Targeting Skin Aging Hallmarks In Vitro: Antioxidant, Anti-Inflammatory, and Anti-Senescence Effects of Phenolic-Rich Extracts from Cistus L. Species
Mário Pedro Marques, Euclides Landim, Carla Varela, Ricardo M. F. da Costa, Joana Marques, Luís A. E. Batista de Carvalho, Ana Silva, Maria Teresa Cruz, Rebeca André, Patrícia Rijo, Maria Inês Dias, Aida Carvalho, Paulo J. Oliveira, Célia Cabral

TL;DR
This study explores how plant extracts from Cistus species may help reduce skin aging by showing antioxidant, anti-inflammatory, and anti-senescence effects in lab tests.
Contribution
The study identifies Cistus ladanifer and Cistus salviifolius as promising sources of skin anti-aging compounds with multiple bioactive effects.
Findings
Cistus ladanifer and Cistus salviifolius extracts showed significant antioxidant, anti-inflammatory, and anti-senescence effects in skin cells.
The extracts inhibited enzymes like elastase and hyaluronidase without causing skin irritation.
Ellagic acid, myricetin, and quercetin derivatives were identified as key compounds in the extracts.
Abstract
Plant-based extracts are rich sources of phenolic compounds, which may act as skin antiaging mediators. Herein, Cistus albidus L. (Ca), Cistus ladanifer L. subsp. ladanifer (Cl) and Cistus salviifolius L. (Cs) were selected to test whether their phytochemical profile and bioactive potential align to target human skin aging. Hydroethanolic extracts (HEs) were prepared and characterized using infrared vibrational spectroscopy (FTIR-ATR) and liquid chromatography–mass spectrometry (LC-MS). Non-toxic concentrations were screened, and cytoprotective and antioxidant effects were studied in tert-butyl hydroperoxide-stimulated normal human dermal fibroblasts (NHDFs). Lipopolysaccharide-stimulated RAW 264.7 macrophages were used to assess anti-inflammatory activity, the Organization for Economic Co-operation and Development (OECD) Test Guideline No. 439 was used to assess irritant effects, and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
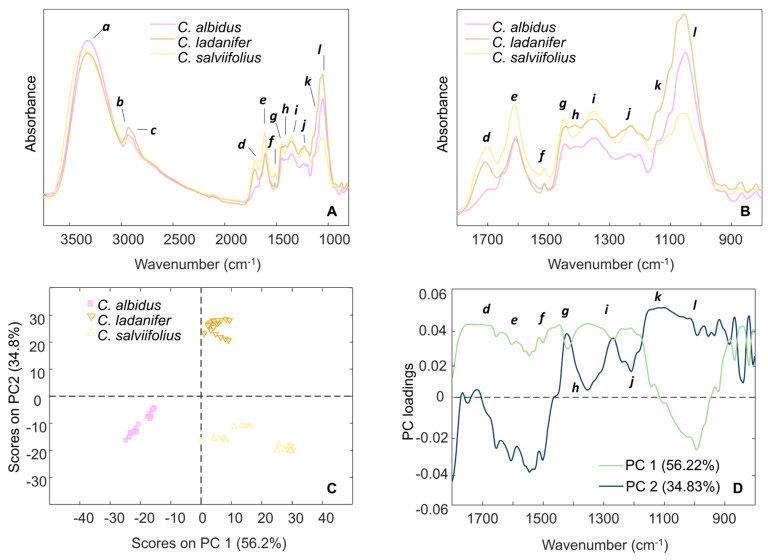 Figure 1
Figure 1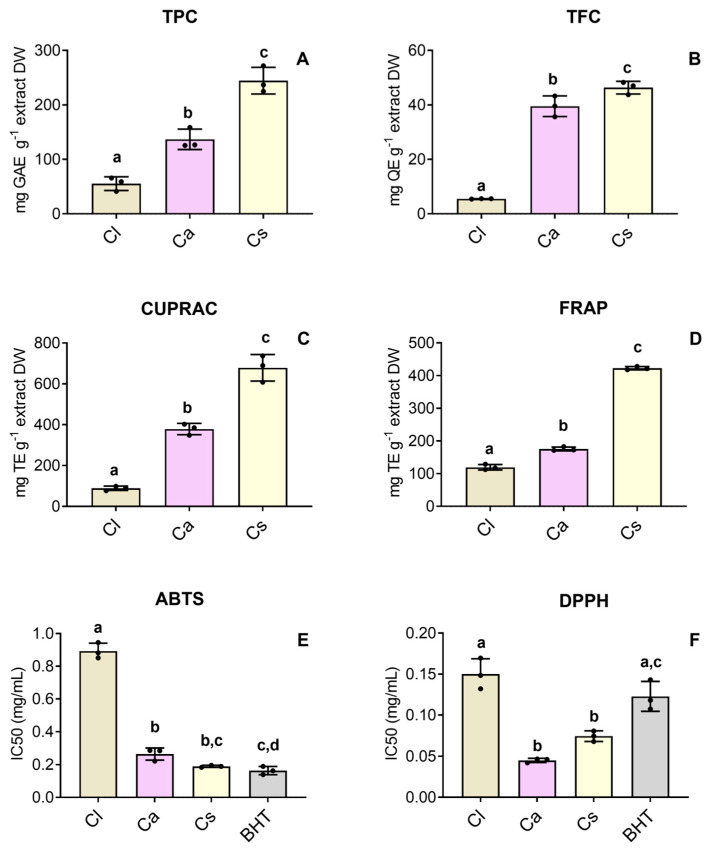 Figure 2
Figure 2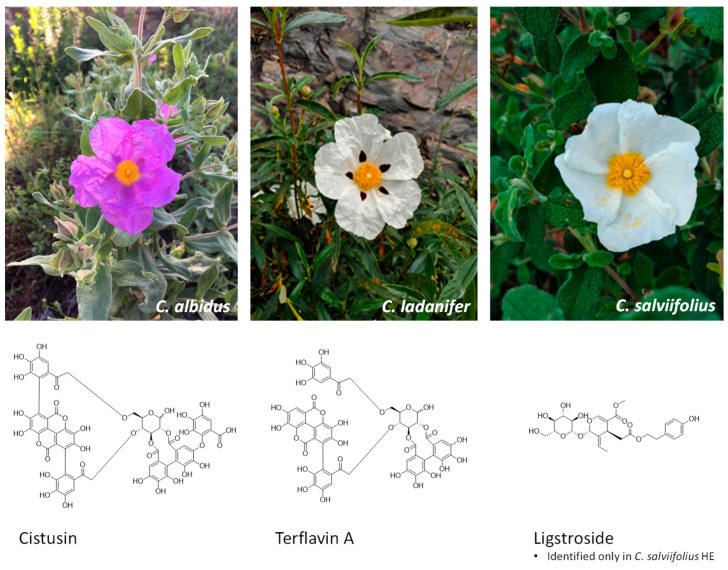 Figure 3
Figure 3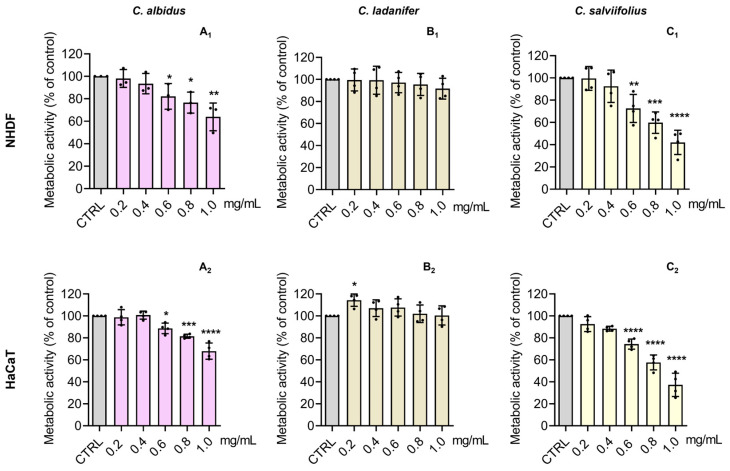 Figure 4
Figure 4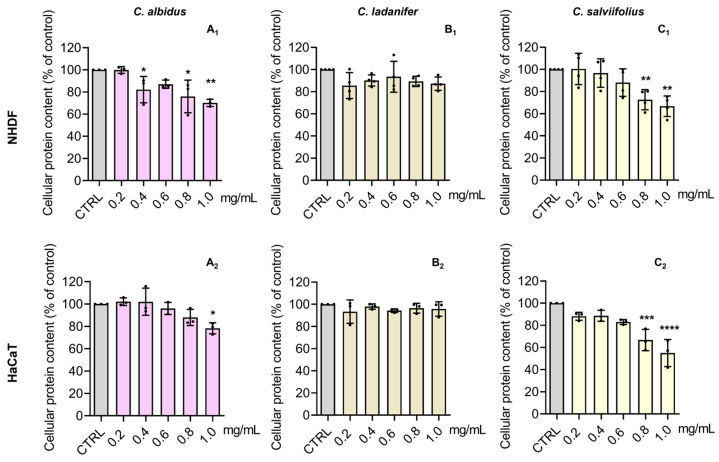 Figure 5
Figure 5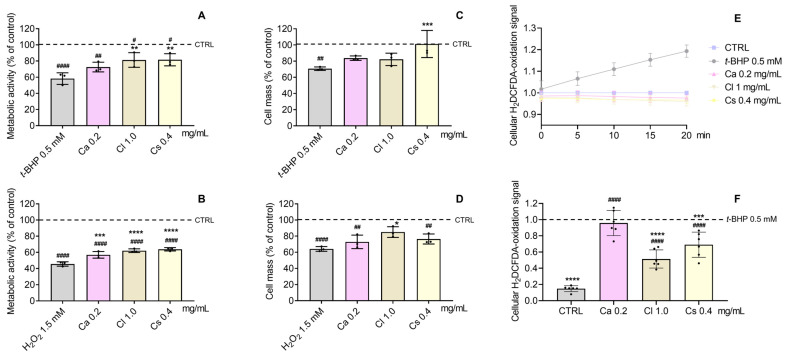 Figure 6
Figure 6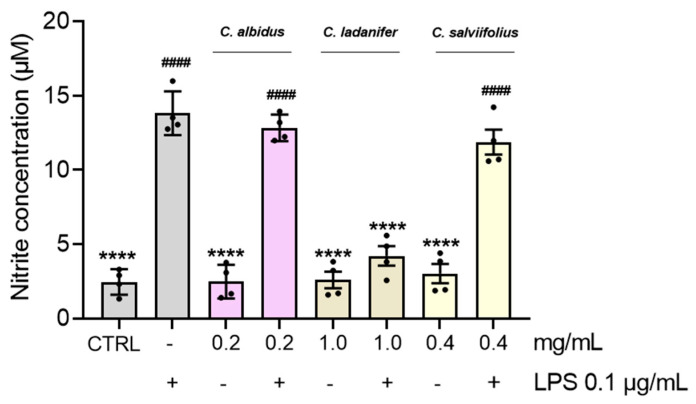 Figure 7
Figure 7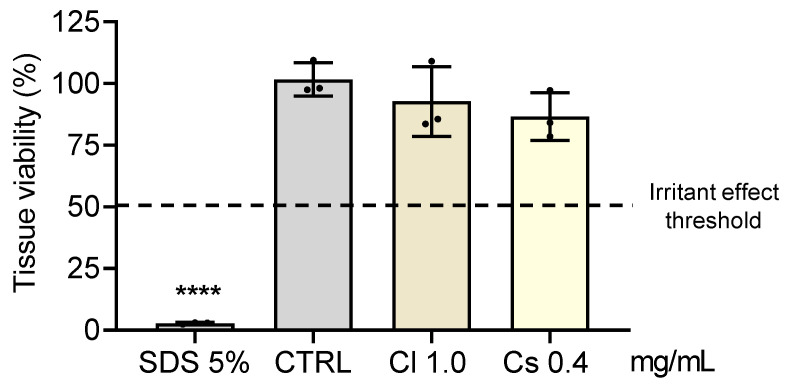 Figure 8
Figure 8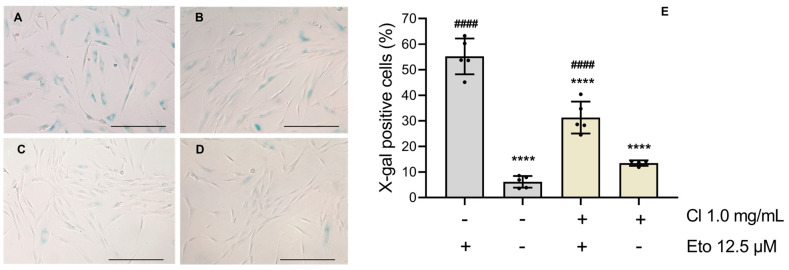 Figure 9
Figure 9- —EU Recovery and Resilience Facility and Portuguese national funds
- —national funds
- —FCT
- —FCT, P.I.
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSesquiterpenes and Asteraceae Studies · Wound Healing and Treatments · Plant chemical constituents analysis
1. Introduction
The skin is primarily composed of a three-layered structure, consisting of (1) the epidermis, in which keratinocytes are the main cells-type at various differentiation stages; (2) the dermis, where fibroblasts play a crucial role in synthesizing extracellular matrix, contributing to the preservation of skin structure and elasticity; and lastly, (3) the hypodermis, fundamentally composed of adipocytes. Covering approximately 1.8 m^2^ of the body’s external surface, the skin is the largest human organ, serving as the primary barrier against pathogens, ultraviolet (UV) radiation, and environmental chemicals or pollutants [1,2]. These stressful conditions may trigger higher levels of reactive oxygen species (ROS) such as superoxide anions (O_2_^•−^), hydrogen peroxide (H_2_O_2_), hydroxyl radicals (^•^OH), and reactive nitrogen species (RNS) such as nitric oxide (NO) [3]. An imbalance between pro-oxidant and antioxidant species may contribute to oxidative injury of biomolecules like structural proteins, lipids, and DNA, promoting cellular senescence or even cell death, processes that are often at the core of both skin aging and pathology [3,4]. Nevertheless, the human body possesses physiological and homeostatic levels of oxidative and inflammatory factors, through an endogenous human antioxidant system constituted by enzymatic and non-enzymatic antioxidants which minimize the oxidative damage in the skin. Cellular antioxidant mechanisms include enzymatic components, as well as non-enzymatic defenses, which are mainly acquired through the diet. These include vitamins (e.g., α-tocopherol and vitamin C), minerals and cofactors, sulfur-containing compounds, non-protein nitrogenous substances, and plant-derived metabolites [4,5,6]. From this perspective, plants constitute a valuable source of chemopreventive compounds, with particular emphasis on polyphenols. These molecules stand out due to their capacity to scavenge ROS/RNS and modulate signaling pathways related to oxidative stress. Their hydroxyl groups are readily oxidized to quinones, endowing them with a powerful redox buffering capacity [6].
The Mediterranean basin is considered a global hotspot of biodiversity [7] and plants native to this biogeographic region are subjected to harsh edaphoclimatic conditions such as high temperatures, prolonged drought, intense UV radiation, and frequent wildfires, which are typical of this biome. These demanding conditions induced plants to develop protective metabolic strategies, particularly the accumulation of phenolic compounds which, for example, confer protection from photo-oxidative stress conditions [8]. Plants from the genus Cistus L. are well represented in the Mediterranean-influenced climatic zone and are known to produce ellagitannins, flavonoids, and phenolic acids derivatives [9]. Interestingly, several works have pointed the beneficial effects of such compounds on human skin cells, constituting natural-based solutions to counteract skin aging [10].
Recently, an emerging interest has driven research studies on Cistus plants [7], particularly those regarding skin application [8,11,12]. Furthermore, ethnomedical practices highlight the use of the Cistus species as traditional remedies for microbial infections, with particular emphasis on their application in the treatment of external wounds [7,13]. To build on this knowledge, we have selected three Cistus species that are particularly abundant in the hillsides of a Portuguese Northeastern region of Portugal, named Côa Valley, which are Cistus albidus L. (Ca), Cistus ladanifer L. subsp. ladanifer (Cl), and Cistus salviifolius L. (Cs). Important ethnobotanical studies provide evidence of their use in the treatment of several skin conditions in different territories of the Iberian Peninsula. As such, C. albidus, locally called hierba lobera in the South Alava region (Spain), has traditional applications in the treatment of skin injuries and infections resulting from wolf bites, with decoctions of its aerial parts used as external washes [14]. Also, in the region of Biscay and Alava (Spain), the leaves from C. salviifolius have ancient topical applications for the treatment of topical wounds and cuts, dog bites, and external ulcers [15]. For instance, in Trás-os-Montes (northern Portugal), the flowers and leaves of C. ladanifer are externally used in cataplasms and baths as an anti-infectious and wound healing treatment [16].
Considering this, although few studies have described the phytochemical composition or selected bioactivities of the Cistus species, most have examined isolated endpoints and lack an integrated evaluation relevant to the context of skin aging. Our work aims to advance the current state of the art regarding these plants species by combining detailed chemical profiling with a broad panel of biological assays that simultaneously address key hallmarks of skin aging, including oxidative stress, inflammation, senescence, enzyme-mediated extracellular matrix components degradation, and robust cell toxicity endpoints. Furthermore, the direct comparison of three species of the same botanical genus under standardized conditions, together with validated safety assessments, offers new insights into species-specific activities and their translational relevance for anti-aging applications. Overall, this multidimensional approach provides a more comprehensive understanding than previously reported of the potential topical application of Cistus extracts as promising candidates to be included in innovative anti-aging formulations.
2. Materials and Methods
2.1. Reagents and Chemicals
From Sigma-Aldrich (St. Louis, MO, USA) we obtained sodium bicarbonate (S5761), D-(+)-Glucose (G8270), Dulbecco’s Modified Eagle’s Medium without glucose (DMEM5030), L-glutamine (G3126), sodium pyruvate (P5280), penicillin–streptomycin solution (P4333), 0.25% Trypsin-EDTA solution (T4049), trypan blue solution (T8154), dimethyl sulfoxide (DMSO) (D2438), 2,2-diphenyl-1-picrylhydrazyl (D9132), 2,4,6-tris(2-pyridyl)-s-triazine (≥99%) (93285), ammonium acetate (≥98%) (A7262), butylated hydroxytoluene (BHT) (≥99%) (W218405), gallic acid (97.5–102.5% (titration)) (G7384), (+)-Catechin hydrate (≥98%) (C125), ferulic acid (PHR1791), quercetin (≥95%) (Q4951), neocuproine (≥98%) (N1501), sodium acetate (≥99%) (S5636), trolox (97%) (238813), iron (III) chloride FeCl_3_·6(H_2_O), (≥98%) (31232-M), Lipopolysaccharide (LPS) from Escherichia coli serotype 026:B6 (L2654), acetylcholinesterase from Electrophorus electricus (electric eel) (C3389), acetylthiocholine iodide (≥98% (TLC)) (A5751), 5,5′-Dithio-bis-(2-nitrobenzoic Acid) (DTNB) (322123), N-(1-naphthyl)-ethylenediamine dihydrochloride (>98%) (222488), sulphanilamide (≥99%) (S9251), tert-Butyl hydroperoxide solution (t-BHP) 70 wt. % in H2O (458139), hydrogen peroxide (H_2_O_2_) 30% (w/w) in H_2_O (H1009), thiazolyl blue tetrazolium bromide (MTT) (≥97.5%) (M5655), sodium dodecyl sulfate (SDS) (≥98.5%) (L4509), N-Succinyl-Ala-Ala-Ala-p-nitroanilide (≥98%) (S4760), tyrosinase from mushroom (T3824), L-Tyrosine (99.0–101.0%) (T8566), kojic acid (≥98.5%) (K3125), and sodium hydroxide (NaOH) (≥98%) (S5881). The reagents aluminum chloride (AlCl_3_, for synthesis) (801081), copper (II) chloride (CuCl_2_, for synthesis) (818247), potassium chloride (KCl) (≥99%) (104936), sodium carbonate (≥99.5%) (106392), and sodium acetate trihydrate (CH_3_COONa^•^3H2O) (99.5%) were obtained from Merck (Oeiras, Portugal). Fetal bovine serum (FBS) (S1810-500) and DMEM Low glucose (P0061-N10L) were acquired from BioWest (Nuaillé, France). Resazurin sodium salt (B21187), Sulforhodamine B sodium salt (SRB) (A14769.06), Tris Base (BP152-1), 2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulphonic acid) diammoniumsalt (≥98%) (J65535.06), p-Coumaric acid (trans-4-Hydroxycinnamic acid) (≥97.5%) (A15167.14), 2′,7′-dichlorodihydrofluorescein diacetate (H_2_DCFDA) (D399), phosphoric acid 85% aqueous solution (A18067.0D), and potassium metaborate hydrate (KBO_2_^•^H_2_O) (99.98%) (047365.30) were purchased from Thermo Scientific Chemicals (Waltham, MA, USA). Glacial acetic acid (27225), methanol (24229), ethanol absolute (24194), hydrochloric acid (HCl) (37–38.0%) (30721), and potassium persulfate (≥99%) (216224) were bought from Honeywell (Carnaxide, Portugal). The Folin–Ciocalteu’s reagent (251567.1609) was purchased from Panreac (Barcelona, Spain). From Carlo Erba Reagents (Milan, Italy) we acquired calcium chloride dihydrate (CaCl_2_^•^2H_2_O) (99.0–105.0%) (327607). Kaempferol (≥98.0%) (7W-GP7425), epicatechin (≥95.0%) (BP-BP0538), and myricetin (≥97.0%) (3B-M2131) were bought from Cymit Química (Pamplona, Spain), while from TargetMol (Linz, Austria) (−)-epigallocatechin gallate (EGCG) (99.43%) (T2988) was purchased.
2.2. Plant Material
Harvested plant material followed a botanical identification carried out by Rosa Pinho, Herbarium Curator of the University of Aveiro, and voucher specimens were deposited at the Herbarium (AVE) of the same university. Detailed information about the harvesting of plants is presented in the Supplementary Materials (Table S1).
2.3. Hydroethanolic Extracts Preparation
The HEs were prepared by the procedure previously described by Amorim and collaborators [17]. Further information about the preparation of extracts is presented in the Supplementary Materials (Table S1).
2.4. FTIR-ATR Spectroscopy Analysis
Fourier-transformed mid-infrared (FTIR) spectra were acquired in attenuated total reflectance (ATR) mode for the obtained freeze-dried extracts in the 3700–400 cm^−1^ range, following the approach previously by Marques and collaborators [18]. Preliminarily, spectra from standard phenolic compounds (Figure S1, Supplementary Materials) were acquired in the same conditions as for plant extracts, as these are among the most common constituents in plant-derived samples.
2.5. HPLC–DAD–ESI/MSn Analysis
The extracts were analyzed following the procedure described by Bessada and collaborators [19] using a Dionex Ultimate 3000 UPLC (Thermo Scientific, San Jose, CA, USA) system equipped with a diode array detector coupled to an electrospray ionization mass detector (HPLC-DAD-ESI/MSn).
2.6. Major Phenolics Estimation and Cell-Free Antioxidant Activity Assessment
Spectrophotometric quantifications of total phenolic content (TPC) and total flavonoid content (TFC), and cell-free antioxidant assays were performed as previously described by Marques and collaborators [20]. For these assays, three independent experiments were performed in triplicate.
2.7. Cell Culture
Normal human dermal fibroblasts (NHDFs, CC-2511, Lonza AG Group, Basel, Switzerland) were cultured in low-glucose (5 mM) Dulbecco’s modified Eagle’s medium (DMEM) (pH 7.3), The human keratinocytes (HaCaT, CLS 300493, Eppelheim, Germany) were cultured in DMEM high glucose (25 mM) (pH 7.3). The media of both NHDF and HaCaT cell lines were supplemented with 10% (v/v) heat-inactivated FBS and 1% (v/v) penicillin–streptomycin solution. The mouse leukemic macrophage cell line (RAW 264.7, ATCC TIB-71, Manassas, VA, USA) was cultured in DMEM high glucose (25 mM) (pH 7.3), supplemented with 10% (v/v) non-inactivated FBS, 1% (v/v) penicillin–streptomycin solution.
2.8. Cell Metabolic Activity
The effect of HEs on the metabolic activity of normal skin cells, specifically NHDFs and HaCaT, was evaluated following the resazurin reduction principle and the protocol established by Silva and collaborators [21].
2.9. Cellular Protein Content
Sulforhodamine B (SRB) assay was performed to evaluate the influence of extracts on cellular protein content as previously stated by Silva and collaborators [21].
2.10. Cell-Free Enzymatic Inhibition Assays
The anti-elastase and anti-tyrosinase assays were assessed following the procedures of Andrade and collaborators [22] and the anti-hyaluronidase as stated by Ratnasooriya et al. [23]. For these assays, five independent experiments were carried out in triplicate. The results were expressed as a percentage (%) of inhibition.
2.11. Measurement of Cellular Nitrites Production
The procedure previously published by Moreira and collaborators [24] was carried out to determine nitrite production in lipopolysaccharide-stimulated RAW 264.7 cells.
2.12. Evaluation of Cytoprotective Efficiency
The cytoprotective effect of HEs in the presence of the oxidative stressors tert-butyl hydroperoxide (t-BHP) and hydrogen peroxide (H_2_O_2_) was evaluated in NHDFs following previous procedures [25]. Different peroxides were chosen based on their different mechanisms of action that differ slightly, and preliminarily, dose–response curves were produced to find a suitable concentration of t-BHP (0.5 mM) and H_2_O_2_ (1.5 mM).
2.13. Determination of Intracellular Oxidative Stress
To evaluate intracellular oxidative stress, the oxidation of the H_2_DCFDA fluorescent dye was measured as previously described by Pinho and collaborators [26].
2.14. Skin Irritant Effects
Skin irritation was evaluated through a standard operating procedure using the SkinEthic™ Reconstructed Human Epidermis (RHE) model (EPISKIN Laboratories, Lyon, France), in compliance with OECD guidelines, and as previously described [24].
2.15. Senescence-Associated β-Galactosidase Activity
The anti-senescence potential was assessed as previously stated [24] with minor changes. Briefly, NHDF cells were seeded in 12-well plates at 1.5 × 10^4^ cells/well and allowed to proliferate for 24 h. Afterwards, C. ladanifer HE was added to the cells for another 24 h. Then, the HE was removed, and etoposide (12.5 μM) was added for 24 h more. After inducing senescence, fresh medium without HE and/or etoposide was added, and the cells were allowed to develop a senescence-associated phenotype for 6 days. The senescence-associated β-galactosidase activity of etoposide-stimulated NHDFs was evaluated following the manufacturer’s instructions for the commercial kit used (CS0030; Sigma-Aldrich). Senescent cells were ultimately imaged with a widefield microscope at 40× magnification by counting the proportion of X-gal positive cells across four randomly chosen fields. Five independent experiments, with two experimental replicates per condition, were performed.
2.16. Statistical Analysis
The results are presented as the mean ± standard deviation (SD) of the indicated number of independent experiments. D’Agostino’s, Pearson’s, and Shapiro–Wilk’s normality tests were conducted to check the normality of the data distribution. All calculations for descriptive statistics, one-way analysis of variance (ANOVA), as well as Tukey’s, Dunnett’s, and Sidak’s range tests were performed using GraphPad Prism 9.0 software (GraphPad Software, La Jolla, CA, USA) considering the following significance values: * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, as well as ^#^ p < 0.05, ^##^ p < 0.01, ^###^ p < 0.001, ^####^ p < 0.0001 for Dunnett’s and Sidak’s tests, while for Tukey’s test significant values (p < 0.05) were represented by lowercase letters (a–d).
3. Results
3.1. FTIR-ATR Spectroscopy
As an initial exploratory step, the FTIR-ATR spectra of the hydroethanolic extracts (HEs) from the studied Cistus species were recorded in the range 3700–800 cm^−1^ (Figure 1A), as well as from standard phenolic compounds (Figure S1). Spectral bands were assigned according to chemical functional group (Table 1). Notably, the region 1800–800 cm^−1^ displayed pronounced differences among the samples, revealing substantial compositional diversity (Figure 1B).
To explore inter-sample variability, principal component analysis (PCA) was applied to the FTIR-ATR data. The first two principal components (PC1 and PC2) accounted for 91.0% of the total variance, with PC1 explaining 56.2% and PC2 34.8% (Figure 1C). Along PC1, the spectra of the HE from C. albidus flowering aerial parts was clearly separated, positioned on the negative side. Conversely, the spectra of C. ladanifer and C. salviifolius HEs clustered on the positive side of PC1. This pattern indicates a higher degree of compositional similarity between C. ladanifer and C. salviifolius compared with C. albidus. Further separation was observed along PC2, with C. ladanifer extracts positioned in the positive quadrant, indicating unique spectral characteristics compared to both C. albidus and C. salviifolius. The interpretation of the corresponding loading plots (Figure 1D) highlights the key spectral regions responsible for this segregation, providing further insight into the relative compositional differences between the HEs of the studied Cistus species.
3.2. Total Phenolic and Total Flavonoid Contents and Cell-Free Antioxidant Activity
Regarding TPC (Figure 2A) and TFC (Figure 2B) determinations, the ranking order points consistently to greater amounts of both TPC and TFC in C. salviifolius, followed by C. albidus, and lastly, C. ladanifer. Similarly, the studied HEs registered the same tendency regarding the CUPRAC and FRAP assays, being more effective in reducing copper than iron (Figure 2C and Figure 2D, respectively), with substantially higher CUPRAC activity, ranging between 89.102 ± 10.88 TE g^−1^ extract DW in C. ladanifer and 678.517 ± 65.08 TE g^−1^ extract DW in C. salviifolius. Also, the HEs of C. salviifolius and C. albidus presented a strong ability to inhibit DPPH^•^ (Figure 2F), even outperforming the reference antioxidant butylated hydroxytoluene (BHT; IC_50_ = 0.123 ± 0.018 mg/mL). However, regarding the ABTS assay, the studied HEs did not show the same potential, and only C. salviifolius became close to the BHT inhibition potential (Figure 2E). The determined values (mean ± SD) in these assays are summarized in the Supplementary Materials (Table S2).
3.3. HPLC-DAD-ESI/MSn Analysis
Among the studied samples, 42 compounds were tentatively identified, including 18 in the HE from C. albidus, 9 in the HE from C. ladanifer and 15 in the C. salviifolius HE. The chromatographic and mass responses, as well as the respective tentative identification of the phenolic compounds found, are described in Table 2. The chemical structures of the most relevant phenolics identified in HEs are illustrated in Figure 3. The illustrative phenolic profiles recorded at 280 and 370 nm, are presented in the Supplementary Materials (Figure S2).
Similarly to C. ladanifer and C. salviifolius, two ellagic acid derivatives were found in C. albidus HE: peak 2*^Ca^* (terflavin A) and peak 4*^Ca^(cistusin). Phenolic acids were mainly represented by peak 3^Ca^* ([M-H]^−^ m/z 337, 3-O-p-coumaroylquinic acid) and vanillic acid hexoside (18*^Ca^, [M-H]^−^ m/z 329). Peak 1^Ca^* ([M-H]^−^ m/z 305, gallocatechin) and peak 5*^Ca^* comprised the flavan-3-ols group. Major phenolics were O-glycosylated flavonoids: kaempferol 7-O-(6″-p-coumaroyl)hexoside (16*^Ca^), myricetin, and quercetin derivatives linked to hexosyl, malonyl-hexosyl, pentosyl, and deoxyhexosyl moieties, represented by the peaks 6^Ca^* to 15*^Ca^. The C. ladanifer HE showed C- and O-glycosylated flavonoids (peaks 5′^Cl^, 5⁗^Cl^, 6′^Cl^, 18′^Cl^), ellagitannins (terflavin A 2^Cl^, cistusin 4^Cl^), and dihydroxy-tetramethoxyflavone (5″^Cl^). C. salviifolius HE presented ellagic acid derivatives (terflavin A 1^Cs^, cistusin 2^Cs^), and a secoiridoid (3^Cs^, [M-H]^−^ m/z 453), and several flavonoids were found, including various myricetin and quercetin derivatives (peaks 8^Cs^–13^Cs^*), confirming that plants from the genus Cistus have a highly diverse phenolic profile.
3.4. Cytotoxic Effects on Normal Human Skin Cells
NHDF cells have been used as a toxicological model for skin applications [25,43] and skin fibroblasts are often employed to assess plant-derived phenolics [10]. HaCaT cells, resembling in vivo keratinocytes, are also suitable for cytotoxicity studies [24]. The cytotoxic effect of HEs (0.2–1.0 mg/mL) from Cistus spp. was tested in NHDF and HaCaT cells regarding metabolic activity (Figure 4) and cellular protein content (Figure 5). C. albidus and C. salviifolius showed no significant impact at ≤0.2 mg/mL and ≤0.4 mg/mL, respectively, while C. ladanifer caused no relevant effects across tested doses. Considering both parameters, the toxicity ranking was C. ladanifer (≤1.0 mg/mL, least toxic) < C. salviifolius (≤0.4 mg/mL) < C. albidus (≤0.2 mg/mL, most toxic).
3.5. Antioxidant and Cytoprotective Effects
According to our results (Figure 6), t-BHP (0.5 mM) and H_2_O_2_ (1.5 mM) decreased cell metabolic activity by ~40% and ~60%, respectively. C. salviifolius and C. ladanifer counteracted t-BHP effects (p < 0.01) (Figure 6A), while all HEs antagonized H_2_O_2_-induced decrease (Figure 6B). For cellular protein content, only C. salviifolius (p < 0.001 versus t-BHP) and C. ladanifer (p < 0.05 versus H_2_O_2_) prevented reductions (Figure 6C and Figure 6D, respectively). Given these cytoprotective effects, intracellular oxidative stress was assessed via H_2_DCFDA assay (Figure 6E,F). No pro-oxidant activity was observed in HEs-treated NHDFs, unlike t-BHP, which significantly increased oxidation (Figure 6E). C. ladanifer (p < 0.0001) and C. salviifolius (p < 0.001) markedly reduced oxidative stress in t-BHP-treated cells, whereas C. albidus showed no protection (Figure 6F).
3.6. Anti-Inflammatory Effects Assessed by Nitrite Quantification
Nitrite levels were investigated in LPS-activated macrophages to determine the anti-inflammatory potential of HEs at their highest non-toxic concentrations (Figure 7). Therefore, macrophages were subject to HE treatment in the presence (+) of LPS to unveil their potential anti-inflammatory effects, also without (−) LPS stimulation, showing the absence of pro-inflammatory effects on these cells, with a production of nitrites comparable to the CTRL. A cytotoxic assessment of these HEs on RAW 264.7 cells was also carried out (Supplementary Materials, Figure S3). Overall, the results showed a significant inhibition of nitrite production (p < 0.0001) when macrophages were treated with C. ladanifer HE by reducing nitrite levels by more than half compared with cells treated with LPS alone.
3.7. Cell-Free Evaluation of Enzyme Inhibitory Activity
The efficacy of Cistus spp. HEs, at their highest non-cytotoxic concentrations, in inhibiting the activity of four key enzymes associated with skin aging was assessed using cell-free enzymatic assays (Table 3). In general, C. salviifolius HE consistently presented a remarkable ability to inhibit all tested enzymes, with comparable inhibition rates to the used positive controls. Alongside this, the HE of C. ladanifer also showed remarkable potential against elastase inhibition (around 65%), both presenting a comparable effect to the positive control. Concerning hyaluronidase’s inhibition, neither C. albidus nor C. ladanifer presented the capacity to inhibit this enzyme. Regarding tyrosinase inhibitory activity, C. salviifolius reached the effect of the positive control kojic acid.
3.8. Skin Irritation
Considering the promising results obtained throughout this study with C. salviifolius and C. ladanifer extracts, their potential to evoke skin irritation was evaluated in a reconstructed human epidermis 3D model (SkinEthic™ Reconstructed Human Epidermis), in compliance with the OECD Test Guideline No. 439. According to ISO 10993-10: 2010 [44], a substance is considered a non-irritant if it does not reduce tissue viability to ≤50% (as observed for SDS, used as positive control). Since the tissues exposed to both extracts presented tissue viabilities higher than 50%, the results demonstrated the absence of skin-irritating effects for these HEs (Figure 8).
3.9. Anti-Senescence Potential
Considering the potential of C. ladanifer HE on several age-related skin dysfunctions tested in this work, its effect on cellular senescence was assessed in terms of senescence-associated β-galactosidase activity (Figure 9). As such, etoposide was shown to evoke a significant (p < 0.0001) increase in X-galactose (X-gal)-positive cells (50–60%; Figure 9A,E), in comparison to untreated NHDF cells (Figure 9C). In turn, for the group of pre-treated NHDF cells (Figure 9B) with C. ladanifer HE, the number of X-gal-positive cells (around 30%; Figure 9E) was significantly lower (p < 0.0001).
4. Discussion
In this study, we prospected the potential of three Cistus spp. for the development of innovative plant-based solutions to target skin aging, and for that purpose, hydroethanolic extractions (80:20%, v/v) were employed for their efficiency in recovering a broad range of polar and non-polar compounds [17]. As such, we first established their chemical profiles through FTIR-ATR spectroscopy and the chemometric analysis of the FTIR-ATR spectra revealed distinct clustering patterns, with C. ladanifer and C. salviifolius showing a closer compositional similarity to each other than to C. albidus. This distinction was particularly evident in the region 1800–800 cm^−1^, with notable variation in the intensity of the carbonyl stretching band around 1701–1693 cm^−1^ (d), which is prominent in C. salviifolius and C. ladanifer spectra, but weak or absent in C. albidus. This feature corresponds to C=O stretching in carboxylic groups (COOH) [29,30,33], and aligns with the HPLC-PDA-ESI-MS^n^ data, which revealed a relatively higher abundance of ellagic acid derivatives in C. salviifolius and C. ladanifer. Furthermore, this relatively higher abundance of ellagic acid derivatives in the HEs of C. salviifolius and C. ladanifer, compared with C. albidus, is also consistent with the more intense FTIR-ATR signals observed at band d (ν(C=O) in COOH), band e (ν(C=C)), and band f (ν(C-C) aromatic). These bands reflect the presence of aromatic ring systems and are consistent with a higher phenolic acid content in C. salviifolius when compared with C. ladanifer and C. albidus.
Additionally, these findings were further supported by the TPC assay based on GAE, where the highest results were observed for this species, as well as the results from the TFC, CUPRAC, and FRAP assays, altogether indicating C. salviifolius as the most phenolics-rich extract and the extract with the greatest metal reduction capacity. Indeed, the study of plant-based extracts rich in phenolic acids has been reported to strongly and positively correlate with antioxidant activity [45], thus reinforcing the significance of these results in Cistus HEs. Moreover, the structural–activity relationship behind this observation results from the fact that a higher abundance of COOH groups augments the radical-scavenging potential of phenolic OH groups, ultimately promoting the antioxidant effect of phenolic acids [46]. In contrast, the HE from C. ladanifer is apparently more influenced by non-aromatic compounds, such as polysaccharides and aliphatic structures. Indeed, the nature of this spectral profile agrees with the comparatively weaker antioxidant activity observed in C. ladanifer, particularly in cell-free assays. Furthermore, noticeable differences between Cistus spp. HEs were observed in the DPPH^•^ and ABTS^•+^ inhibition assays, which may be the result of the differential scavenging activity demonstrated by phenolics in these HEs upon DPPH^•^ and ABTS^•+^ inhibition [47]. In fact, extracts from C. salviifolius have been presenting better antioxidant potential in comparison to other Cistus species [48,49].
The predicted cell-free antioxidant activity and the differences observed in the FTIR-ATR spectra of Cistus HEs demanded an in-depth analysis of their phenolic composition through HPLC-PDA-ESI-MS^n^. As mentioned before, the three studied Cistus species are markedly composed of ellagic acid derivatives in their composition. Structurally, ellagic acid is a dimeric form of gallic acid, exhibiting two lactones, a hydrophobic part made of two hydrocarbon rings and a hydrophilic moiety constituted by four OH groups [4,50]. This phenolic acid is a naturally occurring compound present in numerous medicinal plants and food crops [4], appearing either as complex water-soluble ellagitannins or as ellagic acid itself and derivatives [51]. On the other hand, the presence of flavan-3-ols, such as catechins and gallocatechins, was confirmed in C. albidus, despite their absence in C. ladanifer and C. salviifolius. This is coherent with the work of [13], and assumed to be a chemotaxonomic pattern, since plants from the subgenus Cistus, where C. albidus belongs, are particularly rich in flavonoids and subgroups like the flavan-3-ols. In comparison, C. ladanifer and C. salviifolius, both part of the Leucocistus subgenus, are mostly rich in ellagic acid derivatives [9], as corroborated by our HPLC-DAD-ESI-MS^n^ analysis. Nevertheless, the work of [8], previously identified flavan-3-ols in methanolic extracts of C. ladanifer. Besides flavan-3-ols, C. albidus and C. salviifolius were characterized by a considerable variety of myricetin and quercetin derivatives, and also some kaempferol-derived glycosides. Altogether, these compounds have been suggested as powerful antioxidants [49], once the higher frequency of OH groups in the B-ring of the aglycone core enhances their antioxidant activity [52]. Also, in the HE from C. salviifolius, we quantified an appreciable amount of ligstroside, an iridoid glycoside previously identified in this species [42], which is associated with the antioxidant capacity of several natural matrices like olives [53]. From this phenolic profile, it is conceivable that the observed cell-free antioxidant results found for C. salviifolius must be a mixed contribution of flavonol derivatives, ellagic acid derivatives, and ligstroside. On the other hand, C. ladanifer extract was shown to be poor in quercetin and myricetin derivatives, in agreement with previous chemical characterizations [38].
Surprisingly, in these Cistus spp. HEs, despite the predominance of ellagic acid derivatives like terflavin A and cistusin, other ubiquitously found ellagitannins such as punicalin and punicalagin [9,13,38,54] were not identified. This is possibly due to differences in the used solvents and extraction methodologies, harvesting period, or even edaphoclimatic and geographical influences, relevant to the Côa Valley (Portugal) microclimate from where these plants were harvested. Ellagitannins, classified as hydrolysable tannins, are primarily derived from gallotannins and are by far the largest and most complex family of tannins [55]. These molecules are built from the oxidative C-C coupling between at least two adjacent galloyl units, resulting in the formation of hexahydroxydiphenoyl (HHDP) moieties esterified to a glycosidic residue [51,55]. As such, terflavin A originally identified in the leaves of Terminallia catttapa L. (Combretaceae) by Tanaka and collaborators [56], alongside with terflavin B, were firstly hypothesized to be important biosynthetic precursors of punicalin and punicalagin. Further evidence about the biogenesis of these compounds was demonstrated in Terminallia chebula Retz. [57]. Meanwhile, the chemistry of cistusin, isolated from the leaves’ extracts of Cistus × incanus L. [36], was only recently elucidated as being structurally similar to terflavin A and punicalagin. Bearing in mind the structural similarity and biosynthetic relationships between these ellagitannins, it is conceivable that their properties may align with what is known for compounds like punicalin and punicalagin, or ellagic acid itself. Most of the bioactive potential of these molecules arises from the presence of four OH groups and lactone systems that enable these compounds to counteract both RNS and ROS [50]. While the bioactive potential of punicalagin and punicalin [51] has been widely studied, little is still known about the bioactive potential of ellagitannins like terflavin A and cistusin present in the genus Cistus [36].
According to ISO 10993-5:2009 guidelines [58] and OECD recommendations for cytotoxicity assays, concentrations that maintain ≥70% cell viability do not exhibit cytotoxic effects. However, to ensure that the bioactive effects evaluated on cells were not confounded with cytotoxic effects, we deliberately selected concentrations that exhibited no detectable toxicity and that resembled the control conditions. Therefore, from our cytotoxic assessment, C. ladanifer was the only extract with no significant deleterious effect on both cells’ metabolic activity and mass. Indeed, the work carried out by Andrade and collaborators [59] found that extracts from C. ladanifer promoted metabolic performance in a cell line of skin fibroblasts. In turn, C. albidus HE was the most toxic one, possibly given the presence of flavan-3-ols like gallocatechin and (+)-catechin, which were otherwise absent in C. salviifolius and C. ladanifer. Interestingly, the synergistic effect of catechins has been assumed as a possible mechanism that explains the antiproliferative effect in tumor cell lines [60]. From this perspective, it is plausible to assume that catechins may play a role in the higher toxicity of C. albidus in normal skin cells. Lastly, the work of Guzelmeric and collaborators [48] reported that HEs of C. salviifolius significantly impaired the growth of both 2D and 3D models of human pancreatic cancer but did not significantly affect the growth of human dermal fibroblasts. Noteworthy, and in line with what is further discussed, the potential of C. salviifolius and C. ladanifer extracts at their highest non-toxic concentrations (0.4 mg/mL and 1 mg/mL, respectively) was evaluated to ensure their biocompatibility for human skin application. Importantly, both extracts did not induce skin irritation, as demonstrated following OECD Test Guidelines, thereby confirming their safety profile.
Skin fibroblasts are the principal cell type in the dermal layer, having a fundamental role in the production of the extracellular matrix, such as fibronectin, collagen, elastin, and glycosaminoglycans. These are key components that maintain the skin’s elasticity and hydration [61]. However, given the permanent exposure of the skin to environmental hazards like UV radiation and pollutants, an imbalance may result in damaged proteins, lipids, and DNA, which contribute to the development of aging-related stressed skin [4]. As such, the assessment of the antioxidant protective effect of bioactive compounds in skin fibroblasts has been successfully employed by subjecting these cells to in vitro treatment with oxidative stressors like t-BHP and H_2_O_2_ [25,43]. From our results, C. ladanifer and C. salviifolius stood out for their cytoprotective capacity and as cellular oxidative modulators in NHDF cells. As previously discussed, since ellagic acid derivatives were commonly identified as major compounds in both species, they are likely to be responsible for the identified pharmacological activities. As such, an extract from T. catappa showed cytoprotective effects and reduced ROS levels by 23.1% in H_2_O_2_-stimulated human skin fibroblasts (Hs68 cell line) [62]. In turn, Dudonné and collaborators [63], using DNA macroarrays in a model of replicative senescence in NHDF cells, demonstrated that plant extracts rich in ellagitannins, such as castalagin and vescalagin, were able to upregulate the expression of catalase, a key antioxidant enzyme involved in the coordinated reduction and neutralization of reactive oxygen species (ROS) alongside other cellular antioxidant defenses. Another investigation proved the cytoprotective effect of the ellagitannin punicalagin in skin fibroblasts affected by UVA radiation by augmenting cells’ viability and reducing ROS levels generated by photooxidative stress [61]. Also related to this, the protective effect of a standardized pomegranate extract rich in ellagitannins was elucidated previously in UVA- and UVB-stimulated SKU-1064 human skin fibroblasts [64].
For instance, the potential anti-inflammatory effect of our extracts was investigated using LPS-stimulated RAW 264.7 macrophages, which is a well-known in vitro model of inflammation. When external LPS binds to the toll-like receptor 4 (TLR4) at the macrophages’ surfaces, it promotes the translocation of the Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) into the nucleus [24,65], inducing the expression of an arsenal of pro-inflammatory mediators and cytokines, including nitric oxide (NO), prostaglandin E2 (PGE2), tumor necrosis factor (TNF-α), IL-1β, and IL-6 [66]. Our results revealed the remarkable ability of C. ladanifer HE to inhibit NO production, indicating significant anti-inflammatory potential. In contrast, C. salviifolius and C. albidus HEs did not exhibit any notable anti-inflammatory activity. Interestingly, the ethanol-based extract of the stem bark of T. catappa, essentially composed of ellagic acid derivatives, like terflavin A and castalagin isomers, was demonstrated to reduce IL-1β levels and nitrites production in LPS-stimulated RAW 264.7 macrophages [67]. Another study with this plant species investigated a methanolic extract particularly rich in the ellagitannin α-punicalagin, showing that it reduces the LPS-induced production of NO and other inflammatory markers, besides an evident reduction in ROS levels [68]. Similarly, punicalin and ellagic acid from pomegranate peels were shown to reduce nitrite levels in LPS-activated macrophages, as well as a series of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6) [69]. Interestingly, the chemical structure of ellagitannins present in pomegranate peels, and the HHDP moieties, flavogallonyl and/or gallagyl parts, were evidenced as fundamental functional groups involved in the inhibition of NO production [70]. Overall, these studies are particularly relevant, reinforcing the assumption that the identified ellagitannins, terflavin A and cistusin, are fundamental for the inhibition of cellular nitrite production promoted by the C. ladanifer’s HE herein demonstrated.
Additionally, senescent fibroblast cells are known to exhibit several key hallmarks, from which increased lysosomal β-galactosidase activity stands out [24,71]. According to our work, C. ladanifer presents greater potential over C. salviifolius and C. albidus, and the HE of C. ladanifer was tested for cell senescence and shown to counteract the etoposide-induced senescence of NHDFs. Several recent works showed the potential anti-senescence effect of natural products in fibroblasts, such as phenolic-rich extracts of E. globulus [24] and Thymbra capitata (L.) Cav. [71]. Furthermore, skin aging is not only influenced by ROS and RNS, but also by the dysregulation of multiple enzymatic activities. This enzymatic imbalance contributes to reduced skin moisture, hyperpigmentation, a loss of elasticity, and wrinkle formation [72]. Bearing in mind these aspects, C. salviifolius stood out for its enzymatic inhibitory activity, steadily inhibiting all the tested enzymes at rates comparable to those of the positive controls. Alongside this, the HE of C. ladanifer also presented a remarkable potential to inhibit elastase. Previous works have suggested that ellagitannins, due to their glucosyl moiety connected to HHDP moieties, interact with the elastase’s aromatic side chains [73]. As discussed before, the extracts content in terflavin A and cistusin are apparently major drivers of the elastase inhibitor role played by HEs of C. salviifolius and C. ladanifer. In fact, a recent investigation testing extracts from Cytinus hypocistis (L.) L. confirmed that hydrolysable tannins like ellagic acid derivatives are responsible for the observed anti-aging properties, namely those related to elastase inhibition [74]. In our work, although C. ladanifer HE exhibited comparatively lower activity (~30% inhibition), it was still identified as a potential inhibitor of tyrosinase. On the other hand, C. albidus presented promising inhibition potential against tyrosinase and elastase, inhibiting around 50% of both enzymes. The HEs of C. ladanifer and C. albidus showed no inhibitory potential against hyaluronidase. Based on these cell-free anti-aging findings, C. salviifolius, followed by C. ladanifer, emerge as the most promising candidates for the development of advanced formulations containing bioactive ingredients that target skin remodeling enzymes.
Based on the provided phenolic profile and compound quantification (mg/g extract), the tested non-toxic extracts’ solutions delivered significantly different amounts of major ellagitannins to cells, which translated into different cytoprotective, antioxidant, and anti-inflammatory potentials. Specifically, C. ladanifer HE (1 mg/mL) provides 6.4 µg/mL of terflavin A and 4.7 µg/mL of cistusin. Meanwhile, C. salviifolius (0.4 mg/mL) delivers 7.9 µg/mL of terflavin A and 1.7 µg/mL of cistusin. The differences become even more pronounced for C. albidus HE, where a 0.2 mg/mL extract solution results in approximately 0.5 µg/mL of both terflavin A and cistusin being administered to cells. It is therefore plausible to hypothesize that these quantitative differences in ellagitannin content per extract solution applied to macrophages are one of the major reasons behind the exclusively anti-inflammatory role of C. ladanifer HE, as well as for its antioxidant and cytoprotective activities in NHDFs. The authors point out that although the three studied HEs share similar qualitative profiles, their quantitative differences result in distinct biological inputs at the cellular level. In particular, the higher delivery of ellagitannins by C. ladanifer HE, allied to its lower cytotoxicity and combined with its specific phytochemical ratios, may contribute to its stronger and more consistent activity at the cellular level. Conversely, the lower amounts of these key compounds in the C. albidus HE may limit its efficacy despite the superficial compositional similarities. These findings underscore the importance of considering not only qualitative composition but also the effective concentrations of individual bioactive constituents that reach cells when interpreting extract bioactivity.
Lastly, the authors recognize the limitations of this study that pave the way for future work. Firstly, the biological activities were evaluated only in vitro, which limits extrapolation to the complexity of human skin. In light if this, confirmation in ex vivo or in vivo models is needed. Secondly, the use of extracts prevented the identification of the specific compounds driving each bioactivity, highlighting the need for the fractionation or targeted isolation of major compounds, specifically the ellagitannins identified. Also, mechanistic pathways underlying the antioxidant, anti-inflammatory, and anti-senescence effects were not explored, and should be addressed in the next step through molecular-level investigations. Finally, broader toxicological and formulation-related assessments are required to substantiate the anti-aging potential for human skin of the HE of C. ladanifer, before translational application into the cosmetics industry.
5. Conclusions
This study revealed the potential of plants from the genus Cistus as candidates for the obtention of natural bioactive compounds targeting skin aging. Among the tested extracts, C. ladanifer, notably rich in ellagitannins, emerged as the most promising one. Its potent antioxidant and cytoprotective activity, potential anti-inflammatory effect, and remarkable anti-senescence activity, in combination with significant anti-enzymatic effects, underscore its suitability as a candidate to include in anti-aging cosmetics. Importantly, the absence of skin irritation in OECD-compliant tests supports the safety profile of such natural extracts, namely C. ladanifer, for future topical applications. By bridging modern phytochemical and pharmacological evidence, these findings offer direct relevance to the industry, particularly in the context of growing consumer demand for natural, eco-conscious, and scientifically substantiated skincare solutions. Lastly, the HE from C. ladanifer is highlighted as a promising candidate for inclusion in innovative skin anti-aging formulations in the cosmetics industry.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Michalak M. Plant Extracts as Skin Care and Therapeutic Agents Int. J. Mol. Sci.2023241544410.3390/ijms 24201544437895122 PMC 10607442 · doi ↗ · pubmed ↗
- 2Khavkin J. Ellis D.A.F. Aging Skin: Histology, Physiology, and Pathology Facial Plast. Surg. Clin. N. Am.20111922923410.1016/j.fsc.2011.04.00321763983 · doi ↗ · pubmed ↗
- 3Costa E.F. Magalhães W.V. Di Stasi L.C. Recent Advances in Herbal-Derived Products with Skin Anti-Aging Properties and Cosmetic Applications Molecules 202227751810.3390/molecules 2721751836364354 PMC 9658815 · doi ↗ · pubmed ↗
- 4Alfei S. Marengo B. Zuccari G. Oxidative Stress, Antioxidant Capabilities, and Bioavailability: Ellagic Acid or Urolithins?Antioxidants 2020970710.3390/antiox 908070732759749 PMC 7465258 · doi ↗ · pubmed ↗
- 5MateșL. Banc R. Zaharie F.A. Rusu M.E. Popa D.S. Mechanistic Insights into the Biological Effects and Antioxidant Activity of Walnut (Juglans regia L.) Ellagitannins: A Systematic Review Antioxidants 20241397410.3390/antiox 1308097439199220 PMC 11351988 · doi ↗ · pubmed ↗
- 6Liu H.M. Cheng M.Y. Xun M.H. Zhao Z.W. Zhang Y. Tang W. Cheng J. Ni J. Wang W. Possible Mechanisms of Oxidative Stress-Induced Skin Cellular Senescence, Inflammation, and Cancer and the Therapeutic Potential of Plant Polyphenols Int. J. Mol. Sci.202324375510.3390/ijms 2404375536835162 PMC 9962998 · doi ↗ · pubmed ↗
- 7Zalegh I. Akssira M. Bourhia M. Mellouki F. Rhallabi N. Salamatullah A.M. Alkaltham M.S. Khalil Alyahya H. Mhand R.A. A Review on Cistus sp.: Phytochemical and Antimicrobial Activities Plants 202110121410.3390/plants 1006121434203720 PMC 8232106 · doi ↗ · pubmed ↗
- 8Gaweł-Bęben K. Kukula-Koch W. Hoian U. Czop M. Strzępek-Gomółka M. Antosiewicz B. Characterization of Cistus × incanus L. and Cistus ladanifer L. Extracts as Potential Multifunctional Antioxidant Ingredients for Skin Protecting Cosmetics Antioxidants 2020920210.3390/antiox 903020232121584 PMC 7139296 · doi ↗ · pubmed ↗