communities at these levels interact and influence each other in the development of allergic respiratory diseases.
3.2. Microbial Taxa Associated with Allergic Asthma
Airway Microbiome
Changes in the composition and diversity of the airway microbiota are now widely recognized as important features of allergic asthma, reflecting both disease-related dysbiosis and immune–microbial interactions. The phylum Proteobacteria, with prominent genera such as Haemophilus and Moraxella, is often overrepresented in the respiratory tract of individuals with asthma. These taxa have been linked to epithelial barrier disruption and reduced responsiveness to corticosteroid therapy [144]. Certain members of the Firmicutes phylum, including Streptococcus species, may influence airway immune tone and inflammatory signaling, though their impact appears to vary depending on strain-specific characteristics and local microenvironmental conditions. In contrast, commensal Actinobacteria, notably Corynebacterium and Dolosigranulum, are associated with a stable, health-related airway microbiome. Their early colonization in infancy has been correlated with a lower risk of respiratory infections and allergic airway disease later in life, suggesting a protective role in maintaining epithelial and immune homeostasis [43].
Gut Microbiome
The gut microbiota exerts a profound influence on immune homeostasis and allergic sensitization. Reduced abundance of Bacteroidetes in infancy has been consistently linked to an increased risk of asthma development [31]. In contrast, Clostridia (class Firmicutes), including genera such as Faecalibacterium and Roseburia, are known producers of SCFAs that enhance regulatory T-cell function and promote immune tolerance [138]. These compositional patterns suggest that both local (airway) and distal (gut) microbial shifts are intertwined in shaping asthma susceptibility and phenotype expression.
Co-sensitization to ragweed pollen and house dust mites induces a more severe allergic asthma phenotype, especially in animals receiving a high-fructose diet, which exacerbates systemic inflammation, obesity, dyslipidemia and airway hyperresponsiveness. These changes are commonly associated with diet-induced dysbiosis and metabolic imbalance [145].
3.3. Mechanisms: Immune Modulation and Metabolite Signaling
Dysbiosis may disrupt immune homeostasis by promoting Th2 polarization (via increased IL-4, IL-5, IL-13 production) and diminishing regulatory responses. Conversely, specific commensals or their metabolites enhance Treg differentiation and suppress effector T-cell activation [146]. Dendritic cells and innate lymphoid cells (ILC2s) are also modulated by microbial signals, influencing antigen presentation, cytokine release and epithelial crosstalk [71].
Microbial metabolites are key mediators of the gut–lung axis.
SCFAs such as acetate, propionate and butyrate are fermentation products of dietary fibers by Clostridia species. SCFAs enhance Treg expansion, suppress airway eosinophilia and modulate gene expression through histone deacetylase (HDAC) inhibition and G-protein-coupled receptor (GPR41, GPR43, GPR109A) activation [146]. Early-life exposure to propionate, including through breast milk, has been associated with reduced airway inflammation, while short-chain fatty acids modulate immune responses through HDAC and activation of G-protein-coupled receptors [147,148,149]. Recent studies also show that butyrate can selectively inhibit Tfh13 cells involved in allergic sensitization [136,148,149].Tryptophan metabolites, particularly indole derivatives, act as ligands for the aryl hydrocarbon receptor (AhR), promoting epithelial integrity and limiting allergic inflammation [31]. Recent studies in asthma show that type-2 inflammation perturbs tryptophan metabolism and that exogenous indoles restore microbial diversity, reduce OVA-IgE/cytokines and ameliorate airway disease in vivo through AhR-dependent signaling [150].Secondary bile acids generated by gut microbial metabolism may also exert systemic immunoregulatory effects, though their contribution to asthma pathophysiology remains less well characterized [138]. Secondary BA supplementation has been shown to reduce allergen-driven airway inflammation, but evidence in human asthma remains limited and inconsistent [28].
3.4. Microbiota-Targeted Interventions: Probiotics, Prebiotics and Dietary Modulation
Emerging therapeutic strategies aim to restore microbial balance and enhance host-microbe interactions.
Probiotics: Supplementation with Lactobacillus, Bifidobacterium, Lachnospira or Akkermansia strains has shown potential to reduce airway inflammation and improve asthma control, though clinical results remain heterogeneous and strain-specific [151]. The PRObiotics in Pediatric Asthma Management (PROPAM) study evaluated the efficacy of a probiotic formulations containing Ligilactobacillus salivarius LS01 + Bifidobacterium breve B632. A reduction in both the incidence and severity of asthma exacerbations was observed among the children receiving probiotic supplementation [152]. Multistrain formulations containing Lactobacillus and Bifidobacterium species have been shown to enhance asthma symptom control and modulate inflammatory biomarkers, although their impact on lung function parameters remains variable across studies [151]. Recent review on gut–lung axis modulation identifies Lachnospira as a short-chain fatty acid-producing genus associated with protective, anti-inflammatory effects in asthma, though direct clinical evidence from supplementation trials remains limited [19].Prebiotics and dietary modulation: High-fiber diets enhance SCFA production, improve mucosal immune tolerance and reduce airway hyperresponsiveness in animal models [19]. In murine allergic asthma models, a high-fiber diet (or cellulose-enriched diet) attenuated airway inflammation and symptoms, altered gut microbial composition toward beneficial taxa and in some cases increased SCFAs [153]. Human observational data associate higher fiber intake with reduced biomarkers of airway inflammation and better asthma control, though prospective evidence is limited [154]. Increasing the intake of fermentable foods are being explored as adjuncts in asthma therapy with the aim of modulating the gut–lung axis via microbiota and metabolite pathways [19,155]. Higher fiber intake during mid-childhood appears to be associated with a lower risk of allergen sensitization later in life, although part of this association may reflect reduced exposure to dietary allergens due to food avoidance in sensitized individuals [154]. Dietary interventions targeting microbiota composition are being actively explored as adjunctive strategies for asthma management.Advanced approaches: Innovative microbiome-based approaches, including fecal microbiota transplantation (FMT) and helminth-derived immunomodulators, have recently attracted interest as potential adjunctive strategies for restoring immune balance in allergic asthma. Experimental studies indicate that transferring gut microbial communities from healthy donors can ameliorate airway hyperresponsiveness, reduce eosinophilic infiltration and normalize intestinal microbial diversity in murine asthma models, largely through the enhancement of SCFAs synthesis and expansion of regulatory T-cell populations [156]. Clinical evidence remains preliminary, with only limited human data and no proven efficacy in asthma management. Research on helminth-derived molecules shows immunoregulatory activity through suppression of Th2/Th17 inflammation, induction of IL-10-producing Tregs and modulation of TLR-mediated signaling [157,158].
Microbiota-based strategies hold promise for complementing standard asthma therapies by re-establishing immune balance along the gut–lung axis. However, further randomized controlled trials are needed to determine optimal microbial strains, dosage, patient phenotype, regiotype or endotype and treatment duration for clinical benefit.
4. Microbiota in Atherosclerosis
4.1. Role of the Gut Microbiota in Cardiovascular Disease
In recent years, mounting evidence has implicated the intestinal microbiome as a significant modulator of CVD, including the development and progression of atherosclerosis. The “gut–heart axis” concept emphasizes that alterations in gut microbial ecology may influence vascular health through metabolic, inflammatory and immune-mediated pathways [159].
Several human cohort studies and animal investigations demonstrate that dysbiosis, for example shifts in the relative abundance of major phyla such as Firmicutes versus Bacteroidetes, or expansion of potentially pathogenic taxa is associated with increased markers of subclinical atherosclerosis and coronary artery disease [160,161,162].
The gut microbiota may not only modulate classical risk factors (hypertension, dyslipidemia, diabetes mellitus and/or obesity) but may act directly on the vascular wall and atherogenic process [163].
4.2. Microbial Metabolites Implicated in Atherosclerosis
One of the central mechanistic links between the gut microbiome and atherosclerosis is the generation of biologically active microbial metabolites that enter the systemic circulation and act on vascular or immune targets.
SCFAs (acetate, propionate and butyrate) are major fermentation products of dietary fiber in the gut [164]. SCFAs exert pleiotropic cardiovascular effects, improving endothelial function and blood pressure control while attenuating systemic inflammation [164,165]. Recent studies indicate that increasing SCFAs levels attenuate risk factors for atherosclerosis by promoting eubiosis and reinforcing intestinal barrier integrity [164,165,166]. Dysregulation of acetyl-CoA metabolism, essential for SCFA synthesis, has been reported in hypertensive individuals, whereas Bacteroides acidifaciens-derived acetate and propionate have shown cardioprotective effects [167,168]. Likewise, butyrate-producing bacteria such as Roseburia intestinalis reduce vascular inflammation and ACVD by preserving mucosal integrity and limiting systemic endotoxin translocation [169,170]. SCFAs act primarily via G-protein-coupled receptors (OLFR78, GPR41 and GPR43) that regulate vascular tone and immune balance [148,171]. Propionate signaling through GPR41/OLFR78 elicits antihypertensive effects, whereas butyrate and acetate enhance nitric oxide bioavailability, supporting endothelial homeostasis [171,172,173]. High dietary fiber intake correlates with lower blood pressure and improved vascular outcomes in humans, in part due to the anti-inflammatory activity of SCFAs [174]. Butyrate maintains epithelial barrier integrity and inhibits histone deacetylases, leading to epigenetic suppression of pro-inflammatory mediators, thereby reducing cytokines such as IL-1β and TNF-α in experimental models [175,176,177]. However, excessive circulating SCFA levels, especially under high-protein/high-fiber diets, have been associated with unfavorable lipid profiles, indicating that SCFA effects depend on dietary composition and microbial context [178].TMAO represents one of the most extensively characterized gut–vascular co-metabolites [179]. Formed from choline, L-carnitine and phosphatidylcholine via microbial trimethylamine (TMA) synthesis and hepatic oxidation, elevated TMAO levels consistently correlate with higher atherosclerotic burden and cardiovascular risk [163,179]. Meta-analyses of large cohorts (>26,000 participants) reveal a dose-dependent association between TMAO and adverse cardiovascular outcomes [179,180]. Experimental evidence demonstrates that transplanting TMAO-producing microbiota into ApoE−/− mice accelerates plaque development, while TMAO suppression reverses these effects [181,182]. TMAO formation is diet-dependent, with omnivorous diets producing more TMAO than vegetarian or vegan diets [182]. Mechanistically, TMAO promotes atherosclerosis through foam cell formation, platelet hyperreactivity and increased inflammatory signaling [164,177,179]. Elevated TMAO levels associate with upregulated C-reactive protein, IL-1β and vascular nuclear factor kappa B (NF-κB) activation, contributing to plaque instability [177,179,183]. Microbial enzyme TMA lyase inhibition has shown promise in reducing TMAO-driven vascular injury [184,185].Bile acids (BAs) are another class of microbiota-influenced metabolites. They are saturated or hydroxylated steroids that aid absorption of dietary fats, lipophilic vitamins and metabolic regulation of lipids, glucose and systemic metabolic signaling [163,164,186]. Gut microbiota convert hepatic primary BAs into secondary BAs through deconjugation and dehydroxylation reactions, largely involving Lactobacillus, Bacteroides, Enterococcus and Clostridium spp. [164,177]. BA signaling through Farnesoid X-activated receptors (FXR) and G-protein-coupled bile acid receptors (TGR5) modulates cholesterol biosynthesis and inflammatory pathways relevant to CVD [177,187]. FXR activation regulates lipid metabolism and flavin-containing monooxygenase 3 (FMO3), a key enzyme in TMAO synthesis, whereas TGR5 activation exerts anti-inflammatory and anti-atherogenic effects via nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) inhibition [188]. In contrast, pregnane X receptor (PXR) signaling may enhance macrophage CD36 expression and lipid uptake, promoting plaque growth [189]. Emerging data point to altered bile acid profiles and gut microbiota-derived LPS/leaky gut endotoxaemia as additional mechanisms linking microbial ecology to vascular inflammation and cholesterol metabolism [177].Emerging evidence suggests that, in addition to hepatic bile acid synthesis, the gut microbiota actively participates in cholesterol metabolism by converting dietary or endogenous cholesterol into coprostanol, a sterol that is poorly absorbed and therefore eliminated in feces [177,190]. This transformation is primarily mediated by bacterial species from the Eubacterium and Bacteroides genera, such as E. coprostanoligenes and Bacteroides strain D8, although other microbial contributors are likely yet to be identified [190]. Experimental studies in animal models have shown that supplementation with coprostanol-producing bacteria can substantially lower plasma cholesterol concentrations, with effects persisting for several weeks after treatment discontinuation [191]. In contrast, human investigations remain inconclusive due to small study populations, limited microbial isolation success and demographic variability [192]. Moreover, the specific microbial genes and enzymes driving intestinal cholesterol conversion are not fully defined, emphasizing the need for further mechanistic studies to clarify the role of these pathways in cholesterol regulation and cardiovascular protection [192].Additional microbial metabolites, including succinate, imidazole propionate (ImP) and tryptophan (Trp) derivatives, also modulate vascular inflammation. Succinate acts as a pro-inflammatory ligand for SUCNR1/GPR91, stimulating HIF-1α, IL-1β and ROS generation [177,193]. Elevated serum succinate correlates with coronary inflammation and enhanced NLRP3 inflammasome activation, linking metabolic stress to vascular damage [177,194]. Similarly, ImP, produced from histidine by specific Clostridioides species, interferes with insulin and AMPK signaling, promoting endothelial dysfunction and metabolic inflammation [179,195]. Finally, Trp metabolism yields multiple bioactive compounds, such as indole-3-propionic acid (IPA) and kynurenine pathway intermediates, that engage aryl hydrocarbon receptor (AhR) signaling to regulate macrophage activity and immune tolerance [196,197,198,199]. Reduced IPA and Peptostreptococcus-mediated Trp conversion correlate with impaired ABCA1/miR-142-5p signaling, increased foam-cell formation and aggravated atherosclerosis [199].
4.3. Microbial Taxa of Interest
Gut microbiome: An elevated Firmicutes/Bacteroidetes ratio, indicative of dysbiosis, has been recurrently observed in patients with atherosclerotic disease and associated metabolic comorbidities [159]. Taxa in the Enterobacteriaceae family (e.g., Escherichia coli) are enriched in atherosclerosis and correlate with pro-inflammatory gene expression [200]. Veillonella and Streptococcus spp., which appear enriched in plaque samples and peripheral circulation, are emerging as potential biomarkers of atherogenic microbiota [201,202,203].Oral microbiome: Periodontal pathogens including Porphyromonas gingivalis and Aggregatibacter actinomycetemcomitans have been linked to vascular inflammation and atherosclerosis through mechanisms such as molecular mimicry, direct invasion of vascular tissues and induction of systemic inflammatory responses [40,204,205,206,207].
Several fundamental questions remain at the forefront of cardiovascular microbiome research. A primary objective is to determine whether distinct microbial and metabolomic signatures possess sufficient specificity and reproducibility to serve as prognostic or diagnostic biomarkers for atherosclerotic disease [161].
Equally important is the rigorous evaluation of microbiota-modulating interventions, including dietary modification, administration of prebiotics or probiotics and selective inhibition of microbial enzymatic pathways, with regard to their translational applicability, safety and efficacy in human populations [208].
Moreover, the intricate interdependence between host determinants such as genetic architecture, metabolic phenotype, environmental exposures and pharmacologic treatments and their collective influence on the gut–vascular axis, remains insufficiently elucidated. Addressing these unresolved issues will require comprehensive, longitudinal and multicentric investigations integrating advanced multi-omic profiling and high-resolution vascular imaging to delineate causal mechanisms and identify clinically actionable microbial targets for precision cardiovascular medicine [209].
5. Shared Mechanisms Linking Allergic Asthma and Atherosclerosis via Microbiota
5.1. Dysbiosis as a Common Inflammatory Driver
Recent studies indicate that allergic asthma and atherosclerosis, traditionally viewed as separate airway and vascular disorders, share microbiome-driven, immunometabolic and microRNA-mediated pathways along a gut–lung–vascular axis (Figure 2 and Figure 3, Table 1, Table 2 and Table 3) [4,138,161].
The Burkitt hypothesis, first proposed in the 1970s and later revisited in recent decades, postulated that the low incidence of “Western diseases” such as colon cancer, cardiovascular and inflammatory/metabolic disorders among rural African populations was attributable to their high dietary fiber intake [210,211,212]. Contemporary epidemiological and mechanistic studies have confirmed that fiber deficiency is strongly associated with increased risks of asthma and also atherosclerosis, largely through gut microbiota dysbiosis and reduced production of SCFAs, key metabolites with anti-inflammatory, immunomodulatory and barrier-protective properties [31,65,136,213]. Conversely, sustained consumption of fiber-rich diets enhances microbial fermentation, restores SCFA levels and supports systemic metabolic and immune homeostasis. Data suggest that increasing dietary fiber intakes by 15 g per day or to 35 g per day could extend lifespan, improve cardiometabolic health and substantially reduce long-term healthcare costs [214,215].
In both conditions, intestinal dysbiosis (reduced diversity and loss of beneficial commensals) associates with disease onset/severity in longitudinal and cross-sectional human studies, including pediatric cohorts for asthma and cardiometabolic cohorts for vascular disease [216,217,218].
5.2. Microbial Metabolites Bridging Lung and Vascular Physiopathology
A recurring signature of dysbiosis is reduced SCFA tone due to depletion of SCFA-producing taxa (e.g., Faecalibacterium, Roseburia) and low fiber intake. SCFAs promote epithelial barrier integrity, induce Treg differentiation through GPCR signaling and HDAC inhibition and dampen type-2/ILC2 and Th17-skewed airway inflammation; human cohorts link higher early-life or maternal SCFAs with lower risk of atopy/asthma [219,220]. Loss of barrier function increases translocation of microbe-derived products (metabolic endotoxemia), including LPS, which activates TLR-NF-κB signaling in distal tissues. In the airway, this fuels Th2/Th17 inflammation and bronchial hyperresponsiveness; in the vasculature, LPS/TLR signaling drives endothelial dysfunction, macrophage activation and foam-cell formation, which are central processes in atherogenesis [221,222].
Microbial metabolites also bridge gut and vessel biology. TMAO, generated from dietary choline/carnitine by gut microbes, is associated with adverse cardiovascular events in meta-analyses and cohort studies. It is also mechanistically linked to impaired cholesterol metabolism and increased vascular inflammation [223].
Beyond SCFAs and TMAO, BA signaling has emerged as a shared regulator. Microbiota-modified secondary BAs signal via FXR and the GPCR TGR5 on immune and barrier cells to restrain NF-κB-driven inflammation and remodel macrophage polarization; experimental and translational work suggests BA-receptor pathways influence pulmonary immunity and vascular tone, providing another conduit between gut metabolism and airway/vascular inflammation [220,224].
5.3. Convergent Immune and Cytokine Pathways
A convergent pro-inflammatory cytokine environment is increasingly recognized as a mechanistic bridge between airway and vascular pathology. Elevated levels of IL-6, IL-1β, IL-17, TNF-α and TGF-β orchestrate chronic immune activation and tissue remodeling within both pulmonary and vascular compartments. Through the activation of transcriptional regulators such as NF-κB and JAK/STAT, these cytokines amplify downstream inflammatory signaling, induce adhesion molecule expression and promote fibroblast and smooth muscle proliferation. The persistence of this signaling milieu favors structural changes in the bronchial wall and vascular endothelium, sustaining airway hyperresponsiveness and vascular dysfunction [225,226].
In both conditions, innate lymphoid cells (ILC2 and ILC3) and macrophage polarization toward pro-inflammatory M1 phenotypes amplify these cytokine cascades, perpetuating tissue infiltration and impaired resolution. The resulting oxidative stress, driven by NADPH oxidase, myeloperoxidase and mitochondrial ROS, induces endothelial activation and epithelial barrier dysfunction, key pathogenic events that contribute to both airway hyperresponsiveness and endothelial atherogenesis [227,228].
5.4. Microbial Signatures Across Respiratory and Vascular Systems
Respiratory ecosystems mirror these systemic links: enrichment of pathobiont-leaning genera such as Moraxella, Haemophilus and Streptococcus is repeatedly observed in airway microbiomes of children with asthma and relates to exacerbation-prone phenotypes, indicating that microbial community structure can amplify airway inflammation once systemic pro-inflammatory cues are present [229].
Oral–gut translocation may also contribute to vascular disease. DNA from oral taxa such as Streptococcus, Veillonella and Porphyromonas gingivalis has been detected in human atherosclerotic plaques, experimental studies show that Porphyromonas gingivalis can aggravate plaque biology via TLR-dependent effects on vascular cells and defective efferocytosis [201,205,206,230].
5.5. Integrative Concept: The Gut–Lung–Vascular Axis
Together, these observations support a unifying model: dysbiosis reduces SCFA-mediated immune tolerance and barrier protection, increases systemic exposure to microbial products (e.g., LPS) and pro-atherogenic metabolites (e.g., TMAO) and perturbs BA signaling, changes that converge on shared inflammatory nodes (TLR/NF-κB, Th2/Th17 programs) in the lungs and vessel walls [166,221,222]. Clinically, the confluence of these biological pathways is evident in the coexistence of asthma and atherosclerosis [7]. Individuals with asthma exhibit an increased burden of cardiovascular outcomes, including myocardial infarction, cerebrovascular events and markers of subclinical atherosclerosis, when compared with non-asthmatic populations [3,5,7,231]. Moreover, patients with inadequately controlled asthma frequently display a clustering of cardiometabolic disturbances, such as abdominal obesity, insulin resistance, atherogenic lipid profiles and persistent low-grade inflammation, which collectively exacerbate disease severity and compromise therapeutic responsiveness [2,35,228,232,233].
This pattern of multimorbidity reinforces the idea that the gut–lung–vascular axis provides an integrated explanatory model. In this context, disturbances in the gut microbiota are not limited to gastrointestinal or metabolic consequences, but can simultaneously influence airway inflammation and vascular pathology [177,203,218]. In this way, immune and metabolic signals derived from the microbiota can shape the evolution of respiratory and cardiovascular function in the same individual, providing a unifying mechanism behind the co-expression of asthma and ASCVD.
According to a recent article, a high-fructose diet aggravates metabolic, inflammatory and vascular dysfunction in allergic asthma, while rosuvastatin ameliorates dyslipidemia, systemic inflammation, endothelial reactivity and airway remodeling [234].
The microbiome provides interface modulating these immune and redox processes. Gut dysbiosis, characterized by the loss of Faecalibacterium prausnitzii, Roseburia and Bacteroides species, reduces SCFAs production, weakening Treg induction and enabling unchecked Th17/ILC3 activation [31,166,172,220,235]. Meanwhile, microbial metabolites such as TMAO and LPS trigger endothelial and macrophage TLR signaling, bridging intestinal dysbiosis to systemic vascular inflammation [24,161]. In parallel, airway microbiota alterations, including enrichment of Haemophilus, Moraxella and Streptococcus mirror inflammatory signatures of vascular dysbiosis, suggesting microbial–immune crosstalk between airway and cardiovascular compartments [43,144,164,165].
Complementary to the gut–lung–vascular interactions discussed in our manuscript, recent work on dermatological conditions also highlights the systemic impact of intestinal dysbiosis. A review on chronic spontaneous urticaria reports that reduced microbial diversity, depletion of SCFA-producing bacteria and expansion of Proteobacteria may contribute to increased intestinal permeability, systemic immune activation and mast cell dysregulation, emphasizing the broader relevance of microbiota-driven immune disturbances across organ systems [236].
6. Future Directions
Future research should aim to deepen our understanding of the intricate interplay between gut microbial communities and host immune responses along the gut–lung–vascular axis. Integrative multi-omics approaches, combining metagenomics, metabolomics, transcriptomics and proteomics, are essential for comprehensively mapping the dynamic interactions between the microbiota and host pathways in both asthma and atherosclerosis. For instance, studies of the gut–lung axis have already demonstrated that gut-derived metabolites and immune signatures modulate distant organ immunity, including pulmonary responses [27,83,250]. Likewise, in the context of cardiovascular disease, a growing body of evidence supports a gut–vascular axis, where microbial dysbiosis and microbial-derived metabolites such as TMAO and SCFAs influence endothelial function, inflammation and atherogenesis [159,160,165].
Furthermore, investigation into the gut–lung–vascular axis may elucidate shared mechanistic pathways and identify key microbial or metabolite mediators that link respiratory and cardiovascular pathology. Uncovering the mediators of cross-organ signaling (for example, gut-derived immune cells or metabolites migrating via the circulation or lymphatics) remains an important gap in the current knowledge.
Clinically, a major priority is validating specific microbial and metabolite signatures as biomarkers to identify individuals at increased respiratory and cardiovascular risk and to enhance current risk prediction models. Longitudinal studies combining microbiome profiling with clinical outcomes could enable better stratification of asthmatic patients at higher cardiovascular risk and cardiovascular patients prone to respiratory complications.
Therapeutically, microbiota-targeted interventions offer promising adjuncts to standard care. These include dietary strategies (high-fiber and fermented foods), carefully selected probiotic or synbiotic formulations, pharmacologic inhibition of microbial pathways involved in harmful metabolite production (such as TMA), and, in selected cases, fecal microbiota transplantation. In asthma, such approaches may help restore immune tolerance and reduce exacerbations, while in ASCVD they may improve lipid metabolism, endothelial function and vascular inflammation. Ultimately, integrating multi-omics technologies will be critical for matching patients to the most effective microbiome-based interventions, enabling personalized treatment strategies rather than uniform supplementation approaches.
Advancement of personalized, microbiome-targeted therapies, specific to individual microbial profiles and immune signatures, holds considerable promise for optimizing clinical outcomes and ameliorating disease progression. Recent reviews emphasize that precision microbiome interventions (e.g., probiotics, prebiotics, fecal microbiota transplantation) may become feasible, but their success will depend on the integration of multi-omics datasets and immune phenotyping [34,140,151,251].
Ultimately, combining these results may enable the development of preventive strategies, including dietary, lifestyle or microbial interventions, aimed at restoring microbial-host homeostasis and reducing systemic inflammation before the disease progresses to a clinically evident stage.
7. Conclusions
Current evidence shows that gut microbiota plays a unifying role in linking allergic asthma with atherosclerosis by influencing systemic inflammation, immune signaling and host metabolic pathways across the gut–lung–vascular axis. Distinct microbial patterns and metabolite profiles, such as SCFAs, TMAO and LPS, affect both airway and vascular function, revealing biologically convergent trajectories of disease development. These findings open new translational opportunities: microbiome-informed diagnostics, personalized therapeutic approaches and interventions that modulate microbial metabolites may offer innovative ways to prevent or ameliorate these coexisting disorders. Incorporating microbiome evaluation into clinical risk assessment and therapeutic planning could ultimately enhance patient management, creating a more integrated and personalized framework for individuals with overlapping respiratory and cardiovascular diseases.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wecker H. Tizek L. Ziehfreund S. Kain A. Traidl-Hoffmann C. Zimmermann G.S. Scala E. Elberling J. Doll A. Boffa M.J. Impact of Asthma in Europe: A Comparison of Web Search Data in 21 European Countries World Allergy Organ. J.20231610080510.1016/j.waojou.2023.10080537564904 PMC 10410582 · doi ↗ · pubmed ↗
- 2Aggarwal K. Bansal V. Mahmood R. Kanagala S.G. Jain R. Asthma and Cardiovascular Diseases: Uncovering Common Ground in Risk Factors and Pathogenesis Cardiol. Rev.20253321922610.1097/CRD.000000000000060037594265 · doi ↗ · pubmed ↗
- 3Hirata T. Asthma as Risk for Incident Cardiovascular Disease and Its Subtypes Hypertens. Res.2023462056205810.1038/s 41440-023-01328-537231166 · doi ↗ · pubmed ↗
- 4Zimbru R.-I. Zimbru E.-L. Bojin F.-M. Haidar L. Andor M. Harich O.O. Tănasie G. Tatu C. Mailat D.-E. Zbîrcea I.-M. Connecting the Dots: How Micro RN As Link Asthma and Atherosclerosis Int. J. Mol. Sci.202526357010.3390/ijms 2608357040332077 PMC 12026532 · doi ↗ · pubmed ↗
- 5Tattersall M.C. Guo M. Korcarz C.E. Gepner A.D. Kaufman J.D. Liu K.J. Barr R.G. Donohue K.M. Mc Clelland R.L. Delaney J.A. Asthma Predicts Cardiovascular Disease Events: The Multi-Ethnic Study of Atherosclerosis Arterioscler. Thromb. Vasc. Biol.2015351520152510.1161/ATVBAHA.115.30545225908767 PMC 4441553 · doi ↗ · pubmed ↗
- 6Cazzola M. Hanania N.A. Rogliani P. Matera M.G. Cardiovascular Disease in Asthma Patients: From Mechanisms to Therapeutic Implications Kardiol. Pol.20238123224110.33963/KP.a 2023.003836739655 · doi ↗ · pubmed ↗
- 7Jiang Y. Huang X. Yu D. Xu C. Wang Y. Wang X. Shen Y. Asthma and the Risk of Cardiovascular Diseases and Mortality: A Meta-Analysis of Cohort Studies Ther. Adv. Respir. Dis.2025191753466625133396510.1177/1753466625133396540272211 PMC 12035221 · doi ↗ · pubmed ↗
- 8Zhang B. Li Z.-F. An Z.-Y. Zhang L. Wang J.-Y. Hao M.-D. Jin Y.-J. Li D. Song A.-J. Ren Q. Association Between Asthma and All-Cause Mortality and Cardiovascular Disease Morbidity and Mortality: A Meta-Analysis of Cohort Studies Front. Cardiovasc. Med.2022986179810.3389/fcvm.2022.86179835369308 PMC 8968068 · doi ↗ · pubmed ↗