Transcriptome and Hormone Analysis Revealed Jasmonic Acid-Mediated Immune Responses of Potato (Solanum tuberosum) to Potato Spindle Tuber Viroid Infection
Iva Marković, Bernard Jarić, Jana Oklešťková, Jitka Široká, Kristina Majsec, Jasna Milanović, Snježana Kereša, Ivanka Habuš Jerčić, Ondřej Novák, Snježana Mihaljević

TL;DR
This study explores how jasmonic acid influences potato plants' immune responses to a viroid infection, revealing its role in symptom development and defense mechanisms.
Contribution
The study identifies a modulatory role of jasmonic acid (JA) in potato's immune response to PSTVd infection through transcriptomic and hormone analyses.
Findings
JA-deficient opr3 plants showed enhanced JA signaling and increased abscisic acid and auxin content after PSTVd infection.
JA-insensitive coi1 plants exhibited reduced hormone levels and lower expression of defense-related genes, leading to milder symptoms but increased viroid accumulation.
Methyl jasmonate treatment alleviated symptoms in infected wild-type potato plants.
Abstract
Potato is a globally important non-cereal crop in which infection with potato spindle tuber viroid (PSTVd) can cause stunted growth and significantly reduce tuber yield. We previously showed that PSTVd induces accumulation of the plant hormone jasmonic acid (JA) and alters antioxidant responses in potato plants. To clarify the role of JA in response to PSTVd, we analyzed disease development in transgenic JA-deficient opr3 and JA-insensitive coi1 lines compared to the wild-type. Transcriptomic analysis using RNA-Seq revealed that most genotype-specific differentially expressed genes (DEGs) in all comparisons were enriched in plant hormone signal transduction, plant-pathogen interaction, and MAPK signaling pathways, although the number of DEGs varied. These differences were confirmed by independent data from RT-qPCR, hormone, and hydrogen peroxide (H2O2) analyses. After PSTVd infection,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
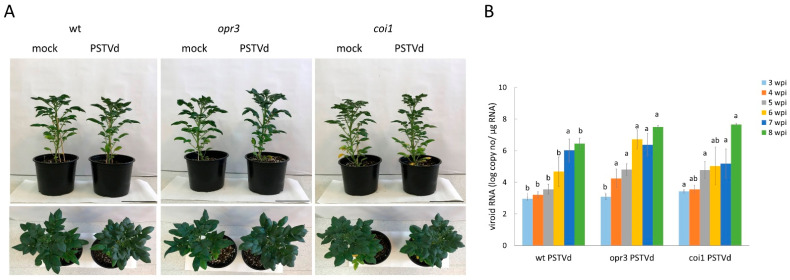 Figure 1
Figure 1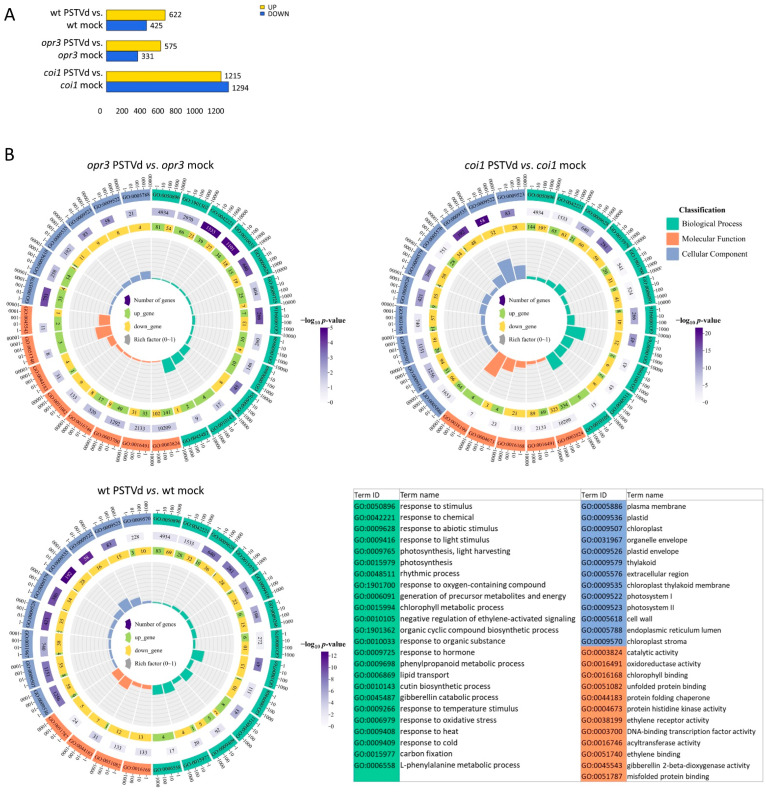 Figure 2
Figure 2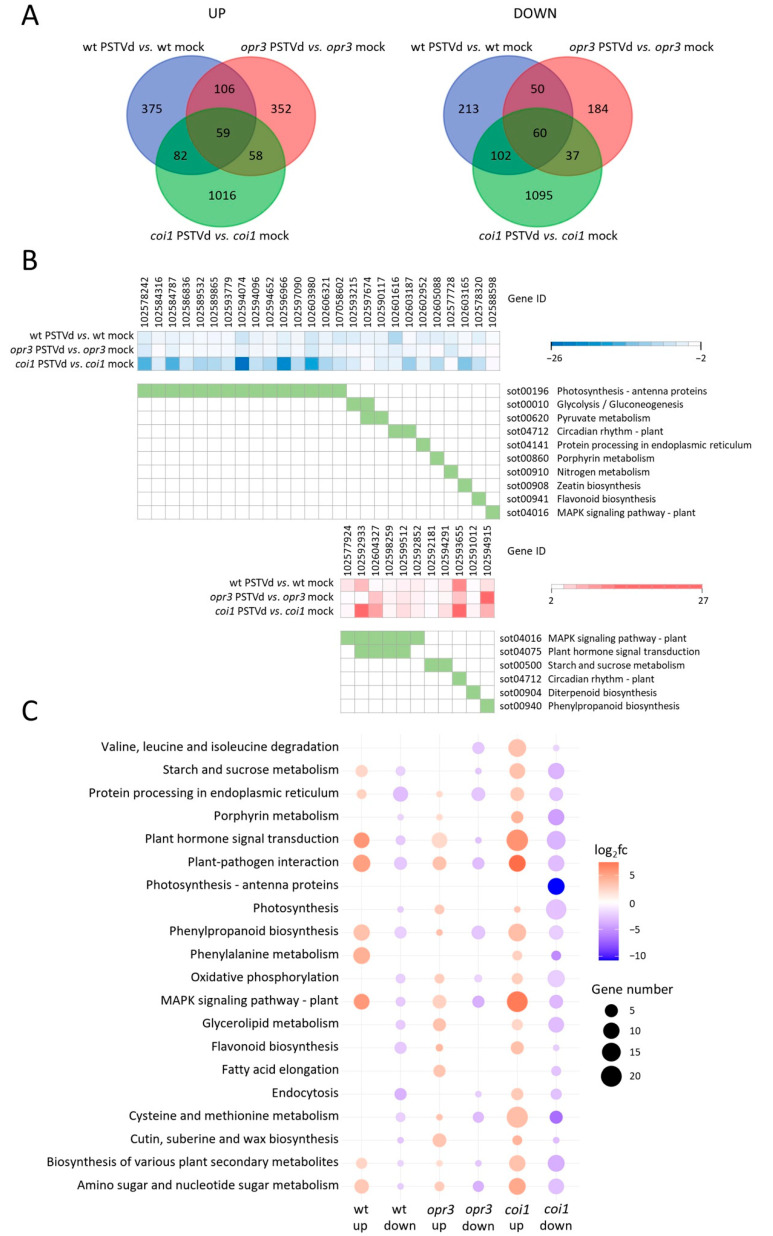 Figure 3
Figure 3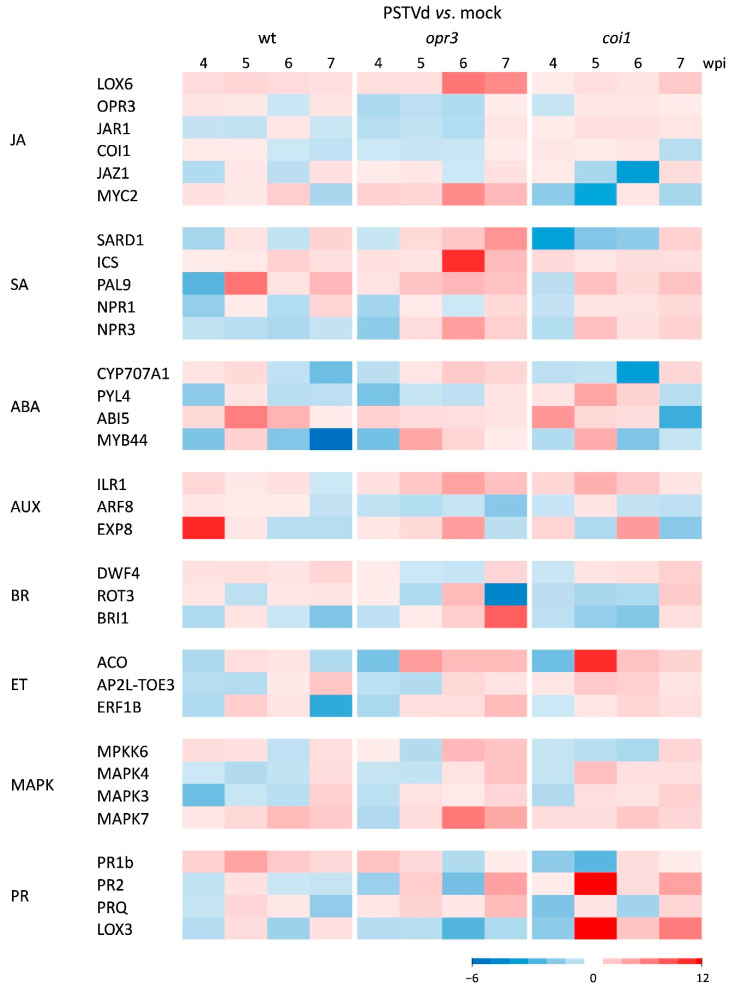 Figure 4
Figure 4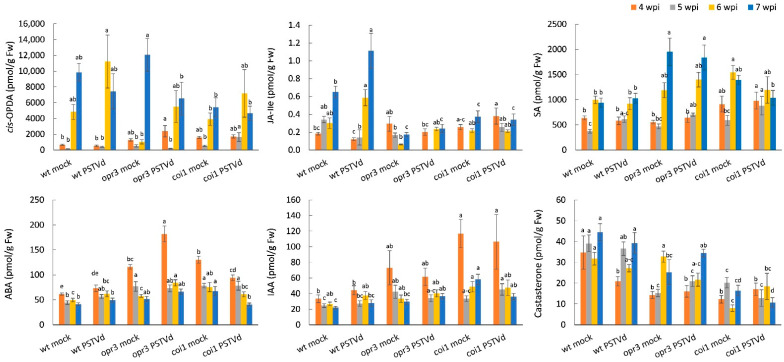 Figure 5
Figure 5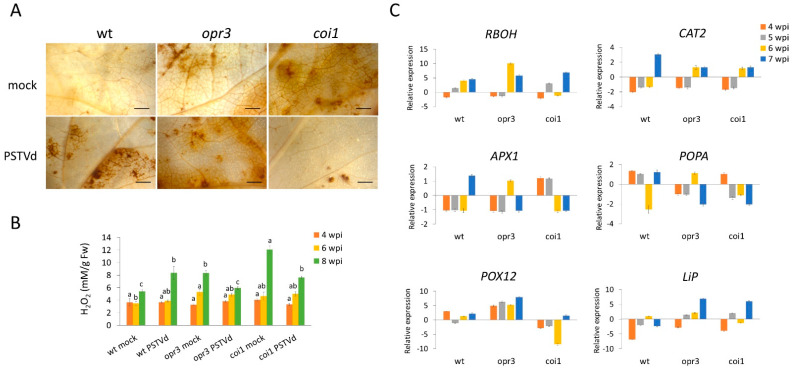 Figure 6
Figure 6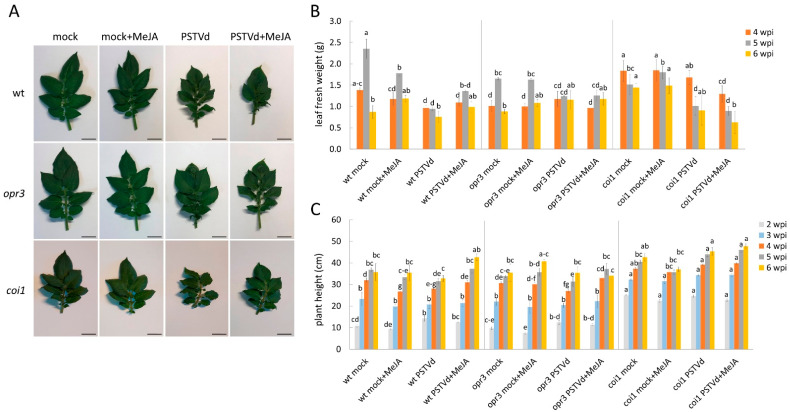 Figure 7
Figure 7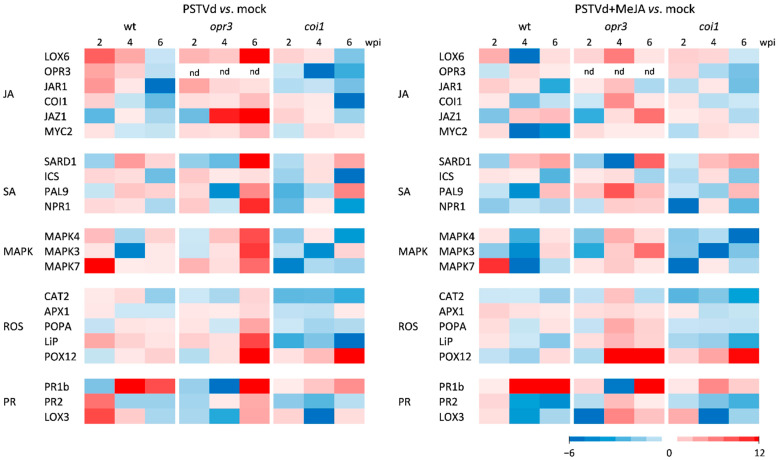 Figure 8
Figure 8- —Croatian Science Foundation
- —Czech Science Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect-Plant Interactions and Control · Plant Virus Research Studies · Plant-Microbe Interactions and Immunity
1. Introduction
Potato (Solanum tuberosum) is the fourth most important food crop worldwide, after rice, wheat, and maize [1]. Potato spindle tuber viroid (PSTVd) is a subviral plant pathogen that can cause significant losses in yield and marketable quality of potato tubers. This threat is heightened by the fact that no potato varieties are known to be naturally resistant to this pathogen, and no curative treatment exists for this disease [2].
PSTVd is a small, non-coding, highly structured, circular, single-stranded RNA molecule consisting of approximately 341–364 nucleotides [3]. It is not clear how plants recognize this non-coding biotrophic pathogen, but transcriptomic studies on some viroid-host interactions suggest that viroids can activate pattern-triggered immunity (PTI) and effector-triggered immunity (ETI), as well as general downstream defense responses, including the generation of reactive oxygen species (ROS), mitogen-activated protein kinase (MAPK) cascades, and hormone signaling, which ultimately lead to transcriptional reprogramming of cells [3,4].
The plant hormone jasmonic acid (JA) is generally considered to play a defensive role in plant responses to necrotrophic pathogens, while salicylic acid (SA) is involved in responses to biotrophic pathogens [5,6]. However, increasing evidence supports a positive role for JA and its signaling pathway in defense against certain biotrophic fungal, bacterial, and viral pathogens in various plant species [7,8,9,10,11,12]. The accumulation of JA during pathogen perception promotes the expression of CORONATINE INSENSITIVE1 (COI1), which binds jasmonate ZIM domain proteins (JAZ) and promotes their degradation. This enables positive-acting transcription factors such as MYC2 to activate the transcription of JA-responsive genes involved in a wide range of defense responses, including maintenance of redox homeostasis, modulation of signaling crosstalk, and stimulation of defense compound production [13,14,15]. In response to pathogens, JA does not act alone but interacts with other hormones and signaling pathways through the actions of JAZ and MYC2 regulatory proteins [14,15].
Transcriptomic data from plant hosts infected with viroids indicate that the expression of genes involved in JA biosynthesis and signaling pathways is affected, depending on the specific viroid-host interaction [16,17,18,19,20,21,22,23]. However, studies examining whether changes in gene expression coincide with altered accumulation of endogenous JA levels in affected host plants are rare [24,25]. The accumulation of JA in potato leaves during PSTVd infection is also significant because there is little or no accumulation of SA [25], another important defense hormone that, along with JA, determines the outcome of defense responses. To elucidate the molecular mechanisms underlying basal defense responses in potato, this study analyzed the role of JA accumulation in response to PSTVd. We conducted a comparative transcriptomic analysis during PSTVd infection in transgenic lines with downregulated JA biosynthesis or JA signaling and in the corresponding wild-type potato using RNA-Seq. Transcriptome analysis showed that PSTVd infection suppresses the expression of genes related to photosynthesis and primary metabolism while activating genes involved in protein metabolism, signal transduction pathways, and defense responses. In this study, we focused on hormone, MAPK, and ROS signaling pathways. We also analyzed the temporal dynamics of gene expression, hormone levels, and H_2_O_2_ accumulation to determine how the timing of their expression influences plant susceptibility to PSTVd infection. Furthermore, we tested the effect of exogenous methyl jasmonate (MeJA) on gene expression and the perception of viroid infection. Key genes and pathways involved in basal defense against PSTVd in potato were identified. Results show that JA and its signaling modulate many defense responses to PSTVd in potato, and that their absence increases viroid accumulation but reduces symptom development, revealing new aspects of plant-viroid interactions.
2. Materials and Methods
2.1. Plant Material, Viroid Inoculation, and Sampling
Potato cv. Désirée wild-type, two transgenic StOPR3-RNAi silenced lines (opr3A3 and opr3Z2) resulting in JA deficiency, and two transgenic StCOI1-RNAi silenced lines (coi1H1 and coi1X5) resulting in JA insensitivity [26], were propagated in stem node tissue culture. The in vitro plantlets were transferred to a soil/perlite mixture (3:1) and grown in a phytochamber at 22 °C, 60% relative humidity, and a 16-h photoperiod with a light intensity of 160 μmol m^−2^ s^−1^. Healthy potato plants with 3–4 leaves were inoculated with H_2_O (mock) or a viroid inoculum (5.7 × 10^7^ viroid copies/μL) prepared from plants infected with PSTVd (GenBank: KF418768) as described [25]. Briefly, the two lower leaves were dusted with Celite 545 (Sigma-Aldrich, St. Louis, MO, USA) and rubbed with 2 µL of inoculum per leaf. After 10 min, the leaves were thoroughly washed with H_2_O. After inoculation, plants were grown at 26 °C to enhance viroid replication and symptom development [16]. The first three intact leaves above the inoculated leaves were harvested weekly from 1 to 8 weeks post-inoculation (wpi), immediately frozen in liquid nitrogen, and stored at −80 °C for RNA, hormone, and H_2_O_2_ quantification. Leaves from at least four mock- and PSTVd-inoculated plants per genotype per time point were harvested. Each biological replicate consisted of leaves from a single plant. The experiment was repeated twice.
For exogenous MeJA treatment, plants were sprayed twice with H_2_O or 1 mM MeJA (Sigma-Aldrich, St. Louis, MO, USA): one day before and six days after viroid inoculation (i.e., 24 h before the first sampling). Leaf samples from at least four mock- and PSTVd-inoculated plants per genotype per treatment per time point were harvested weekly from 1 to 6 wpi.
To distinguish effects of viroid inoculation from possible developmentally regulated changes, healthy, mock-inoculated plants served as controls in all experiments. The appearance of symptoms, such as epinasty and pointed upper leaves, was assessed visually, while growth retardation was evaluated by measuring plant height and fresh weight of the apical leaves (Figure S1).
2.2. RNA Extraction and RT-qPCR
Total RNA was extracted using the Direct-zol RNA Mini Prep Kit (Zymo Research, Irvine, CA, USA) according to the manufacturer’s instructions. Absolute quantification of PSTVd RNA was performed using the Brilliant III Ultra-Fast qRT-PCR Master Mix (Agilent, Santa Clara, CA, USA) in a duplex reaction with specific primers and probes for PSTVd and cytochrome oxidase 1 (COX) gene as described [27,28].
For plant gene expression analysis, total RNA was reverse transcribed using the SuperScript IV First-Strand Synthesis System (Thermo Fisher Scientific, Waltham, MA, USA). qPCR was performed with the SsoAdvanced Universal SYBR Green Supermix (Bio-Rad, Hercules, CA, USA) in a CFX96 Touch Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). The PCR cycling conditions were 95 °C for 30 s, followed by 40 cycles of 10 s at 95 °C and 20 s at 60 °C, then a dissociation curve was generated to verify amplification specificity. Data were normalized using an inter-plate calibrator to correct for run-to-run variability. Relative gene expression was calculated using the 2^−ΔΔCT^ method [29], with elongation factor 1 (EFα1) as the reference gene [30]. Primers were designed using Primer3Plus version 3.3.0 or as described in [31,32,33,34,35] (Table S1). Primer efficiency and melt curves were checked and found acceptable for RT-qPCR.
2.3. RNA-Seq Gene Expression Analysis
Total RNA from three independent biological replicates (each an independent plant) per genotype and treatment was used to construct cDNA libraries with the TruSeq Stranded mRNA Library Prep Kit (Illumina, San Diego, CA, USA). Sequencing was performed by Macrogen (Seoul, South Korea) using paired-end sequencing on an Illumina NovaSeq 6000 platform. Quality control, adapter trimming, read filtering, mapping, and analysis were performed by Macrogen through their internal pipeline. Reads with mean Phred values below 15 and lengths shorter than 36 bp were removed. Quality statistics are shown in Table S2. Paired-end RNA-Seq reads were mapped to the reference genome SolTub_3.0 [36] with HISAT2 (v2.1.0) using the Bowtie2 (v2.3.4.1) aligner [37]. Mapped reads were assembled with StringTie to generate read counts, which were further analyzed in R with DESeq2 [38]. In DESeq2, size factors were estimated and used to normalize the libraries with the Relative Log Expression method. The negative binomial Wald test was applied to the normalized count data to determine p-values for differentially expressed genes (DEGs) between each comparison pair. Macrogen reported expression data with FDR-adjusted p-values for all comparisons. The criteria for DEGs were |log_2 _ fold change| ≥ 2 and p-value < 0.05. Gene ontology (GO) enrichment analysis was performed using g:Profiler (https://biit.cs.ut.ee/gprofiler/gost (accessed on 12 August 2022)), and data were visualized using ChiPlot (https://www.chiplot.online/polarPlot/circle_enrich_plot.html (accessed on 11 January 2025)). MapMan analysis was performed as described [39]. Venn diagrams and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis were generated using R packages (v4.2.2), including VennDiagram and KEGGREST. Transcription factor (TF) analysis was performed using PlantTFDB v5.0 [40].
2.4. Plant Hormone Analysis
The acidic plant hormones (SA; jasmonoyl-L-isoleucine, JA-Ile; cis-(+)-12-oxo-phytodienoic acid, cis-OPDA; abscisic acid, ABA; indole-3-acetic acid, IAA) were determined as described [41]. Briefly, samples (10 mg fresh weight) were extracted in cold 10% aqueous methanol with the addition of internal standards (IS) labeled with stable isotopes (20 pmol SA-d4, Sigma Aldrich, St. Louis, MO, USA; 5 pmol (−)-JA-d2-Ile, 10 pmol OPDA-d5 and 10 pmol IAA-^13^C_6_ OlChemIm Ltd., Olomouc, Czech Republic; 10 pmol ABA-d6, National Research Council Canada, Saskatoon, SK, Canada). The extracts were purified on Oasis^®^ HLB solid phase extraction columns (1 cc/30 mg, Waters) as described [42]. Analysis was performed on an Agilent 6490 Triple Quadrupole LC/MS system coupled to a 1290 Infinity LC system (Agilent Technologies, Santa Clara, CA, USA). The amount of SA-d_4_ is set higher than that of other IS due to chromatographic issues related to peak shape (peak broadening associated with the history of the analytical column) and detection sensitivity. A level of 20 pmol ensures well-detectable chromatographic peaks of SA-d_4_ in most matrices analyzed. Additionally, the higher SA-d_4_ peak areas more closely match those of endogenous SA, which is important for accurate quantification. In contrast, the amount of (−)-JA-d_2_-Ile is set lower than that of other IS, as 5 pmol is sufficient to obtain well-detectable chromatographic peaks in all tested matrices. Furthermore, for quantification, the lower JA-d_2_-Ile peak area is closer to the peak areas of endogenous JA-Ile. The methodology was validated for accuracy and precision as described in [41].
Extraction and quantification of inactive and active endogenous BR were performed as described previously [43,44]. Samples (10 mg fresh weight) were extracted in ice-cold 60% acetonitrile, and 25 pmol deuterium-labeled internal standards of BRs (OlChemIm Ltd., Olomouc, Czech Republic) were added to each sample. After 12 h, samples were centrifuged (36,670 g, 15 min, 4 °C) and supernatants were purified using 50 mg Discovery DPA-6S cartridges (Supelco, Bellefonte, PA, USA). After evaporation to dryness, samples were reconstituted in 40 µL methanol and analyzed by liquid chromatography with tandem mass spectrometry (UHPLC-MS/MS) using an ACQUITY UPLC I-Class System (Waters, Milford, MA, USA) with a triple quadrupole mass spectrometer Xevo TQ-S MS (Waters MS Technologies, Manchester, UK).
2.5. Hydrogen Peroxide Analysis
Localization of H_2_O_2_ in leaf tissue was performed using 3,3-diaminobenzidine (DAB, Sigma-Aldrich, St. Louis, MO, USA) as described [45] and observed under a stereomicroscope (Zeiss SteREO Discovery.V20, Jena, Germany). The H_2_O_2_ content in leaves was determined spectrophotometrically using the TiOSO_4_ method [46]. Absorbance was measured at 405 nm.
2.6. Statistical Analysis
The number of biological replicates in each experiment is indicated in the figure captions. When comparing two mean values, a Student’s t-test was performed after checking the homogeneity of variance between the two samples with the F-test. Differences between the means of more than two groups were statistically assessed using two-way analysis of variance (ANOVA) followed by a post hoc Duncan’s multiple range test (DMRT).
3. Results
3.1. Differences in Transcriptomic Responses Between JA-Deficient, JA-Insensitive, and Wild-Type Plants Infected with PSTVd
To identify genes and pathways specifically regulated by JA in response to PSTVd infection, transcriptomic profiles of PSTVd-infected leaves were compared with control leaves in wild-type, JA-deficient opr3, and JA-insensitive coi1 lines. Samples were collected at 5 wpi, when symptoms began to appear (Figure 1A and Figure S1), and viroid load was significantly higher (p < 0.05) in opr3 and coi1 plants (6.3 × 10^5^ and 5.8 × 10^5^ copies/μg RNA, respectively), compared to wild-type plants (3.6 × 10^4^ copies/μg RNA) compared to wild-type plants (Figure 1B).
Overall, twice as many DEGs were detected after PSTVd infection in coi1 than in opr3 or wild-type (2509, 906, and 1047 DEGs, respectively) (Figure 2). GO enrichment analysis revealed that the Biological Process (BP) GO terms related to responses to chemical and abiotic stimuli, and photosynthesis were significantly enriched in all examined lines after PSTVd infection (Figure 2, Table S3). However, the BP GO terms response to oxygen-containing compounds and negative regulation of ethylene signaling were enriched only in coi1, while response to hormone, phenylpropanoid metabolism, and lipid transport were enriched only in opr3. In the Molecular Function (MF) category, GO terms related to catalytic activity, oxidoreductase activity, and ethylene binding were enriched in coi1 and opr3; chlorophyll binding was enriched in coi1 and wild-type; while unfolded protein binding was enriched in opr3 and wild-type. Among the Cellular Component (CC) GO terms, those linked to the chloroplast and thylakoid membrane were significantly enriched in all examined lines in response to PSTVd infection.
DEGs from the comparison between PSTVd- and mock-inoculated plants of all tested lines were assigned to MapMan functional classes. In the Biotic Stress overview, the most represented DEGs are related to signaling, proteolysis, and cell wall modification, followed by genes involved in transcriptional regulation, hormone signaling, and redox state (Figure S2, Table S4). Among DEGs encoding transcription factors (TFs), those for ethylene responsive factors (ERFs) and WRKYs were the most abundant in infected plants of all tested lines, as confirmed by PlantTFDB analysis (Figure S3). Additionally, activation of genes from the MYB and bHLH TF families was observed in wild-type and opr3, respectively. In coi1, a significant number of DEGs for ERF, MYB, and TCP TFs were downregulated compared to the wild-type after PSTVd infection. Besides stress responses and developmental processes, ERFs also participate in ET signaling. Regarding other hormone signaling pathways, more AUX-related genes were upregulated in opr3 and coi1 compared to the wild-type after PSTVd infection (Figure S2, Table S4).
3.2. Comparative Analysis of DEGs
The overlap between DEGs in PSTVd-infected and control samples of opr3, coi1, and wild-type plants is shown in Venn diagrams (Figure 3A). Functional KEGG analysis identified only 11 induced genes and 28 repressed genes common to both wild-type and transgenic lines, suggesting regulation by the same or overlapping pathways (Figure 3B, Table S5). Common upregulated genes were enriched in MAPK signaling and plant hormone signaling, including MAPK7 and ET signaling genes (green ripe-like 1 GRL1, ethylene receptors ETR2-like and EBF1-like, and ERF1B-like). Common downregulated genes were enriched in photosynthesis-antenna proteins, glycolysis/gluconeogenesis, and pyruvate metabolism, indicating repression of primary metabolism after PSTVd infection.
Most genotype-specific DEGs detected after PSTVd infection in all comparisons were enriched in plant hormone signal transduction, plant-pathogen interaction, and MAPK signaling pathways, although the number of DEGs varied (Figure 3C). The coi1-specific DEGs related to hormone signaling were mainly involved in ET signaling (ERF1B-like), AUX signaling (Aux/IAAs, SAURs, and AUX-amido synthetase GH3.6 and GH3.1 genes), ABA signaling (ABA receptor PYL4-like, protein phosphatase 2C (PP2C)), GA signaling (GID1B-like), and SA signaling (regulatory protein NPR3-like and TGA2) (Table S5). The opr3-specific upregulated DEGs related to hormone signaling were mainly involved in AUX signaling (GH3.5, auxin transporter-like protein LAX3, and AUX/IAA14-like genes), JA signaling (MYC4-like), ET signaling (ETHYLENE INSENSITIVE 3-like 3 protein EIL3 gene), and MAPK signaling (serine/threonine-protein kinase SAPK3-like), while genes related to CK biosynthesis and calcium signaling, were downregulated (Table S5). In addition, several more pathways were enriched with coi1-specific upregulated DEGs, such as cysteine and methionine metabolism (including ET biosynthesis genes ACO and ACS) and phenylpropanoid biosynthesis, while photosynthesis and oxidative phosphorylation pathways were enriched with downregulated coi1-specific DEGs. In contrast, only infected opr3 plants showed upregulated DEGs assigned to glycerolipid metabolism, and cutin, suberin, and wax biosynthesis, while opr3-specific downregulated DEGs were assigned to protein processing and phenylpropanoid biosynthesis, including class III peroxidases (Figure 3C).
To gain better insight into the opposing regulation of defense responses in PSTVd-infected potato relative to JA-controlled pathways, we analyzed shared genes with contrasting expression between wild-type and transgenic lines (Figure S4A). KEGG analysis showed that DEGs upregulated in coi1 but downregulated in wild-type were enriched in alanine, aspartate, and glutamate metabolism, as well as phenylpropanoid biosynthesis (Figure S4B, Table S6). In contrast, DEGs downregulated in coi1 but upregulated in wild-type were enriched in pentose and glucuronate interconversions, amino sugar and nucleotide sugar metabolism, and plant-pathogen interaction pathways, highlighting the role of COI1-dependent JA signaling in the potato-PSTVd interaction. No enriched pathways were detected in the comparison between opr3 and wild-type.
Identified DEGs were further analyzed using RT-qPCR. Transcript accumulation estimates for 15 selected genes from RNA-Seq closely matched those from RT-qPCR, indicating that the RNA-Seq data are reliable (Figure S5).
3.3. Expression Profiles of Hormone-Related, MAPK, and PR Genes
According to KEGG functional analysis, most genotype-specific DEGs in all comparisons were enriched in plant hormone signal transduction, plant-pathogen interaction, and MAPK signaling pathways. These differences were validated with independent data from RT-qPCR and hormone analyses. The dynamic expression patterns of selected phytohormone-related genes were analyzed at different time points, before and after symptom onset (Figure 4, Table S7). The same tissue samples were used for hormone quantification (Figure 5).
After PSTVd infection, earlier activation of JA biosynthesis and perception genes (LOX6, OPR3, JAR1, and COI1) was detected in wild-type and coi1 plants compared to opr3 plants. Conversely, activation of JA signaling genes (JAZ1 and MYC2) was stronger in infected opr3 plants. Hormone analysis showed that the level of cis-OPDA, a precursor of JA-Ile, peaked at 6 wpi and then decreased at 7 wpi in all tested lines, while significant JA-Ile accumulation was detected only in infected wild-type plants. SA biosynthesis (ICS and PAL9) and SA signaling (SARD1 and NPR1) genes were upregulated in all tested lines (opr3 > coi1 > wt); however, SA levels remained unchanged. ABA content and ABA hydroxylase gene expression increased only in opr3 plants at the early stage of infection, while increased expression of ABI5 and MYB44 suggested activation of ABA signaling in all tested lines. The relative expression of auxin-responsive genes (ILR1, ARF8, and EXP8) was stronger in opr3 and coi1 compared to wild-type, while IAA content increased slightly in opr3 and wild-type but decreased in coi1, after PSTVd infection. PSTVd-induced activation of the BR receptor gene BRI1 was higher in opr3, coinciding with a slight increase in castasterone (CS) content. The relative expression of ET biosynthesis (ACO) and ET signaling (ERF1B and AP2-TOE3) genes was elevated in all tested lines at 5–7 wpi (coi1 > opr3 > wt).
Since cross-talk among MAPK, hormone, and ROS signaling pathways is important for the fine-tuned modulation of plant immunity [47,48], we measured the time course of expression of selected MAPK and PR protein genes after PSTVd infection (Figure 4, Table S7). Results showed activation of MPKK6, MAPK4, and MAPK3 genes in wild-type plants at the early stage, but in opr3 and coi1 plants at the late stage of infection. Expression of the gene for MAPK7, an H_2_O_2_-responsive MAPK [49], was upregulated in all tested lines. Relative PR1b gene expression was stronger in wild-type plants, while PR2 and PRQ gene expression was stronger in opr3 and coi1 plants, after PSTVd infection. Activation of LOX3, a pathogen-responsive lipoxygenase, was strongest in infected coi1 plants.
3.4. Regulation of Redox Processes in opr3, coi1, and Wild-Type Potato Plants During PSTVd Infection
To investigate the mechanisms underlying altered H_2_O_2_ accumulation in JA-deficient plants, we analyzed transcript levels of ROS-producing (RBOH) and ROS-scavenging genes, including catalase (CAT2), ascorbate peroxidase (APX1), suberization-associated anionic peroxidase (POPA), lignin-forming anionic peroxidase (LiP), and peroxidase 12 (POX12). After PSTVd infection, wild-type plants accumulated H_2_O_2_, while opr3 and coi1 plants showed reduced H_2_O_2_ levels at the late stage of infection (Figure 6A,B). RT-qPCR revealed earlier activation of RBOH, CAT2, and APX1 genes in coi1 and opr3 plants compared to wild-type plants (Figure 6C, Table S7). For Class III peroxidases, a plant-specific family of antioxidant enzymes involved in diverse functions [50], POPA expression was upregulated in wild-type plants, LiP was upregulated in coi1 and opr3 plants, and POX12 expression was upregulated in opr3 plants, but downregulated in coi1 plants.
3.5. Effect of MeJA Treatment on Defense Responses in PSTVd-Infected Potato
Treatment with MeJA alleviated the negative effects of PSTVd infection on apical leaf and stem growth in wild-type plants, but had no significant effect on symptom appearance in opr3 and coi1 plants (Figure 7 and Figure S6).
In PSTVd-infected wild-type plants, MeJA caused a transient decrease in the expression of nearly all tested genes involved in JA, SA, MAPK, and ROS signaling (Figure 8, Table S8), along with an increase in H_2_O_2_ content (Figure S6). In PSTVd-infected opr3 plants, MeJA initially decreased but then increased the expression of JA-related and SA-related genes, compared to untreated infected opr3 plants. A decrease in H_2_O_2_ content was also observed. As expected, MeJA had little effect on the expression of selected genes and H_2_O_2_ content in JA-insensitive coi1 plants. Despite its notable impact on gene expression and symptom development, MeJA did not significantly affect the dynamics of PSTVd RNA accumulation in any of the tested lines (Figure S6).
4. Discussion
Functional analyses of differentially expressed genes (DEGs) showed that PSTVd infection in all tested potato lines suppressed genes related to photosynthesis and primary metabolism, while activating genes involved in proteolysis, signaling, and defense responses. This indicates a redirection of energy from metabolic processes to defense, a widespread and complex feature of pathogen-infected plants [23,51,52]. The late stage of infection in wild-type plants was characterized by the accumulation of JA-Ile, consistent with our previous results [25]. As expected, the transgenic opr3 and coi1 plants produced less JA-Ile than the wild-type, but no significant increase in JA-Ile levels in response to PSTVd was observed. However, the relative expression of the JA signaling gene MYC2 was higher in opr3 plants, while the expression of early JA biosynthesis genes such as LOX was higher in coi1 plants. We hypothesize that this may be related to increased production of other jasmonates and oxylipins [10,14]. Both opr3 and coi1 transgenic lines had higher viroid loads than the wild-type, suggesting that JA is important for the basal defense response of potato against PSTVd. Transient accumulation of cis-OPDA, a precursor of JA-Ile and a JA-independent signaling molecule [53], was observed in infected plants of all tested lines. Therefore, it likely did not contribute to the difference in susceptibility to PSTVd among the tested lines, which requires further investigation.
Consistent with its defensive role, JA accumulation may inhibit plant growth [54]. When infected with PSTVd, JA-deficient opr3 plants showed milder symptoms in leaf and stem growth, while JA-insensitive coi1 plants exhibited significantly elongated stems, contrary to the typical PSTVd symptom of stunted stem growth. Detailed analysis of PSTVd-induced transcriptomic, hormone, and physiological responses in transgenic opr3 and coi1 lines, compared to the wild-type line, revealed several phenomena possibly associated with altered basal defense responses and symptom development in these lines: (i) activation of other hormone signaling pathways and their interactions with the JA signaling pathway; (ii) alteration of MAPK signaling, transcription factors, and PR gene expression; and (iii) transcriptional modification of antioxidant responses and metabolic pathways.
4.1. Activation of Other Hormone Signaling Pathways and Their Interactions with the JA Signaling Pathway
An imbalance between JA and AUX pathways can increase plant susceptibility to pathogens because high AUX levels promote cell growth, facilitating pathogen invasion and reducing the plant’s ability to activate JA-mediated defense responses [55]. At the transcriptome level, PSTVd-infected opr3 plants showed increased expression of several AUX metabolism and early AUX signaling genes, including LAX3, SAURs, and AUX/IAA genes [56]. For example, expression of StGH3.1 (an ortholog of AtGH3.1 and OsGH3.8 [57] was upregulated in JA-insensitive and JA-deficient plants compared to wild-type plants, suggesting that JA negatively regulates GH3.1 expression in response to PSTVd in potato. An interplay between JA and AUX signaling pathways during potato-PSTVd interaction is also suggested by the differential expression of auxin-responsive genes ARF8 [56] and EXP8 [58] in opr3 and coi1 lines compared to wild-type plants. We hypothesize that impaired JA biosynthesis and signaling affect the regulation of AUX signaling, contributing to symptom alleviation in opr3 and coi1 plants to PSTVd infection. However, auxin content remains relatively unchanged after PSTVd infection in all lines tested.
Abscisic acid mediates abiotic stress tolerance and regulates growth and development, but its role in disease resistance is complex and depends on the specific plant-pathogen interaction [5]. Jasmonate deficiency increases stress sensitivity in rice, and ABA biosynthesis or catabolism in response to osmotic stress can be modulated by JA [59]. Similarly, ABA accumulation in the early stage of PSTVd infection and increased expression of genes encoding the transcription factors MYB44 and MYC2 suggest enhanced stress response and stomatal regulation in JA-deficient opr3 plants. Both MYB44 and MYC2 contribute to stomatal closure under stress conditions; however, MYC2 acts through JA signaling and stimulation of ABA biosynthesis [60], while MYB44 acts through ABA signaling [61]. Multifunctional MYB44 can also act as a negative regulator of ABA signaling in leaf senescence [61]. In contrast, in JA-insensitive coi1 plants, ABA levels decreased, while the expression of genes for ABA receptors PYR1 and PYL4 [62] and the transcription factor ABI5-like protein 5 increased, suggesting activation of ABA signaling in response to PSTVd infection. ABA receptors and ABI transcription factors can, in turn, influence other signaling pathways, including the JA pathway, to fine-tune downstream stress responses and growth regulation [63].
Ethylene can have both negative and positive effects on plant defense responses and often acts synergistically with JA to activate defense-related genes [6,64]. Coordinated crosstalk between ET and JA converges at the transcriptional activation of ERF1. Arabidopsis ERF1 is an early JA-responsive gene in the ERF branch of the JA defense response pathway, while ethylene promotes a longer-lasting induction of ERF1 expression [64,65]. The AP2/ERFs, in addition to their role in hormone crosstalk, are also recognized for mediating plant responses to biotic stress, as transcriptional activators or repressors [66,67]. About half of the PSTVd-induced genes shared by wild-type, opr3, and coi1 lines encode ET-responsive proteins (GRL1, ETR2-like, EBF1-like, and ERF1B-like), suggesting that activation of ET signaling is a core response to PSTVd in potato. However, the total number of ET-related DEGs in response to PSTVd was significantly higher in opr3 and especially in coi1 plants, which may also suggest an alternative, JA-independent activation of ET signaling and/or that COI1 is not completely silenced [26]. The involvement of ET signaling in other plant-viroid interactions has also been reported [11,17,68,69,70], while ET accumulation was mostly associated with symptom development [71]. To clarify the role of ET in the potato response to PSTVd, future studies should analyze ET content.
Several lines of evidence indicate that suppression of GA biosynthesis genes, which is associated with reduced GA responses, plays a major role in the development of stunting in viroid-infected plants [16,17,24,56,68,72]. Our study showed downregulation of the GA receptor GID1B gene, and upregulation of GA-responsive RSI-1 genes after PSTVd infection in wild-type and opr3 plants, while in coi1 plants, the infection was accompanied by opposite responses. StRSI-1 is an ortholog of AtGASA5 (Gibberellin-stimulated transcript GAST1 protein homolog 5), a downstream gene of DELLA, which acts as a suppressor of gibberellin responses and stem growth in Arabidopsis thaliana [73]. In addition, GASA proteins are involved in JA-dependent biotic defense responses [74,75]. Therefore, we hypothesize that the increased expression of RSI-1 in PSTVd-infected coi1 plants may be associated with the absence of stem stunting symptoms in coi1 plants, as part of the crosstalk between growth-promoting GA signaling and defense-promoting JA signaling [54].
Regarding the other hormones analyzed in this study, transcriptomic data showed a limited effect of PSTVd infection on the activation of SA and BR biosynthesis genes. This is consistent with the small changes in endogenous SA and CS content measured in infected compared to uninfected plants across all three lines. However, RT-qPCR analysis detected increased expression of SA biosynthesis (ICS and PAL9) and SA signaling (NPR1 and NPR3) genes in wild-type plants during the early stage, and in opr3 and coi1 plants during the late stage of PSTVd infection. Unlike Arabidopsis, in some plant species such as potato and rice, SA levels do not increase significantly after pathogen infection, but the SA signaling pathway remains active and contributes to disease resistance [76,77].
Interestingly, transcriptome analysis also revealed contrasting expression of genes involved in the biosynthesis and metabolism of growth-promoting hormones CK (IPT- and ZOG-like) and PSK (phytosulfokines 3-like) in opr3 and coi1 plants infected with PSTVd. This suggests a possible interaction between CK, PSK, and JA signaling [78] during potato-PSTVd interaction, but further investigation is required.
4.2. Alteration of MAPK Signaling, Transcription Factors, and PR Genes in Response to PSTVd
MAPK cascades have been reported to regulate JA biosynthesis and JA-dependent gene expression [79]. JA also regulates both MAPK activity and MAPK gene expression, and silencing JA-related genes can affect MAPK activity [48]. After PSTVd infection, late activation of MAPK3 and MAPK4 genes was observed in opr3 and coi1 plants compared to wild-type plants, while pretreatment with MeJA accelerated gene activation in opr3 plants. In Arabidopsis, MAPK4 acts as a negative regulator of SAR and SA signaling but as a positive regulator of JA-dependent gene expression [80]. Our transcriptome analysis also revealed coi1-specific activation of a MAP3KA-like gene [81] and opr3-specific activation of SAPK3-like and MAP2K-like genes [82], suggesting their pathogen-dependent but JA-independent activation in potato. In addition, we detected PSTVd-induced expression of MAPK7 in all tested potato lines, in all tested lines, which may be related to changes in JA and H_2_O_2_ status [49] associated with PSTVd infection.
By integrating signals from both hormones and ROS, the MAPK pathways can orchestrate a more precise and efficient plant defense response [47,83]. MapMan analysis showed that most defense-related genes, except miraculin [84], were upregulated in wild-type and opr3 plants compared to coi1 plants after PSTVd infection. Additionally, the expression of PR1 genes [85] was lower and PR2 genes [86] higher in opr3 and coi1 plants, which may be related to their higher susceptibility to PSTVd compared to wild-type plants, as indicated by viroid RNA accumulation. These results suggest the activation of alternative defense responses in opr3 and coi1 plants and highlight the role of JA signaling in regulating PR gene expression during PSTVd infection, possibly through interactions with other signaling pathways, including the ET and SA pathways [87].
Transcription factors (TFs) play a key role in establishing plant defense and symptom development during viroid infections [56,72]. The importance of bHLH TFs, including MYCs that directly interact with JA-responsive genes, in the tomato-PSTVd interaction has also been reported [88]. In our study, the primary TF families showing significant responses to PSTVd in potato were identified, and the potential role of certain TFs in regulating hormone, MAPK, and ROS signaling during the potato-PSTVd interaction is discussed. However, given the numerous TFs and their diverse roles in regulating various defense-related genes, further research on specific TFs in the potato-PSTVd interaction is needed.
4.3. Transcriptional Modification of Antioxidant Responses and Metabolic Pathways in opr3 and coi1 Plants During PSTVd Infection
The physiological effects of JA include activation of ROS signaling pathways and recruitment of the cellular antioxidant defense system to maintain moderate ROS levels [89]. After PSTVd infection, DEGs for enzymes that regulate the cellular redox state, including APX [90], were mainly downregulated in opr3 and coi1 compared to wild-type. Since opr3 and coi1 plants accumulate less H_2_O_2_ than wild-type plants after PSTVd infection, their reliance on APX is less pronounced. However, not all antioxidant responses followed this pattern. DEGs for glutathione S-transferases were mainly induced by PSTVd infection in opr3 and especially in coi1, suggesting their involvement in ROS reduction in plants with compromised JA pathways. JA signaling mediated by COI1 is important for controlling ROS accumulation by influencing the activity of MYC proteins, which regulate the expression of genes encoding antioxidant enzymes, including NOX/RBOH and POX12, involved in ROS production during plant defense against pathogens [89,91]. In PSTVd-infected opr3 plants, one of the most prominent features was increased expression of MYC2, RBOH, and POX12 genes, whereas in infected coi1 plants, both MYC2 and POX12 expression were reduced. Treatment with MeJA followed by PSTVd infection induced short-term H_2_O_2_ production, enhancing POX12 expression, which in turn reduced H_2_O_2_ content in opr3 plants at the late infection stage. High concentrations of MeJA can act pro-oxidatively to stimulate activation of the antioxidant system and some components of basal defense responses [92,93]. Reduced H_2_O_2_ accumulation may enhance the plant’s tolerance to the initial stressor and reduce symptom development, but may also promote pathogen progression [94].
Among other PSTVd-induced POX genes, LiP and POPA encode enzymes that use H_2_O_2_ as a substrate for lignification and suberinization, respectively. LiP and POPA showed contrasting expression patterns in wild-type and opr3 compared with coi1 plants. Increased expression of LiP, along with the lignin biosynthesis genes 4CL and CCoAOMT, suggests increased H_2_O_2_ consumption during monolignol biosynthesis in infected coi1 plants. Lignin is a complex polyphenolic polymer that serves as a structural component in plant cell walls, providing rigidity and strength, and acts as a physical and chemical barrier in basal plant immunity [95]. Lignification is negatively regulated by JA [92], which is consistent with the observed increased expression of lignification-related genes in coi1 plants. In contrast, several PAL genes were specifically upregulated in wild-type plants after PSTVd infection, suggesting the production of various other phenolic secondary metabolites [96]. Altered expression of many genes encoding lignin-forming peroxidases and genes from the lignin-specific pathway was also observed in PSTVd-infected tomato [18].
Plants with impaired JA biosynthesis or signaling often activate alternative metabolic pathways, due to a bypass in the JA biosynthesis pathway, allowing accumulation of alternative defense compounds [7]. In PSTVd-infected opr3 plants, genotype-specific activation of genes related to cutin, suberin, and wax biosynthesis (CYP94A1 and feruloyl transferase FHT), and glycerophospholipid metabolism (glycerol-3-phosphate acyltransferases GPATs), was observed. Glycerolipids are important components of plant membranes, which are essential for development, growth, photosynthesis, and stress responses [97]. Cutin and suberin are protective polyester barriers formed by the polymerization of oxygenated fatty acids and glycerol [98]. Because glycerolipids and JA share fatty acids as precursors, inhibition of one pathway enhances regulation of the other, strengthening alternative defense pathways [99,100]. Therefore, we hypothesize that silencing OPR3 alters glycerolipid and oxylipin metabolism and promotes cutin, wax, and suberin production in potato in response to PSTVd and increased osmotic stress in JA-deficient plants [59].
5. Conclusions
This study showed that silencing JA biosynthesis or JA signaling significantly alters the reprogramming of multiple signaling pathways, including hormone, MAPK, and ROS signaling, which disrupts defense regulation and increases susceptibility to PSTVd in potato. JA deficiency at the late stage of infection reduced symptom development in opr3 plants, while MeJA treatment at the early stage of infection reduced symptoms in wild-type plants. The weak coi1 response also supports COI1-dependent MeJA action. However, because only one MeJA dose and timing were used, conclusions about timing and dose–response are limited. Further studies, such as testing different MeJA concentrations, applying stage-specific inhibitory treatments, and conducting more detailed time-course sampling, should be conducted to provide more direct evidence supporting a positive role for JA in the early stage of PSTVd infection. Numerous transcriptional changes caused by PSTVd infection in coi1 plants indicate the activation of alternative responses that compensate for compromised JA signaling, leading to symptom relief but increased viroid accumulation. This is consistent with the established role of JA as a regulator in the trade-off between growth and defense in plants [101]. Overall, these results enhance our understanding of the role of JA in basal immune responses against PSTVd and provide valuable gene resources for future functional analyses of key genes and regulatory mechanisms underlying immune responses to this pathogen in potato.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ferranti P. Berry E. Anderson J.R. Encyclopedia of Food Security and Sustainability Elsevier Amsterdam, The Netherlands 20181894 p 9780128126882
- 2Mackie A.E. Barbetti M.J. Rodoni B. Mc Kirdy S.J. Jones R.A.C. Effects of a potato spindle tuber viroid tomato strain on the symptoms, biomass, and yields of classical indicator and currently grown potato and tomato cultivars Plant Dis.20191033009301710.1094/PDIS-02-19-0312-RE 31567060 · doi ↗ · pubmed ↗
- 3Navarro B. Flores R. Di Serio F. Advances in viroid-host interactions Annu. Rev. Virol.2021830532510.1146/annurev-virology-091919-09233134255541 · doi ↗ · pubmed ↗
- 4Sano T. Progress in 50 years of viroid research-molecular structure, pathogenicity, and host adaptation Proc. Jpn. Acad. Ser. B Phys. Biol. Sci.20219737140110.2183/pjab.97.02034380915 PMC 8403530 · doi ↗ · pubmed ↗
- 5Alazem M. Lin N. Roles of plant hormones in the regulation of host–virus interactions Mol. Plant Pathol.20141652954010.1111/mpp.1220425220680 PMC 6638471 · doi ↗ · pubmed ↗
- 6Aerts N. Pereira Mendes M. Van Wees S.C.M. Multiple levels of crosstalk in hormone networks regulating plant defense Plant J.202010548950410.1111/tpj.1512433617121 PMC 7898868 · doi ↗ · pubmed ↗
- 7Antico C.J. Colon C. Banks T. Ramonell K.M. Insights into the role of jasmonic acid-mediated defenses against necrotrophic and biotrophic fungal pathogens Front. Biol.20127485610.1007/s 11515-011-1171-1 · doi ↗
- 8Tamaoki D. Seo S. Yamada S. Kano A. Miyamoto A. Shishido H. Miyoshi S. Taniguchi S. Akimitsu K. Gomi K. Jasmonic acid and salicylic acid activate a common defense system in rice Plant Signal Behav.20138 e 2426010.4161/psb.2426023518581 PMC 3906320 · doi ↗ · pubmed ↗