Dose- and Time-Dependent Modulation of Cx43 and Cx45 Expression and Gap Junction Conductance by Resveratrol
Gintarė Jančiukė, Rokas Mickus, Vytautas Raškevičius, Vytenis Arvydas Skeberdis, Ieva Sarapinienė

TL;DR
Resveratrol, a plant compound, affects gap junction communication in a dose- and time-dependent way, with different effects on two types of connexins.
Contribution
First comparative analysis of resveratrol's effects on Cx43 and Cx45 gap junction intercellular communication using direct conductance measurements.
Findings
Resveratrol inhibited Cx43 gap junction conductance but enhanced Cx45 conductance in HeLa cells.
Resveratrol caused dose- and time-dependent changes in connexin expression and plaque size.
Resveratrol reduced cell viability and altered Cx43 phosphorylation patterns linked to junction stability.
Abstract
Plant extracts are rich in various bioactive compounds, such as polyphenols, flavonoids, tannins, terpenoids, phenolic acids, saponins, alkaloids, and polysaccharides. Antioxidant polyphenols are increasingly attracting attention, not only as dietary components but also as valuable food industry byproducts. Resveratrol, present in a wide range of plants, is well recognized for its diverse biological activities, including antioxidant, antitumor, cardioprotective, and neuroprotective effects. Given the importance of intercellular communication in these physiological processes, gap junctions (GJs) composed of connexin (Cx) family proteins are of particular interest because they provide a direct pathway for electrical and metabolic signaling and are key players in maintaining normal organ function and cell development. Aberrations of GJ intercellular communication (GJIC) may result in the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
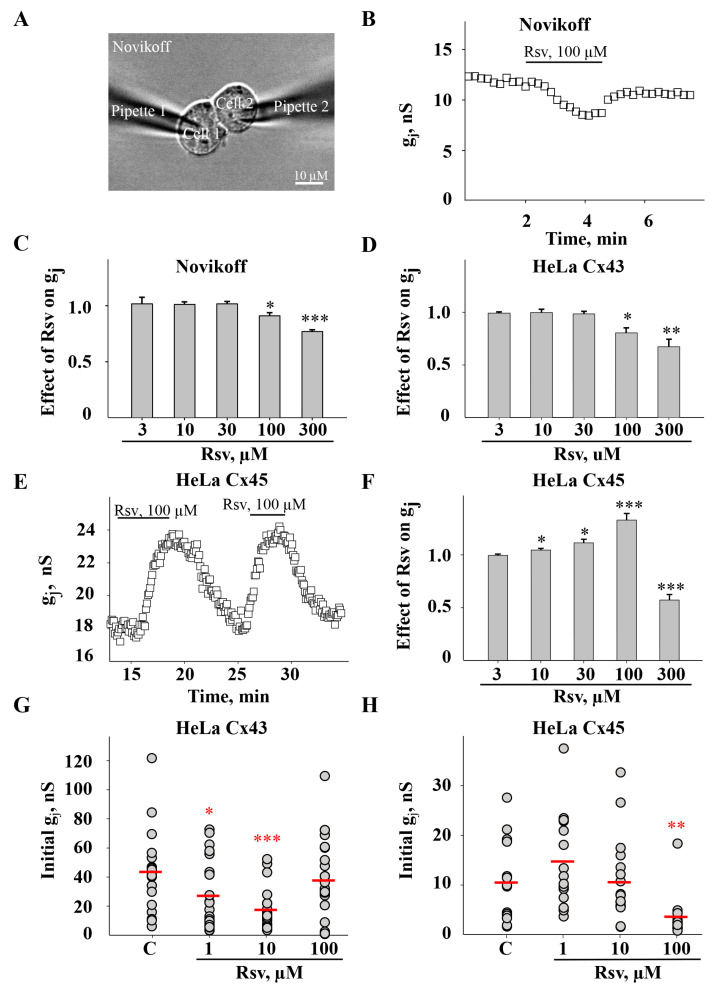 Figure 1
Figure 1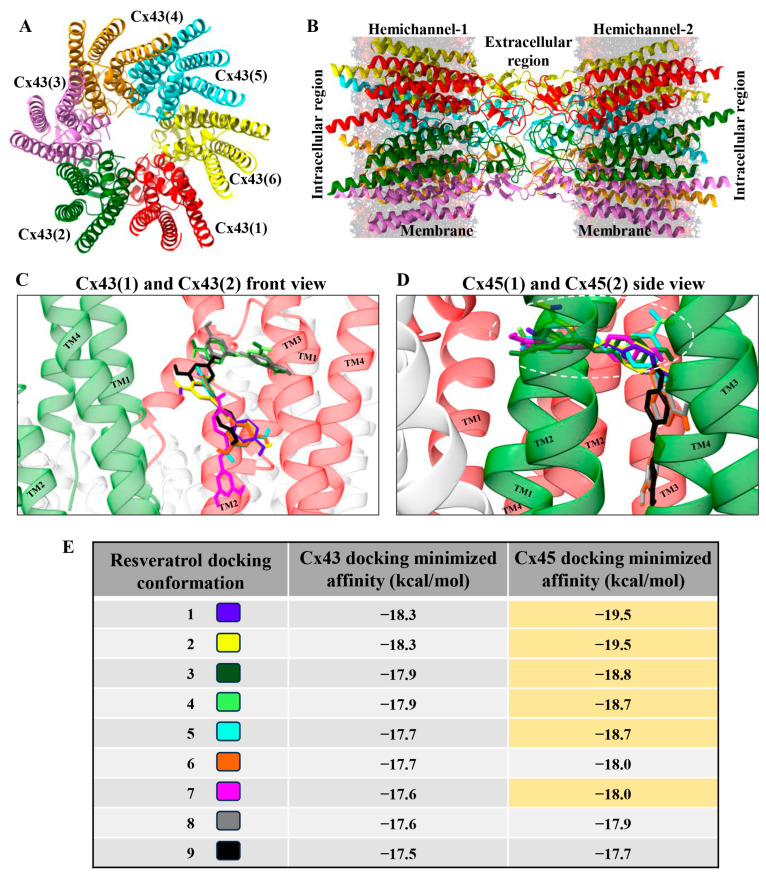 Figure 2
Figure 2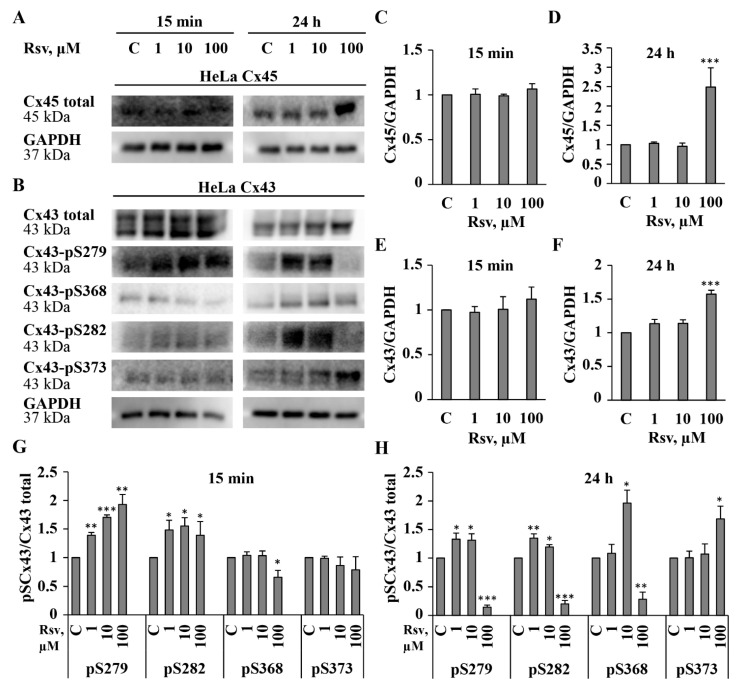 Figure 3
Figure 3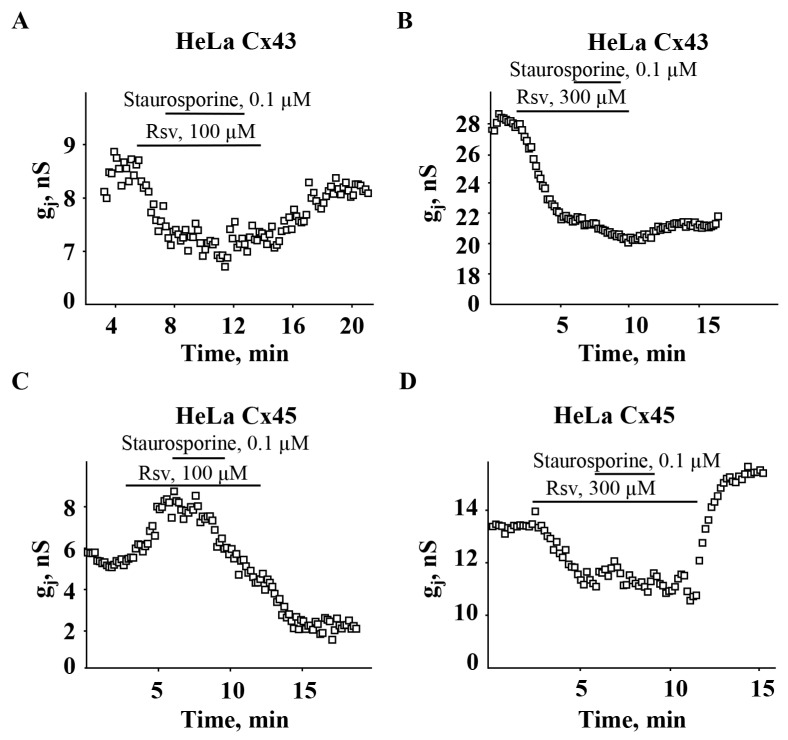 Figure 4
Figure 4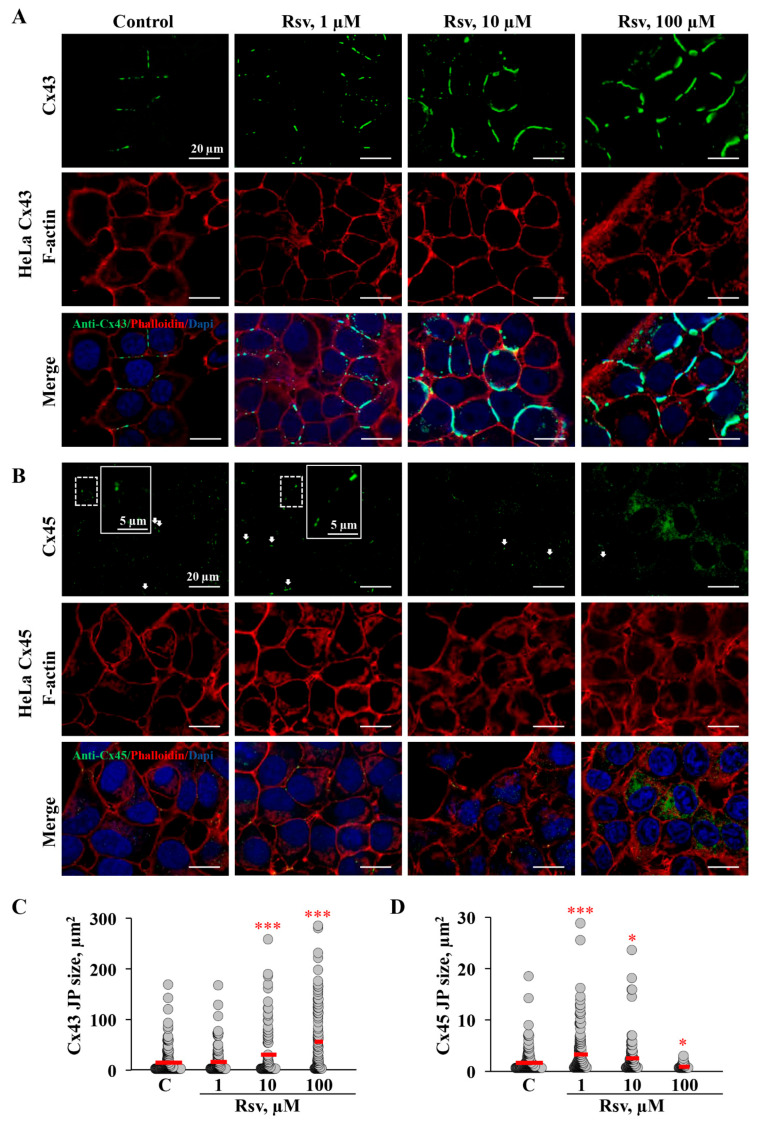 Figure 5
Figure 5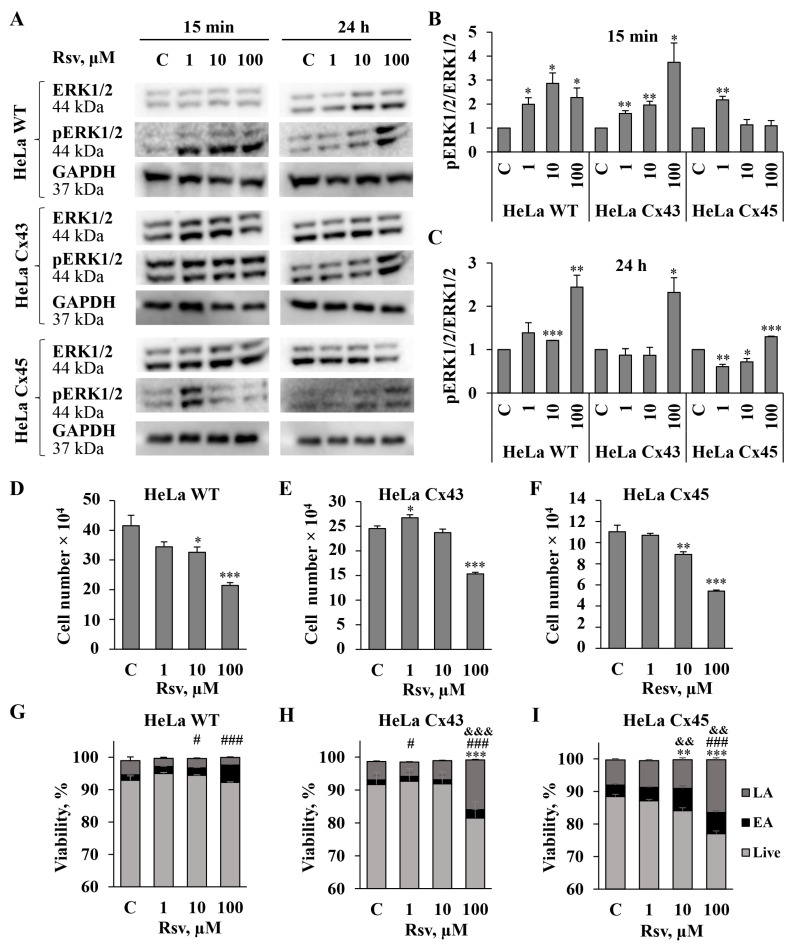 Figure 6
Figure 6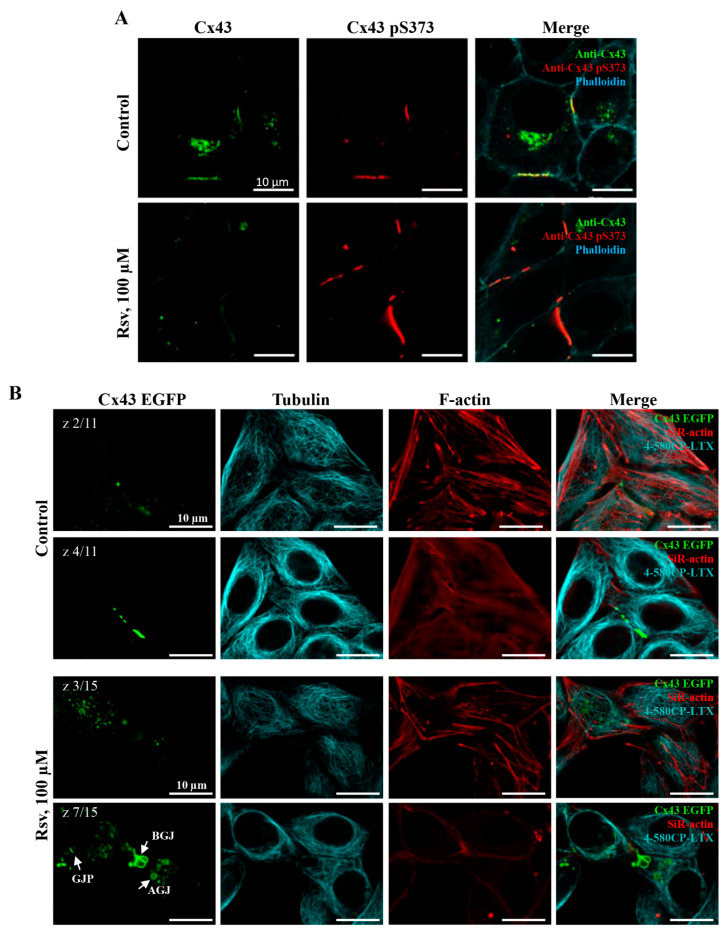 Figure 7
Figure 7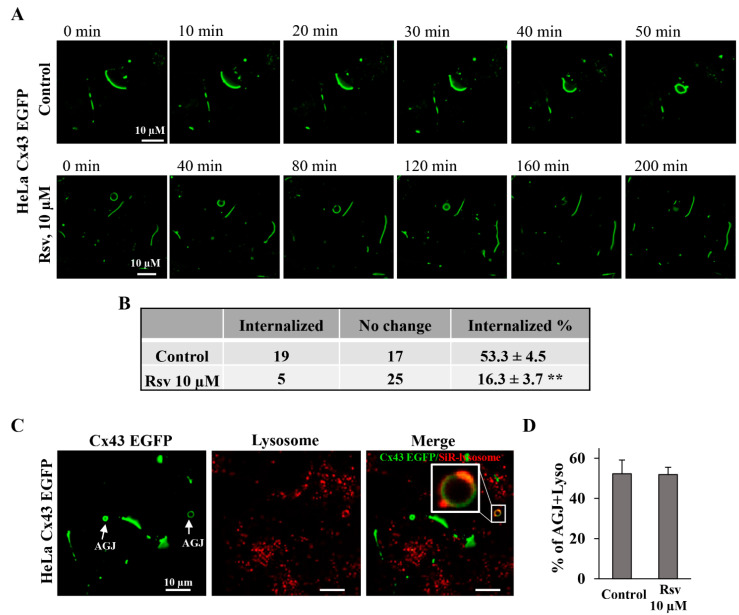 Figure 8
Figure 8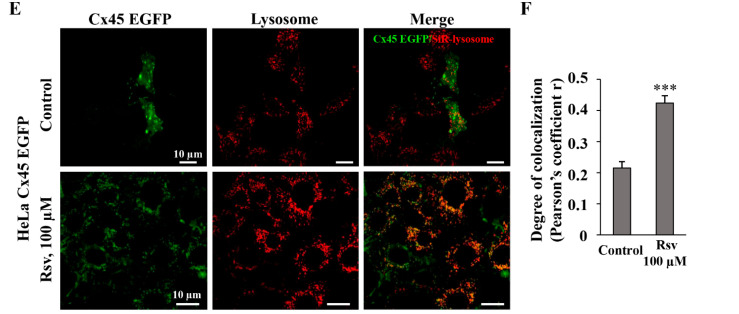 Figure 9
Figure 9- —Research Council of Lithuania
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsConnexins and lens biology · Ion Channels and Receptors · Barrier Structure and Function Studies
1. Introduction
Connexins (Cxs) are a large family of transmembrane proteins with four membrane-spanning domains, two extracellular loops, one intracellular loop, and intracellular N- and C-termini (CT) [1]. Cxs are synthesized in the endoplasmic reticulum, oligomerized in the Golgi, and delivered to the sarcolemma as hemichannels composed of six Cx subunits [2]. When hemichannels from adjacent cells align, they form GJs that enable electrical communication and the exchange of metabolites and signaling molecules [3]. Dense clusters of hundreds to thousands of individual channels form highly organized structures known as GJ plaques [4]. GJ conductance and permeability are modulated by factors such as junctional voltage, intracellular pH, divalent cations (Ca^2+^, Mg^2+^), phosphorylation, and various chemical agents [5,6]. Cxs interacting with cytoskeletal proteins, scaffolding molecules, and enzymes function not only as channels but also as signaling hubs that regulate cell behavior [7]. There are 21 known Cx isoforms in humans, classified by molecular weight and distribution in tissues [1]. Cxs differ mainly in the length of their CT that contains multiple phosphorylation sites; for example, human Cx43 has 33 phosphorylatable amino acid residues [8].
Cx43 is extensively studied due to its expression across the majority of tissues. It plays a key role in electrical communication in the heart and the brain [9]. Cx45 is also expressed in the heart and CNS neurons and is the first Cx to appear during embryonic development, marking early heart contractions [10]. In the heart, Cx45 localizes mainly in the conduction system, while Cx43 localizes in the working myocardium [11]. The role of Cx45 remains poorly explored, as knockout models are not viable postnatally.
Cx43 is described as one of the major players in tumorigenesis [12]. Tumor promoters often inhibit intercellular communication through GJs (GJIC), thereby promoting tumor growth, and the upregulation of Cx expression can suppress tumor formation [13]. However, contradictory data have also been reported, indicating that the inhibition of GJIC did not affect the proliferation and viability of Novikoff rat hepatoma cells expressing endogenous Cx43 and, under normal conditions, exhibiting large GJ conductance [14]. GJIC depends on several processes, including Cx synthesis, GJ channel formation, and pharmacological regulation.
Natural modulators such as resveratrol can induce a pro-apoptotic effect through the enhancement of GJIC [15,16]. Studies in rat liver epithelial cells showed that resveratrol can restore GJIC in a time- and dose-dependent manner [17]. Exposure time is a critical factor when evaluating the effects of resveratrol on cell viability. For example, treatment of liver stellate cells with 50 μM resveratrol for 24 h reduced cell viability due to oxidative stress; however, cytotoxicity decreased after 120 h, which suggests that surviving cells became more resistant to resveratrol-induced damage over time [18]. Alongside exposure time, resveratrol concentration is equally important. Studies in vitro use a wide concentration range of resveratrol, but the optimal dose in vivo requires further investigation in order to maximize its health benefits without toxicity. Resveratrol has been shown to produce anti-cancer effects in many in vivo and in vitro studies by inhibiting proliferation, inducing apoptosis, suppressing metastasis, and modulating the tumor microenvironment in colon, breast, prostate, and lung cancers [19]. It also enhances cancer cell sensitivity to chemotherapy and radiotherapy and can suppress multidrug resistance, making it a promising adjunct to conventional therapies [20,21]. Resveratrol is classified as a phytoestrogen due to its structural similarity to estrogen and its ability to interact with estrogen receptors (ER) [19]. It can bind to both ERα and ERβ as a mixed agonist/antagonist, influencing estrogen-responsive gene expression according to cell type, receptor subtype, and context [22]. For this reason, resveratrol should be used cautiously in cancer patients. However, the dominant biological activity of resveratrol is attributed to its potent antioxidant effects, through which resveratrol protects cells against oxidative stress-induced damage. Studies involving animal and cellular models demonstrate that resveratrol reduces reactive oxygen species production and lipid peroxidation while enhancing endogenous antioxidant enzyme activity across multiple disease models [23]. Resveratrol has been reported to prevent oxidative stress-induced cardiac hypertrophy by protecting liver kinase B1 and restoring activated protein kinase activity, suppressing pro-hypertrophic signaling in both in vitro cardiomyocytes and in vivo hypertensive rat models [24]. In a rat model of Parkinson’s disease induced by 6-hydroxydopamine, resveratrol demonstrated antioxidant effects by decreasing reactive oxygen species levels, reducing lipid peroxidation, and preserving glutathione content, alongside improved antioxidant enzyme activities in brain tissue [25].
Although resveratrol is most studied for its antioxidant properties, increasing evidence suggests that its effects on the cell extend beyond the regulation of redox balance alone. Oxidative status is closely linked to intercellular communication [26,27]; therefore, the influence of resveratrol on cell–cell communication, particularly through connexins and GJs, has become an important area of research. While much is known about the impact of resveratrol on Cx43 expression, data on its direct effect on Cx43 GJ conductance remain scarce. Resveratrol has been shown to enhance GJIC by upregulating Cx43 expression and function in various cell types. In retinal pigment epithelial cells under high glucose, resveratrol prevented Cx43 downregulation, restored GJIC, and reduced inflammation [28]. In melanoma, resveratrol increased Cx43 expression levels and GJIC, enhancing chemosensitivity [29]. Resveratrol also counteracted GJIC inhibition caused by tumor promoters or oxidative stress by blocking Cx43 and extracellular signal-regulated protein kinase 1/2 (ERK1/2) phosphorylation [30]. ERK1/2 phosphorylation is a crucial step in the mitogen-activated protein kinase (MAPK) pathway, playing a fundamental role in regulating key cellular processes such as proliferation, differentiation, survival, growth, metabolism, migration, and gene expression. The effects of resveratrol on Cx45-dependent GJIC are much less explored. While substantial evidence exists on the impact of resveratrol on ERK1/2 signaling, studies directly linking resveratrol, ERK1/2, and Cx45 are lacking in the current literature. In HeLa cells expressing exogenous Cx45, resveratrol, vitamin E, and tempol were found to reduce hemichannel opening. Although resveratrol appears to inhibit Cx45 hemichannel activity, likely due to its antioxidant properties through reducing free radical generation and potentially protecting against oxidative stress [31], there are currently no data on its effects on Cx45-mediated GJIC.
Previous studies have mostly focused on changes in Cx phosphorylation, expression, or GJIC assessed by molecular assays or dye transfer methods, rather than direct electrophysiological measurements. Therefore, the aim of this study was to improve the understanding of the dose- and time-dependent effects of resveratrol in cells expressing different types of Cxs by combining several approaches, such as dual whole-cell patch-clamp, molecular modeling, time-lapse imaging, Western blotting (WB), and fluorescence microscopy.
2. Materials and Methods
2.1. Cell Lines and Culture Conditions
Experiments were conducted on HeLa cells (ATCC CCL-2) stably transfected with either mouse or rat wild-type Cx45 and Cx43, or with tagged enhanced green fluorescent protein (EGFP). Additionally, Novikoff rat hepatoma cells expressing endogenous Cx43 were used. All cell lines were generously provided by the late Dr. F. Bukauskas (Albert Einstein College of Medicine, New York, NY, USA). Cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM) containing 10% fetal bovine serum and a mixture of 100 U/mL penicillin and 100 μg/mL streptomycin (Sigma-Aldrich, Steinheim, Germany) and maintained at 37 °C in a humidified atmosphere with 5% CO_2_. trans-resveratrol (Cat no. R5010) and all other reagents were purchased from Sigma-Aldrich.
2.2. Molecular Docking
Molecular docking was carried out as described previously [32]. The human Cx43 structure (PDB ID 7F94) [33], featuring the I164V mutation incorporated using ChimeraX software (v. 1.6.1) [34], was adapted to represent the rat Cx43 (rCx43) structure for molecular docking. The resultant Cx43 model underwent validation by ProSA [35] and Procheck (v. 3.5.4) [36]. The mouse Cx45 (mCx45) structure was modeled with AlphaFold 2 and validated with an internal validation score [37]. The 3D molecular structure of resveratrol was downloaded from the PubChem database [38]. Molecular docking was performed using Utilizing Smina software (v. Oct 15 2019) [39] alongside a customized Vina scoring function [40]. To prevent the placement of symmetric docking conformations within neighboring subunits, the docking software was configured to use two adjacent Cx43 hemichannel subunits. The random seed was steadily set to one. The most favorable nine docking conformations were recorded. Two-dimensional docking images were created with LigPlot+ (v. 2.2.9) software [41].
2.3. Electrophysiological Measurements
For electrophysiological recordings, cells grown on glass coverslips were transferred to a continuously perfused experimental chamber mounted on the stage of an inverted microscope Olympus IX81 (Olympus Europa Holding GmbH, Hamburg, Germany) equipped with an Orca-R2 cooled digital camera (Hamamatsu Photonics K.K., Hamamatsu, Japan). Junctional conductance g_j_ between adjacent cells was determined using the dual whole-cell patch-clamp technique, as previously described [42]. Before measuring the initial g_j_, cells that were incubated with resveratrol for 24 h were perfused for 30 min with a control solution.
2.4. Flow Cytometry Assay
Initially, 5 × 10^4^ cells were seeded in a 12-well plate per well in 1 mL of complete medium. Cells were treated with resveratrol for 24 h, then washed twice with PBS, trypsinized, and collected by centrifugation. Subsequently, cells were maintained with Guava Nexin^®^ Reagent (Millipore, Burlington, MA, USA) at room temperature for 20 min according to the manufacturer’s protocol. Analysis of the samples was performed using a Guava^®^ easyCyte™ HT flow cytometer (Millipore, USA), and the data were analyzed by GuavaSoft 2.2.3 InCyte software.
2.5. Western Blotting
Cells were lysed on ice using cell extraction buffer (Invitrogen, Waltham, MA, USA) containing 1 mM PMSF (Abcam, Cambridge, UK), 20 μL/mL protease inhibitor cocktail, and 10 μL/mL phosphatase inhibitor cocktail (both from Sigma-Aldrich, Steinheim, Germany) for 30 min. Samples were centrifuged at 4 °C for 10 min at 13,000 rpm. Total protein levels were quantified with a Qubit^®^ protein assay kit and a Qubit 3.0 fluorometer (Invitrogen, USA). Protein samples were resolved by Bolt™ 4–12% Bis-Tris plus gels (Invitrogen, USA) and transferred onto PVDF membranes (Millipore, USA), then incubated for 1 h with primary antibodies against Connexin-45 (40–7000; Invitrogen, USA), Connexin-43 (C6219; Sigma-Aldrich, Steinheim, Germany), phospho-Connexin-43 (Ser368) (PA5-118544; Invitrogen, USA), phospho-Connexin-43 (Ser282) (PA5-64641; Invitrogen, USA), phospho-Connexin-43 (Ser279) (PA5-64640; Invitrogen, USA), phospho-Connexin-43 (Ser373) (PA5-64670; Invitrogen, USA), ERK1/ERK2 (MA5-15134; Invitrogen, USA), phospho-ERK1/ERK2 (MA5-15173; Invitrogen, USA), and GAPDH (AM4300; Invitrogen, USA). Proteins were detected with the WesternBreeze^®^ chemiluminescent kit (Invitrogen, USA) according to the manufacturer’s protocol. Band signals were captured using the G:Box Chemi Gel Documentation system (Syngene, Baltimore, MD, USA).
2.6. Immunocytochemistry
Cells were cultured on sterile glass coverslips and fixed for 15 min at room temperature in 4% paraformaldehyde. Cells were washed and then permeabilized with 0.2% Triton X-100 in PBS for 3 min. Next, samples were exposed to the primary antibodies for 1 h at 37 °C, diluted in 1% BSA/PBS. After washing with 1% BSA/PBS, cells were maintained at 37 °C for 30 min with species-specific secondary antibodies conjugated to distinct fluorophores: Alexa Fluor 488 and Alexa Fluor 647 (all at 1:125, Life Technologies, Carlsbad, CA, USA). F-actin was labeled by incubating cells with Alexa Fluor™ 594 phalloidin (A12381; Invitrogen, USA) for 30 min. Coverslips were mounted using a Vectashield Mounting Medium with DAPI (Vector Laboratories, Newark, CA, USA). The samples were analyzed with an Olympus IX83 inverted confocal microscope (Olympus Europa Holding GmbH, Hamburg, Germany). For the estimation of GJ plaque size, 3D images were obtained as Z-stacks at 0.25 μm intervals. The estimation of GJ plaque surface area (µm^2^) was performed using Fiji/ImageJ 3D Object Counter software (Version 1.54), where at least 3 zones were analyzed from 3 biological replicates, and no less than 100 GJ plaques were analyzed for each condition.
2.7. Live Cell and Time-Lapse Imaging
Live-cell imaging was carried out at 37 °C with 5% CO_2_ using an incubation system INUBG2E-ONICS (Tokai Hit, Shizuoka-ken, Japan) with an incubator, mounted on the stage of an Olympus IX83 inverted confocal microscope (Olympus Europa Holding GmbH, Hamburg, Germany). HeLa cells were seeded onto glass-bottom dishes and maintained in full DMEM. For the connexin/lysosome colocalization assay, HeLa Cx45-EGFP and HeLa Cx43-EGFP cells were labeled with SiR-Lyso 1 h prior to imaging. Data were assessed from 3 biological replicates. AGJ-lysosome quantification was performed by analyzing 20 AGJs per replicate. Cx45/lysosome colocalization was assessed by analyzing 20 cells per replicate. Analysis was performed using Fiji/ImageJ Colocalization software (Version 1.54). For cytoskeletal images, HeLa WT, HeLa Cx43, Hela Cx45, and HeLa Cx43 EGFP cells were labeled with SiR-Actin and 4-580CP-LTX fluorescent probes. Time-lapse imaging for GJ plaque internalization was automatically set to be taken at 10 min intervals. Images were collected as Z-stacks using 0.5–1 μm steps. Data were assessed from 4 biological replicates.
2.8. Statistical Analysis
Experimental data are presented as means ± standard error of the mean. Statistical comparisons were conducted using a two-tailed Student’s t-test, with p-values < 0.05 considered statistically significant. All statistical analyses were performed with SigmaPlot version 12.0 software.
3. Results
3.1. Resveratrol Differently Modulates Cx43 and Cx45 GJ Conductance
We used the dual whole-cell patch-clamp technique (Figure 1A) to assess the effect of resveratrol on GJ conductance (g_j_) between Novikoff cells expressing endogenous Cx43 and HeLa cells expressing exogenous Cx43 and Cx45. Resveratrol had no effect on Cx43 GJ g_j_ at low doses (3, 10, 30 µM) and inhibited it at higher concentrations (100 and 300 µM) in both Novikoff and HeLa Cx43 cells (Figure 1B–D). In contrast, in HeLa cells expressing Cx45, resveratrol stimulated the g_j_ at 10 and 100 µM but inhibited it at 300 µM (Figure 1E,F). To assess the long-term effect of resveratrol, we preincubated cells with it for 24 h and measured the initial Cx43 and Cx45 GJ g_j_ in HeLa cell pairs right after the initiation of patch-clamp conditions. Surprisingly, a decrease in initial Cx43 GJ g_j_ was observed after treatment with resveratrol at lower concentrations (1 and 10 µM), while 100 µM had no effect (Figure 1G). Meanwhile, resveratrol reduced initial Cx45 GJ g_j_ only after incubation with 100 µM (Figure 1H). The effect of 300 µM resveratrol on g_j_ after cell incubation for 24 h could not be precisely assessed, as it drastically affected cell viability.
3.2. Docking of Resveratrol to Cx43 and Cx45
As shown in Figure 1, resveratrol exerted different effects on Cx43 and Cx45 GJ g_j_. At low concentrations, it stimulated only Cx45 GJ g_j_ but at higher doses it inhibited both Cx45 and Cx43 GJs. Therefore, we presumed that Cx45 should exhibit two different docking profiles, the first including high-affinity docking sites responsible for the stimulation of g_j_, and the second including low-affinity docking sites responsible for the inhibition of g_j_. To verify this hypothesis, we performed molecular docking experiments. Figure 2A represents a top view of the GJ hemichannel composed of six Cx43 subunits (subunit number is indicated in parentheses), and Figure 2B shows a side view of the GJ channel composed of two apposed hemichannels. We analyzed the nine most favorable docking conformations and found that they all localized on the outer surface of Cx43 TM2 and TM3 (Figure 2C and Supplementary Movie S1) with minimized docking affinities varying from −18.3 to −17.5 kcal/mol (more negative values indicate stronger interaction between resveratrol and connexin) (Figure 2E). However, only three docking sites on Cx45 localized on its outer surface with minimized affinities from −18.0 to −17.7 kcal/mol, while the remaining six ones were situated in the niche between two neighboring Cx45 subunits towards the channel pore (Figure 2D and Supplementary Movie S2), and the docking affinities of five of them varied from −19.5 to −18.7 kcal/mol (highlighted in yellow in Figure 2E). This finding suggests that higher-affinity docking locations in the niche that are absent in Cx43 could be responsible for Cx45 GJ g_j_ stimulation, with lower concentrations of resveratrol observed in dual whole-cell patch-clamp experiments (see Figure 1), while lower-affinity docking sites that are similar in both Cxs are responsible for g_j_ inhibition. The most favorable amino acid residues for docking resveratrol in the niche were Ile82 and Thr86 in Cx45(1) TM2 and Val27 in Cx45(2) TM1 (highlighted in Supplementary Table S1; also see Supplementary Movie S2).
3.3. The Effect of Resveratrol on Cx43 and Cx45 Expression and GJ Plaque Size
The obtained results have demonstrated that resveratrol induced fast, reversible, Cx type-dependent effects on g_j_ (Figure 1A–F) and an inhibition of initial g_j_ after preincubation for 24 h (Figure 1G,H). Therefore, we further examined the effect of preincubation with resveratrol on Cx expression and GJ plaque size. Quantitative evaluation of Cx43 and Cx45 proteins was performed after 15 min (time needed to reach the steady-state effect in patch-clamp measurements) and after 24 h of resveratrol treatment. After 15 min from the application of resveratrol at any concentration, total Cx45 and Cx43 protein levels remained unchanged. After 24 h, 1 and 10 μM of resveratrol also had no effect; however, 100 μM increased Cx45 and Cx43 levels by approximately 2.5- and 1.6-fold, respectively (Figure 3A–F). It is well established that the synthesis, assembly, and function of GJs are regulated by phosphorylation of Cxs at specific sites [8]. Therefore, we assessed changes in the phosphorylation of Cx43 at serine residues S279, S282, S368, and S373, which have been shown to be involved in the regulation of GJ expression and function [2]. The obtained results were well aligned with our patch-clamp and immunocytochemical findings. Phosphorylation at S282 and S279 is known to inhibit Cx43 channel conductance, whereas phosphorylation at S373 is associated with increased GJ plaque size [43]. Consistent with this, a decrease in Cx43 GJ g_j_ was accompanied by elevated levels of pS282 and pS279 at both time points. Additionally, an increase in pS373 phosphorylation correlated with the formation of larger Cx43 plaques after 24 h of treatment with 100 μM of resveratrol (see data below). Another well-known phosphorylation site involved in GJIC regulation is S368 [44]. Its phosphorylation affects channel permeability and reduces GJIC between cardiomyocytes due to GJ internalization [45]. We found that pS368 levels decreased after both 15 min and 24 h treatment with 100 μM resveratrol, while a 2-fold increase in pS368 was observed after 24 h of treatment with 10 μM resveratrol (Figure 3B,G,H). In contrast to Cx43, only a few phosphorylation sites of Cx45 are documented [2], and no phosphorylation-dependent changes in Cx45 GJ g_j_ have been demonstrated, so far. Overall, these data show that the complexity of the responses depends on the nature, strength, and duration of the stimulus. In addition, GJ gating and/or Cx trafficking may be regulated through second messenger pathways and the combined actions of multiple kinases and sequential phosphorylation events of Cx43 CT [43].
The relationship between Cx45 phosphorylation and Cx45 GJ properties is basically underexplored due to the absence of specific antibodies. In the current study, we examined the effect of resveratrol on only the total expression of Cx45 (Figure 3A,C,D); however, in general, the impact of protein kinase activity on GJ conductance can be examined using kinase inhibitors. We have previously shown that the potency of certain compounds can be allosterically modulated by phosphorylation of Cx43 CT by Ca^2+^-regulated kinases [8]. The observation of resveratrol-induced changes in phosphorylation of Cx43 CT sites (Figure 3B,G,H) prompted us to verify whether the potency of resveratrol is phosphorylation-dependent. However, staurosporine, a nonselective kinase inhibitor, neither altered the inhibitory effect of resveratrol on Cx43 GJ g_j_ (Figure 4A,B) nor its stimulatory and inhibitory effect on Cx45 GJ g_j_ (Figure 4C,D).
Further, we examined whether the application of resveratrol could induce changes in GJ plaque size. Although changes in Cx expression were observed only after 24 h treatment with 100 μM resveratrol in both HeLa Cx43 and HeLa Cx45 cells (Figure 3A–F), initial g_j_ was altered after application of 1 and 10 μM in HeLa Cx43 (Figure 1G). Exposure of HeLa Cx43 cells to 10 and 100 μM resveratrol for 24 h resulted in the formation of exceptionally large GJ plaques (Figure 5A,C). The estimated Cx43 GJ plaque size was 14.7 ± 1.8 µm^2^ under control conditions and increased to 30.4 ± 5.1 µm^2^ and 56.0 ± 6.5 µm^2^ following treatment with 10 and 100 μM resveratrol, respectively. Surprisingly, initial Cx43 GJ g_j_ was reduced after treatment with 1 and 10 µM resveratrol for 24 h (Figure 1G), while 10 μM increased the GJ plaque size, and 1 μM had no effect on it. We rather expected a reduction in GJ plaque size; however, these results could be in part explained by increased phosphorylation at S279 and S282 (Figure 3), which might be responsible for the reduced Cx43 GJ g_j_ (Figure 1G).
Cx45 GJ plaque size increased after 1 and 10 μM but decreased after treatment with 100 μM resveratrol. Cx45 formed much smaller GJ plaques compared to Cx43. The sizes of Cx45 GJ plaques in control and after treatment with 1, 10, and 100 µM resveratrol were 1.7 ± 0.2, 3.3 ± 0.3, 2.5 ± 0.3, and 0.9 ± 0.1 µm^2^, respectively (Figure 5D). Immunofluorescence analysis revealed that after 24 h treatment with 100 μM resveratrol, Cx45 not only formed smaller GJ plaques, but also was more abundant in the cytoplasm (Figure 5B,D). These results could explain a decrease in electrical coupling, as shown above (Figure 1H).
3.4. Resveratrol Modulates ERK1/2 Activation and Cellular Viability
Phosphorylation of ERK1/2 is a key event in the MAPK signaling pathway, influencing cell survival, proliferation, and death [46,47]. The effect of resveratrol may be either activatory or inhibitory, depending on the disease or cell type [48,49]. It has been shown that the enhancement of Cx43 expression and GJIC by resveratrol involves MAPK and ERK1/2 signaling pathways [29,30]. It is known that resveratrol inhibits ERK1/2 phosphorylation and reduces HeLa cell adhesion [50]. In some other cancer cells, such as mouse melanoma and human renal cell carcinoma, resveratrol targets the ERK1/2 pathway and exerts anti-proliferative, anti-migratory, or pro-apoptotic effects via the modulation of upstream regulators [51,52]. Research on the interplay between GJIC and ERK has mostly focused on Cx43, highlighting that ERK-mediated phosphorylation modulates the gating, assembly, and turnover of Cx43 GJ channels. Phosphorylation of Cx43 CT on S279 and S282 has been reported as ERK1/2-dependent [2,43,53]. As demonstrated in Figure 3, resveratrol affects the phosphorylation of several Cx43 residues, including S279 and S282. Due to the aforementioned observations, we examined the effect of resveratrol on pERK1/2/ERK1/2 and the viability of communication-deficient HeLa WT cells and HeLa cells expressing exogenous Cx43 and Cx45 (Figure 6). We examined changes in protein expression after 15 min and 24 h treatment with resveratrol, but the results of the viability assay are shown only for the 24 h time point, because 15 min exposure did not cause any changes in viability. In HeLa WT and HeLa Cx43 cells, all concentrations of resveratrol increased pERK1/2 levels after 15 min treatment, whereas in HeLa Cx45 cells, only 1 µM concentration increased pERK1/2 levels. Treatment with 1 µM resveratrol for 24 h affected only HeLa Cx45 cells by decreasing pERK1/2 levels. Resveratrol at a 10 µM concentration increased pERK1/2 expression only in HeLa WT cells, decreased it in HeLa Cx45 cells, and had no effect in HeLa Cx43 cells. Treatment with 100 µM resveratrol for 24 h upregulated pERK1/2 activity in all HeLa cell lines, concomitantly suppressing cell proliferation and viability. Resveratrol (100 µM) inhibited proliferation by 48%, 38%, and 51% in HeLa WT, HeLa Cx43, and HeLa Cx45 cells, respectively (Figure 6D–F). HeLa Cx43 and Cx45 cells were more sensitive to 100 µM resveratrol treatment since their viability dropped to 81% and 76%, respectively, while the viability of HeLa WT cells remained near the control level (Figure 6G–I). After treatment with resveratrol, the majority of non-viable HeLa WT, HeLa Cx43, and HeLa Cx45 cells were found in a late apoptotic state.
Taken together, our results show that pERK1/2 activation by resveratrol treatment correlates with a decrease in cell viability and proliferation.
3.5. Impact of Resveratrol on Cytoskeletal Structure and Gap Junction Degradation
Our results indicate that GJ plaque size increased following 24 h treatment with resveratrol at 10 or even at 1 µM concentration in HeLa Cx45 cells (Figure 5), while elevation in Cx43 and Cx45 expression was observed only after the application of 100 µM resveratrol (Figure 3A–F). Interestingly, the elevation of Cx43-pS373, which coincides with Cx43 GJ plaque enlargement [43], was observed by WB and immunofluorescence assays only after 24 h treatment with 100 µM (Figure 3 and Figure 7A). This suggests that other Cx43 phosphorylation sites and/or intermediate pathways could be responsible for this effect.
In the following step, we set out to examine whether resveratrol’s effects on GJ turnover are related to its effect on cytoskeletal integrity. We performed imaging of HeLa Cx43, HeLa Cx45, and HeLa WT live cells labeled with F-actin and α-tubulin fluorescent probes (Supplementary Figure S5). Also, HeLa cells expressing Cx43 tagged with EGFP (Cx43-EGFP) were used for time-lapse imaging of GJ plaque dynamics (Figure 7B). Resveratrol treatment did not cause an obvious disarrangement of actin and microtubule cytoskeleton network, and consequently, regular internalization of GJ plaques and formation of annular junctions were supposed to persist.
Next, we carried out time-lapse imaging to monitor the internalization of Cx43 GJ plaques. Some studies suggest that stress may slow down the internalization of Cx43 plaques [54]. It has been reported that the presence of GJ at the plasma membrane persists for up to 5 h [55]. Since it is virtually impossible to monitor the full life-time of GJ plaque starting from the exact zero-time of its formation until complete AGJ formation, we relied on statistics and performed time-lapse imaging of randomly selected cell pairs with GJ plaques and measured the number of internalized GJ plaques during 210 min recording under control conditions and after treatment with 10 µM resveratrol for 24 h (Figure 8A). The duration of the time-lapse experiments was selected because, under control conditions, Cx43 GJ plaques typically disassembled within a 191 ± 20 min (n = 30) period. Our findings indicate that resveratrol stabilized Cx43 plaques, leading to a 3-fold decrease in the internalization rate (Figure 8B). For these experiments, we chose a lower resveratrol concentration (10 µM) since it did not affect cell viability and proliferation (Figure 6D–I).
Furthermore, we examined the possible mechanism underlying retarded Cx43 degradation. Cx43 is regularly internalized from the plasma membrane and directed toward degradation, mainly through the endosome–lysosomal pathway. During this process, GJs are internalized, and AGJs are formed and transported for endosomal degradation [56]. We estimated the number of AGJs fused with lysosomes (Figure 8C) in control and in resveratrol-treated HeLa Cx43-EGFP cells and did not detect any changes in Cx43 and lysosome co-localization (Figure 8D).
Resveratrol at concentrations of 10 and 100 µM increased the Cx43 GJ plaque size after 24 h (Figure 5A,C) with a significant increase in total Cx43 expression only at a concentration of 100 µM (Figure 3B,D). In contrast, the Cx45 GJ plaque size was already significantly increased after treatment with 1 µM resveratrol; however, in contrast to Cx43, 100 µM resveratrol strongly reduced the Cx45 GJ size, although total Cx45 expression increased by 2.5-fold, like in the case of Cx43. Importantly, a decrease in the Cx45 GJ plaque size and g_j_ (Figure 1H) was followed by increased Cx45 degradation, as assessed by Cx45 and lysosome co-localization, which increased by ~2-fold (Figure 8E,F). Pearson’s correlation coefficient was used to quantify the degree of Cx45 and lysosome co-localization due to the fact that Cx45 forms very small GJ plaques (Figure 5) and inestimable AGJs compared to Cx43. However, the reasons why total Cx45 expression increases despite enhanced Cx45 degradation remain incompletely understood. We can only speculate that under stress conditions, when Cx45 GJ plaque internalization and degradation are accelerated, some compensatory mechanisms are activated to stimulate Cx45 synthesis, but not new GJ plaque formation, which is a much more complex process involving connexin oligomerization, hemichannel trafficking to the membrane, and docking of apposed hemichannels.
4. Discussion
In the present study, we found that resveratrol induced time- and dose-dependent changes in Cx43 and Cx45 expression, phosphorylation, GJ plaque size and conduction, and HeLa cell viability. Comparative analysis of Cx43 and Cx45 revealed that resveratrol suppressed GJIC in Cx43-expressing cells but enhanced it in Cx45-expressing cells (Figure 1). The dual effect could be at least in part explained by molecular docking experiments, which showed that Cx45 exhibited two different docking profiles of higher and lower affinity, while only the latter was observed in Cx43 (Figure 2). GJIC modulation is often linked with changes in connexin expression levels and GJ plaque size [57]. However, usually only a small part of the channels composing GJ plaques are functional, as has been shown for Cx43 [58], Cx45 [59], Cx57 [60], and Cx36 [61]. Therefore, the increase in GJ plaque size may not guarantee an increase in its total conductance. Long-term (24 h) treatment with resveratrol caused a reduction in initial g_j_ (Figure 1), but unexpectedly, this effect was followed by an increase in GJ plaque size in HeLa Cx43 cells (Figure 5), suggesting that the functional efficiency of GJ plaques was decreased. In HeLa Cx45 cells, the initial g_j_ remained unchanged (Figure 1), while GJ plaque size showed a tendency to increase (Figure 5). Changes in GJ channel conductance, along with the modulation of connexin expression, GJ plaque size, and turnover, are a complex process with phosphorylation being one of the most important regulators. For example, phosphorylation at S279/282 is linked to reduced GJIC, pS373 increases the amount of Cx43 at the plasma membrane and enhances the size of GJ plaques, while phosphorylation at S368 is involved in the dynamic turnover of GJ, influencing their assembly, internalization, and degradation [43]. Our findings mostly correspond to the previously published data, as the reduction in Cx43 GJ conductance (Figure 1) was accompanied by increased levels of Cx43 pS282 and pS279 (Figure 3), while phosphorylation at S373 (Figure 3) likely contributed to the formation of larger Cx43 plaques (Figure 5 and Figure 7).
GJIC efficiency may depend on Cx43 phosphorylation that affects the trafficking of hemichannels, their docking to form GJ channels, GJ aggregation into GJ plaques, and plaque turnover [43,62]. Most connexins are phosphorylated at specific sites by multiple kinases, and interplay between kinases may determine the ultimate cellular responses [8,63]. It was demonstrated that resveratrol modulates Cx43 phosphorylation via MAPK and phosphoinositide 3 kinase (PI3K)/protein kinase B (Akt) activation [64,65,66]. ERK1/2 phosphorylation is a central step in the MAPK pathway, regulating cell survival, proliferation, and death. Resveratrol modulates this pathway in a context-dependent manner, either activating or inhibiting it depending on the disease or cell type [52,67]. We found that ERK1/2 activation is associated with cell viability, as after treatment with the highest used resveratrol concentration (24 h, 100 µM), pERK1/2 levels peaked, while viability decreased (Figure 6). Resveratrol caused greater reduction in the viability of Cx43- and Cx45-expressing HeLa cells compared with HeLa WT cells (Figure 6). These results resemble the findings of other studies, where experiments with HeLa cells lacking exogenous Cxs and expressing Cx37, Cx40, and Cx43 showed that the pro-apoptotic effect of GJIC varied based on a connexin type, and HeLa Cx43 cells were most sensitive to streptonigrin-induced apoptosis. Moreover, the authors suggested that apoptosis was promoted by Cx43 and Cx40, but not by Cx37, via GJ transfer of IP_3_-dependent pro-apoptotic signals [68]. Our important finding was that treatment with 1 µM resveratrol increased the proliferation of HeLa Cx43 cells (Figure 6). This observation suggests that resveratrol may exert a concentration-dependent biphasic effect, i.e., lower concentrations could promote cell proliferation, while higher concentrations might inhibit it. Many polyphenolic extracts and pure polyphenols, such as quercetin and curcumin, tend to induce apoptosis and reduce cancer cell viability, in a dose-dependent manner, while having little or no cytotoxic effect on normal cells [69,70,71]. Resveratrol displays a dual action as well, being cytoprotective in healthy cells and cytotoxic in cancer cells [72,73]. Interestingly, a proliferation-promoting effect was demonstrated in T47D human breast cancer cells, explained by phytoestrogenic properties of resveratrol [74]. Usually, resveratrol acts as a tumor suppressor, but in certain estrogen-sensitive human breast cancer cells depending on ER isoforms or response elements, it can function as an estrogen agonist, thus promoting cell growth.
In addition to the above described mechanisms, resveratrol has been shown to disrupt the tubulin network by inhibiting tubulin polymerization [75,76,77]. Also, resveratrol has been shown to affect the actin cytoskeleton in a concentration-dependent manner, disrupting it in MDA-MB-231 breast cancer cells [78], or promoting actin accumulation at injury sites in mouse C2C12 myoblasts [79]. The microtubule and actin cytoskeleton is important for hemichannel transport to the plasma membrane, for maintaining their proper localization, and for stabilizing GJ plaques [80]. Our data show that resveratrol did not disrupt the actin cytoskeleton or the tubulin network in HeLa cells (Figure 7), independent of the presence or type of connexin. Although we cannot exclude the possibility that other factors involved in Cx trafficking, such as the scaffold protein ZO-1 [81], motor protein like dynein [82], GTPase dynamin 2 [83], E-cadherin, or N-cadherin [84], could be affected by resveratrol.
The time-lapse imaging showed that resveratrol stabilized Cx43 plaques, leading to a three-fold decrease in the internalization rate compared to the control. This finding is in agreement with previous work showing that genotoxic stress reduces Cx43 plaque internalization in primary bovine corneal endothelium cells that have been transiently transfected with Cx43-GFP [54]. Plaque size and lifespan are closely associated with the lysosomal degradation pathway, and its disruption may affect GJ plaque stability [56]. We hypothesized that resveratrol could trigger lysosomal degradation; however, no obvious changes were observed. Moreover, we expected that an increase in GJ plaque size would result in enhanced GJIC; however, Cx43 GJ g_j_ was decreased after 24 h treatment with 10 µM resveratrol (Figure 1). Presumably, this phenomenon could be explained by our earlier observations that a large total number of channels in the GJ plaques is unrelated to the functional efficiency of the plaques, because only a small number of channels are functional, and the overload of GJ plaques with channels may even hamper their functionality.
Resveratrol was selected for this study due to its broad range of physiological effects, including cardioprotective, anti-inflammatory, and metabolic actions mediated through multiple signaling pathways. Although resveratrol exhibits significant biological activity in vitro at relatively high concentrations (≥100 µM), several studies, including our own, have reported biological effects at lower concentrations [85,86,87]. Notably, in our study, the effects of resveratrol were observed even at concentrations of 1–10 µM. These concentrations, however, are still considerably higher than the plasma levels typically achieved after oral intake of resveratrol supplements or resveratrol-containing products such as red wine, which typically remain below 0.02 µM [88,89]. These pharmacokinetic limitations reflect the rapid metabolism (a half-life of approximately 9 h) and clearance of resveratrol, resulting in a short-lived presence of biologically active concentrations in the circulation. According to previous reports, extensive metabolism of resveratrol in the intestine and hepatic metabolism leads to oral bioavailability of the parent compound of only about 1% [90]. However, due to its lipophilic nature, concentrations of resveratrol in tissues may exceed those observed in plasma [91]. This phenomenon has also been demonstrated in patients with colorectal cancer, supporting the colorectum as a potential target for chemoprevention by oral resveratrol intake [92]. Interestingly, some studies showed that the bioavailability of resveratrol can be enhanced by using more potent resveratrol analogs, such as SRT501 [93], or by employing advanced delivery systems, including liposomal encapsulation [94], nanoparticles, and nanoemulsions. These approaches improve solubility, stability, and systemic exposure, thereby increasing the therapeutic potential [95,96].
Considering the increasing focus on antioxidant polyphenols, both as dietary components and valuable food industry by-products, our results hold notable relevance. Proper intercellular communication is fundamental for maintaining tissue integrity and preventing pathological processes, and therefore, our findings contribute to a better understanding of how resveratrol may support cellular functions through modulation of GJIC. However, it is important to note that the dose of resveratrol, its bioavailability, and long-term effects in humans remain under investigation, making it a promising, but not guaranteed, “miracle cure”.
5. Conclusions
In HeLa cells expressing exogenous Cx43 or Cx45, resveratrol induced time-, dose-, and connexin-dependent changes in GJ properties and cell viability. Resveratrol inhibited Cx43 GJ g_j_, but exerted a biphasic effect on Cx45 GJ g_j_, presumably due to differences in docking profiles, with a high-affinity docking site responsible for g_j_ stimulation absent in Cx43.
Resveratrol increased the connexin expression levels and enlarged GJ plaques in both Cx43- and Cx45-expressing cells.
Resveratrol provoked Cx43 phosphorylation, which was accompanied by the activation of the ERK1/2 signaling pathway, which, in turn, correlated with a reduction in HeLa cell viability and proliferation. Notably, the viability of HeLa cells expressing either of the connexins was more sensitive to resveratrol treatment compared with HeLa WT cells.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Rackauskas M. Neverauskas V. Skeberdis V.A. Diversity and properties of connexin gap junction channels Medicina 201046110.3390/medicina 4601000120234156 · doi ↗ · pubmed ↗
- 2Aasen T. Johnstone S. Vidal-Brime L. Lynn K.S. Koval M. Connexins: Synthesis, Post-Translational Modifications, and Trafficking in Health and Disease Int. J. Mol. Sci.201819129610.3390/ijms 1905129629701678 PMC 5983588 · doi ↗ · pubmed ↗
- 3Söhl G. Willecke K. Gap junctions and the connexin protein family Cardiovasc. Res.20046222823210.1016/j.cardiores.2003.11.01315094343 · doi ↗ · pubmed ↗
- 4Mc Nutt N.S. Weinstein R.S. The Ultrastructure of the Nexus: A Correlated Thin-Section and Freeze-Cleave Study J. Cell Biol.19704766668810.1083/jcb.47.3.6665531667 PMC 2108148 · doi ↗ · pubmed ↗
- 5Palacios-Prado N. Hoge G. Marandykina A. Rimkute L. Chapuis S. Paulauskas N. Skeberdis V.A. O’Brien J. Pereda A.E. Bennett M.V.L. Intracellular magnesium-dependent modulation of gap junction channels formed by neuronal connexin 36J. Neurosci.2013334741475310.1523/JNEUROSCI.2825-12.201323486946 PMC 3635812 · doi ↗ · pubmed ↗
- 6Bukauskas F.F. Verselis V.K. Gap junction channel gating Biochim. Biophys. Acta 20041662426010.1016/j.bbamem.2004.01.00815033578 PMC 2813678 · doi ↗ · pubmed ↗
- 7Dbouk H.A. Mroue R.M. El-Sabban M.E. Talhouk R.S. Connexins: A myriad of functions extending beyond assembly of gap junction channels Cell Commun. Signal.20097410.1186/1478-811X-7-419284610 PMC 2660342 · doi ↗ · pubmed ↗
- 8Mickus R. Raškevičius V. SarapinienėI. Mikalayeva V. Prekeris R. Skeberdis V.A. Phosphorylation-dependent allosteric regulation of Cx 43 gap junction inhibitor potency Biomed. Pharmacother.202417411655010.1016/j.biopha.2024.11655038593702 · doi ↗ · pubmed ↗