Factors Influencing the Production Efficiency of Cloned Pigs: A Large-Scale Retrospective Analysis
Huaxing Zhao, Shouquan Zhang, Xiaopeng Tang, Rong Zhou, Ranbiao Mai, Lvhua Luo, Qiaoyun Su, Sixiu Huang, Zhenfang Wu, Zicong Li, Gengyuan Cai, Junsong Shi

TL;DR
This study identifies key factors affecting pig cloning success, including donor breed, season, and embryo numbers, offering practical guidance to improve cloning efficiency.
Contribution
The study provides new insights into breed-specific effects and optimal embryo transfer numbers for pig cloning based on a large-scale retrospective analysis.
Findings
Pietrain and Duroc donor cells yield higher cloning efficiency compared to Large White and Yorkshire.
Transferring 100–150 embryos per surrogate optimizes cloning efficiency without compromising pregnancy or delivery rates.
Summer is the least favorable season for embryo transfer, with lower success rates compared to other seasons.
Abstract
Pig cloning is an important technology, but its low efficiency limits wider application. By analyzing five years of data involving over 2000 surrogate sows and 367,000 embryos, we identified several important factors affecting cloning success. We found that the breed of the donor cell significantly influences outcomes, with notable differences observed between breeds. Compared to Summer, the other three seasons are more suitable for embryo transfer. Interestingly, using fewer embryos (100–150 per surrogate) resulted in better efficiency than using larger numbers. This contrasts with recommendations from a decade ago, suggesting that optimal embryo numbers need continual adjustment as cloning techniques improve. Our findings provide clear, practical guidance to improve the production of cloned pigs for agriculture and biomedical research. Pig somatic cell nuclear transfer (SCNT) has…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
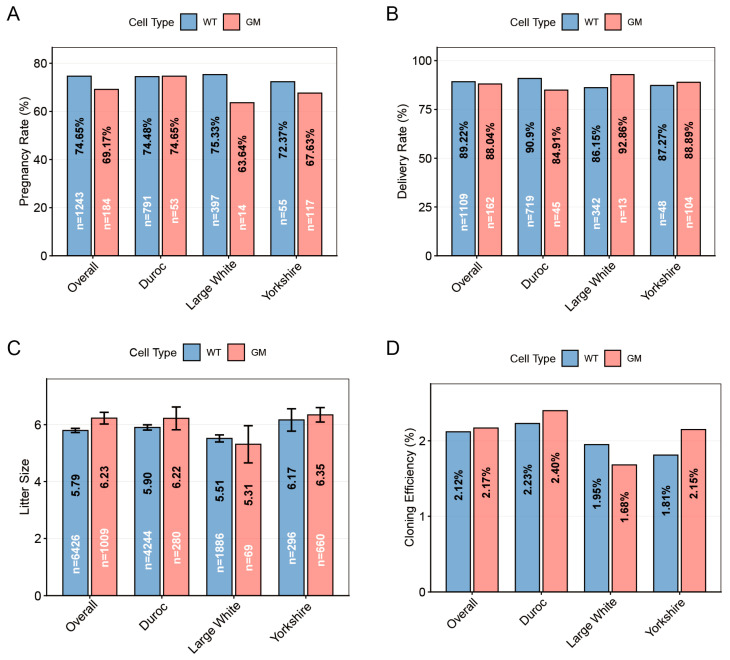 Figure 1
Figure 1- —National Major Agricultural Science and Technology Project
- —Department of Agriculture and Rural Affairs of Guangdong Province, China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReproductive Biology and Fertility · Pluripotent Stem Cells Research · Animal Genetics and Reproduction
1. Introduction
Somatic cell nuclear transfer (SCNT), or cloning, in pigs has valuable applications in agriculture [1], biomedicine [2,3], and life sciences [4,5]. Since the first batch of SCNT pigs was generated in 2000 [6], this technique has been extended to generate a population of elaborately selected boars with superior production performance [7,8] as well as genetically modified (GM) pigs. As recently as ten years ago, the efficiency of developing SCNT pig embryos to birth was only about 1% [9,10].
Animal cloning involves injecting a single donor cell into an enucleated mature oocyte, generating reconstructed embryos after electro-fusion, and transferring them into the uterus of the recipient animal. It is evident that the efficiency of animal cloning is influenced by factors related not only to oocytes, donor cells, embryo, and recipient animals, but also to other procedural and environmental variables. As previously reported, several efforts have been undertaken to improve porcine cloning, including the optimization of oocyte quality [11,12], donor cell preparation [13,14], the adjustment of fusion and activation [15,16], embryo transfer protocol [17], and the ovulation status of the recipient [17,18]. These studies have illustrated that optimizing the SCNT and embryo transfer procedures is necessary to improve the porcine cloning efficiency. Moreover, the number of embryos transferred is an important factor for porcine cloning efficiency. Although sows normally need only a minimum of four normal embryos to maintain pregnancy [19], because of the poor quality of SCNT embryos, people have generally transferred a large number of SCNT embryos to maintain pregnancy. With optimization procedures, the quality of SCNT embryos has been improved; thus, the number of embryos transferred needs to be re-evaluated.
Therefore, we conducted this retrospective study using data from cloned pig production in our laboratory to provide new insights for the pig SCNT field. In the present study, we investigated the influence of multiple factors on porcine cloning efficiency. We analyzed large-scale data from five years of cloned pig production, encompassing the transfer of 367,701 SCNT embryos into 2019 recipients, which resulted in 7781 cloned pigs. This dataset allowed us to evaluate the impact of season, donor cell breed, and the number of embryos transferred on key outcomes. The goal was to establish a more efficient procedure for large-scale production of pig cloning by evaluating the impact of the variable factors on pregnancy, delivery, and cloning efficiency.
2. Materials and Methods
2.1. Animals
All animal experiments in this study were approved by the Institutional Animal Care and Use Committee, South China Agricultural University. All efforts were made to minimize animal suffering.
2.2. Study Design and Data Collection
This retrospective study analyzed data from the large-scale production of cloned pigs at our facility from November 2021 to July 2025. Seasons for analysis were defined as follows: Spring (March–May), Summer (June–August), Autumn (September–November), and Winter (December–February). All embryo transfer procedures were conducted in Guangdong Province, China.
2.3. Oocyte Collection and Maturation
The ovaries of gilts were obtained from a local slaughterhouse and transported to the laboratory in normal saline with the antibiotics penicillin-G (100 IU/mL) and streptomycin sulfate (100 mg/L) at 30–35 °C within 5 h. Using a 10 mL disposable syringe with an 18-gauge needle, follicular fluid with cumulus–oocyte complexes (COCs) was aspirated from antral follicles (3–6 mm diameter), transferred to a 50 mL centrifuge tube, and allowed to stand for 20 min at 38.5 °C to aspirate the supernatant. Porcine COCs with at least three layers of compact cumulus cells and homogenous cytoplasm visible under a stereomicroscope were selected for in vitro maturation (IVM). The IVM medium is reported in our previous publication [20]. Approximately 50 COCs were transferred to each well of a four-well Nunc dish (Thermo Fisher Scientific, Waltham, MA, USA) into 500 μL of fresh IVM medium, and cultured in a humidified atmosphere at 38.5 °C with 5% CO_2_ for 44 h. Then, the surrounding cumulus cells were removed by treating the COCs with Dulbecco’s phosphate buffered saline (DPBS; Thermo Fisher Scientific, Waltham, MA, USA) containing 1 mg/mL hyaluronidase (Sigma-Aldrich, St. Louis, MO, USA) and 0.1% polyvinyl alcohol (Sigma-Aldrich, St. Louis, MO, USA) for 5 min at 38.5 °C by gentle pipetting for approximately 200 times. The oocytes were observed under a stereomicroscope, and those with the first polar body in the perivitelline space and intact cell membranes were selected for further cloning. Oocyte maturation culture was performed by the same core team following an identical protocol.
2.4. Preparation of Donor Cells
Donor cells were prepared as previously described [21,22]. Briefly, WT donor cells were obtained from an adult boar by ear biopsy. Ear tissues were immediately immersed in 75% ethanol, washed thrice with DPBS containing penicillin-G (100 IU/mL) and streptomycin sulfate (100 mg/L), and macerated into 1–2 mm pieces in Dulbecco’s Modified Eagle’s Medium (DMEM; Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Waltham, MA, USA). Then, fragments of ear tissue were seeded into the cell culture dishes and cultured in a humidified incubator with 5% CO_2_ at 37 °C; the culture medium was changed every other day. Fibroblast cells were harvested using 0.25% trypsin–EDTA (Waltham, MA, USA) and frozen in liquid nitrogen with the freezing medium (50% FBS, 40% DMEM, and 10% dimethyl sulfoxide). Primary fetus fibroblasts were isolated from 35-day-old male fetuses of Duroc, Large White, Yorkshire, and Bama pigs. Primary fetus fibroblasts were cultured in DMEM containing 12% FBS and 100 IU/mL penicillin-G/streptomycin in a humidified incubator with 5% CO_2_ at 39 °C. The transgene was mixed with a transposase, pCMV-hyPBase, and transfected into primary fetus fibroblasts by electroporation (BTX, San Diego, CA, USA). Then, the transfected cells were cultured in fresh culture medium. After 24 h, 300 μg/mL G418 (Gibco, Waltham, MA, USA) was added to the culture medium to select transfected cell colonies for about 15 days. The surviving cell colonies with EGFP expression were isolated within colony cylinders (Bellco Glass, Vineland, NJ, USA) and propagated in a fresh 24-well plate. Four colonies that proliferated well, with bright fluorescence, were then expanded and screened for the presence of the positive transgene.
Before SCNT, the fibroblasts were thawed and cultured in DMEM containing 10% FBS for 2–3 days until they reached 80–90% confluence. Adherent cells were harvested with 0.05% trypsin for 1 min and used for generating SCNT embryos.
2.5. Preparation of Somatic Cell Nuclear Transfer Embryos
Matured oocytes and a small number of donor cells were mixed in M199 medium (Gibco, Waltham, MA, USA) containing 2% FBS and 7.5 μg/mL cytochalasin B. The oocytes were enucleated as described in our previous publication [21]. Oocytes were stained with 1 μg/mL Hoechst 33,342, and using microinjection needle, the first polar body was aspirated along with approximately 15% of the adjacent cytoplasm under UV light irradiation. For subsequent SCNT, the absolutely enucleated oocytes were used. A single donor cell with a round and smooth surface was microinjected into the perivitelline space of each enucleated oocyte. The reconstructed oocytes were washed thrice in porcine zygote medium 3 (PZM-3) medium [23], and then electrically fused using two direct current pulses at 150 V/mm for 50 μs, and subsequently incubated in PZM-3 medium for 1 h at 38.5 °C in a humidified atmosphere of 5% CO_2_. The reconstructed oocytes were activated by two direct current pulses of 100 V/mm for 20 μs, and incubated in PZM-3 medium supplemented with 5 μg/mL cytochalasin B for 4 h. The activated embryos were finally cultured in fresh PZM-3 medium for 20 h at 38.5 °C in a humidified incubator with 5% CO_2_ and used for embryo transfer.
The embryo transfer was conducted as previously reported [17], but with some adjustments. In brief, SCNT embryos cultured for 20 h in vitro were examined and the abnormal embryos were removed. Only embryos with intact cytoplasm or normal cleavage were employed for embryo transfer. These normal SCNT embryos were loaded into a transparent transfer tube and placed in a portable incubator before transferring into surrogate sow. Estrus-synchronized Landrace and Yorkshire sows in parity 2–5 and showing natural standing estrus within 30–50 h were used as embryo recipients. These sows were anesthetized with ketamine (25 mg/kg body weight) and xylazine (1.1 mg/kg body weight) for induction and 3% isoflurane for maintenance. A single oviduct was exposed through surgery. The SCNT embryos with 100 μL of culture medium were placed directly using a syringe into the oviduct of the sow. One month after embryo transfer, the pregnancy status of the surrogate sows was monitored by ultrasound equipped with a convex transducer. If spontaneous farrowing did not occur until gestation day 116, the surrogate sows were injected with a prostaglandin analog (cloprostenol, 200 μg/surrogates); after about 24 h, they delivered vaginally under supervision or with assistance. The total number of newborn cloned piglets (including live-born and stillborn) in each litter was recorded to calculate the average litter size of farrowed surrogates. To ensure uniformity, all embryo preparation and transfer procedures were conducted by a single, dedicated team according to a uniform, established protocol.
2.6. Statistical Analysis
Statistical analyses were performed based on the structure of the outcome variables. Pregnancy rate, delivery rate, and overall cloning efficiency were presented as percentages. Comparisons of these rates between groups were performed using Pearson’s chi-square test. When the overall chi-square test was significant (p < 0.05), post hoc pairwise comparisons were conducted with Bonferroni correction to adjust for multiple comparisons. Comparisons of means between two independent groups were performed using Student’s t-test. All statistical analyses were performed using R software (version 4.5.1).
3. Results
3.1. Overview of Five-Year Large-Scale Cloning Production
Over the course of this large-scale production study, a total of 367,701 porcine SCNT embryos were transferred into 2019 surrogate sows in our laboratory from 2021 to 2025. Per surrogate, the average number of SCNT embryos transferred was 182 (range: 112–300). A total of 1488 surrogates (73.70%) became pregnant and 1316 surrogates (88.44%) delivered 7781 piglets, with an average cloning efficiency of 2.12% (Table 1). Donor cells were obtained from adult Duroc, Large White, Pietrain, Yorkshire, and GM pigs.
3.2. Effect of Embryo Transfer Season on Cloning Outcomes
The season in which surrogates received embryo transfer significantly influenced pregnancy maintenance, successful delivery, the number of piglets per litter, and the overall cloning efficiency. Regarding pregnancy rates, those for surrogates receiving embryo transfers in Summer were significantly lower than that those in Spring (67.90% vs. 76.62%, p = 0.0462), but did not differ significantly from Spring, Autumn (72.83%), and Winter (72.43%). A similar pattern was observed for the delivery rates. The delivery rate in Summer was significantly lower than in Spring (80.00% vs. 89.67%, p < 0.01), Autumn (80.00% vs. 89.39%, p = 0.044), and Winter (80.00% vs. 89.28%, p = 0.0346), and no significant differences were found among the latter three seasons. In addition, the overall cloning efficiency was significantly compromised in Summer compared to the other three seasons. The efficiencies in Spring (1.74% vs. 2.20%, p < 0.01), Autumn (1.74% vs. 2.05%, p < 0.01), and Winter (1.74% vs. 2.22%, p < 0.01) were statistically similar and markedly higher. Regarding the average litter size, Winter yielded the highest numerical value (6.23 ± 2.50), which was significantly greater than that in both Spring (5.76 ± 2.45) and Autumn (5.76 ± 2.46). The average litter size in Summer (6.11 ± 2.93) was not statistically different from that in any other season. Collectively, Summer conditions adversely affected cloning success by reducing pregnancy establishment, lowering maintenance, and diminishing the overall cloning efficiency. Outcomes in Spring, Autumn, and Winter were more favorable and generally comparable.
3.3. Influence of Donor Cell Breed on Cloning Efficiency
The breed of the donor cell was a significant factor affecting cloning outcomes, and this influence was consistently observed in both wild-type (WT) and genetically modified (GM) cell populations, which were analyzed separately (Table 2 and Table 3).
In the analysis of WT-donor cells, pregnancy rates showed no significant differences among breeds, ranging from 72.00% to 75.33%. Similarly, delivery rates showed no significant differences, despite numerical variations from 72.22% to 90.90% (Table 2). However, both the number of piglets per litter and cloning efficiency exhibited marked breed-specific variations. WT-Pietrain cells yielded a markedly higher number of piglets per litter (9.77 ± 3.09) and achieved the highest cloning efficiency (2.48%). This litter size was dramatically higher than that of WT-Duroc (9.77 ± 3.09 vs. 5.90 ± 2.49, p < 0.01), WT-Large White (9.77 ± 3.09 vs. 5.51 ± 2.29, p < 0.01), and WT-Yorkshire (9.77 ± 3.09 vs. 6.17 ± 2.73, p < 0.01) cells. WT-Duroc, WT-Large White, and WT-Yorkshire did not differ significantly from each other in litter size. In terms of cloning efficiency, WT-Pietrain achieved the highest efficiency (2.48%), and no significant difference was observed between WT-Pietrain and WT-Duroc (2.23%). Moreover, the cloning efficiency of WT-Duroc was significantly higher than both WT-Large White (2.23% vs. 1.95%, p < 0.001) and WT-Yorkshire (2.23% vs. 1.81%, p < 0.001). Similarly, WT-Pietrain showed significantly higher efficiency than both WT-Large White (2.48% vs. 1.81%, p < 0.001) and WT-Yorkshire (2.48% vs. 1.95%, p = 0.00341). No significant difference was observed between WT-Large White and WT-Yorkshire.
A similar pattern was observed with genetically modified (GM) donor cells, further supporting the significant impact of cell breed on cloning outcomes. Analysis revealed significant variations in cloning efficiency among GM breeds, while pregnancy rates, delivery rates, and the number of piglets per litter remained comparable (Table 3). Among the GM-donor cell breeds, pregnancy rates were highest with GM-Duroc (74.65%) and lowest with GM-Large White (63.64%), though these differences were not statistically significant. Similarly, for delivery rates, the highest value was observed with GM-Large White (92.86%) and the lowest with GM-Bama (74.42%), with no significant differences across groups. The litter size was largest for GM-Bama and smallest for GM-Large White (6.84 ± 2.93 vs. 5.31 ± 2.36, p = 0.745), but the difference was not statistically significant. However, cloning efficiency varied significantly among GM breeds (Table 3). GM-Duroc achieved the highest efficiency (2.40%), and no significant difference was observed between GM-Duroc and GM-Yorkshire (2.15%). The cloning efficiency of GM-Duroc was significantly higher than both GM-Large White (2.40% vs. 1.68%, p = 0.0492) and GM-Bama (2.40% vs. 1.73%, p = 0.00160). Additionally, GM-Yorkshire showed significantly higher efficiency than GM-Bama (2.15% vs. 1.73%, p = 0.0339).
These results indicated that donor cell breeds significantly affected cloning efficiency in analyses of both WT and GM donor cells.
3.4. Comparison of Cloning Efficiency Between WT and GM Donor Cells
To evaluate the effect of genetic modification on the cloning efficiency of pigs, we compared the developmental outcomes of SCNT embryos using WT and GM donor cells across three pig breeds: Duroc, Large White, and Yorkshire. As shown as Figure 1, no statistically significant differences were observed between the WT and GM groups in terms of the pregnancy rate, delivery rate, average litter size, or cloning efficiency. Notably, however, in the Large White breed, the GM group showed a consistent downward trend in both the pregnancy rate (75.33% vs. 63.64%, p = 0.323; Figure 1A) and cloning efficiency (1.95% vs. 1.68%, p = 0.237; Figure 1D) compared to the WT group, although these differences were not statistically significant.
To control for interannual variability and enable a direct comparison, we analyzed the production outcomes of WT and GM donor cells within each calendar year from 2022 to 2025 (Table S1). Embryos derived from each donor type were transferred into recipient sows, and key reproductive parameters were evaluated on an annual basis. Pregnancy rates in recipients receiving WT-derived embryos ranged from 70.69% to 93.53%, while those receiving GM-derived embryos ranged from 64.10% to 92.31%. No statistically significant differences between donor cell types were observed in any individual year (all p > 0.05). Similarly, delivery rates remained high and comparable between groups throughout the study period. The total cloning efficiency showed a consistent upward trend from 2022 to 2025 for both WT (from 1.72% to 2.41%) and GM (from 1.87% to 3.04%) donor cells. The average litter size was similar between groups in most years, with a statistically significant difference observed only in 2024, where GM donor cells yielded a higher average litter size than WT (6.11 ± 2.47 vs. 5.47 ± 2.29 piglets, p = 0.031). In other years, litter sizes did not differ significantly between donor types. Collectively, these within-year comparisons indicate that genetic modification of donor cells did not adversely affect annual cloning efficiency or most reproductive outcomes in recipients.
3.5. Transferred Embryo Number Significantly Affects Cloning Efficiency
According to previous reports, the production efficiency of cloned pigs is influenced by the number of SCNT embryos transferred. For this analysis, we categorized the number of transferred embryos into four groups: 100–150 embryos, 151–200 embryos, 201–250 embryos, and 251–300 embryos. As shown in Table 4, the number of embryos transferred per surrogate had minimal impact on pregnancy rates, delivery rates, and average litter size across the different transfer groups. Statistical analysis confirmed no significant differences in pregnancy rates, delivery rates, and average litter size across the transferred embryo number groups. However, a marked and statistically significant inverse relationship was observed between the transferred embryo number into surrogates and cloning efficiency (p < 0.01). Specifically, the cloning efficiency achieved with 100–150 transferred embryos was significantly higher than that of the 151–200 transferred-embryo group (2.88% vs. 1.91%, p < 0.01), the 201–250 transferred-embryo group (2.88% vs. 1.80%, p < 0.01), and the 251–300 transferred-embryo group (2.88% vs. 1.53%, p < 0.01). Furthermore, the cloning efficiency of the 151–200 transferred-embryo group was significantly higher than that of both the 201–250 (1.91% vs. 1.80%, p < 0.01) and 251–300 transferred-embryo groups (1.91% vs. 1.53%, p < 0.01), while no significant difference was observed between the 201–250 and 251–300 transferred-embryo groups (p = 0.165).
To further verify the effects of the number of transferred embryos on the production efficiency of cloned pigs, we randomly transferred approximately 150 or 200 cloned embryos generated from the same donor cells and constructed on the same day to surrogate sows, respectively. This key finding was robustly validated by the controlled experiment, presented in Table 5. Although increasing the number of transferred embryos from 150 to 200 showed a trend toward reduced pregnancy rates (86.67% vs. 60.00%, p = 0.0986), this difference did not reach statistical significance. Recipients receiving 150 embryos exhibited significantly larger average litter sizes (6.38 ± 3.04 vs. 4.22 ± 1.39, p = 0.0377), and a nearly threefold greater cloning efficiency (3.66% vs. 1.27%, p < 0.01) compared to those receiving 200 embryos.
These results demonstrate that the number of embryos transferred to surrogates significantly affected the efficiency of cloned pig production. Specifically, transferring 150 embryos yielded optimal outcomes, with significantly higher cloning efficiency and larger litter sizes compared to 200 embryo transfers, while maintaining comparable pregnancy and delivery rates. This finding provides important guidance for optimizing SCNT protocols in pig cloning.
4. Discussion
SCNT technology is currently the most efficient and precise approach to generate genetically engineered pig models for agricultural applications and biomedical research [1,2,3], yet its overall cloning efficiency is constrained by multiple interacting factors. In this study, we reported that the season of transferred embryos, breeds of donor cells, and number of embryos transferred can affect the production of cloned pigs. These findings have immediate implications for enhancing the scale, reproducibility, and economic viability of pig cloning in both biomedical research and agricultural applications.
In this study, surrogate sows receiving embryo transfers during Summer exhibited the lowest pregnancy, delivery rate, and cloning efficiency rates compared to the other seasons. In contrast, the performance metrics for Spring, Autumn, and Winter transfers were largely comparable, with Spring showing only a slight, non-significant advantage. The significantly compromised pregnancy and delivery rates in surrogate sows receiving embryo transfers in Summer indicate a substantive failure in the establishment and maintenance of early pregnancy. Environmental heat stress can impair oocyte quality [24,25], which adversely affects the subsequent developmental potential of the SCNT embryos. Furthermore, in both gilts and sows, sustained heat stress and a long photoperiod during Summer are critical factors that not only suppress feed intake but also disrupt the imbalance of the hypothalamic–hypophysial–ovarian axis [26,27], thereby adversely affecting pregnancy maintenance. These impairments may therefore underlie the substantially lower cloning efficiency in surrogate sows that received embryo transfer in Summer. Although previous studies have established that the season of embryo transfer is an important factor for cloning efficiency in surrogate sows, not all studies have reported Summer as the most detrimental period [10,28,29]. These discrepancies are likely attributable to geographic variations in specific climatic conditions.
Donor cells are a critical determinant of cloning efficiency, with previous studies demonstrating the importance of donor cell characteristics, including size [13], source [29,30], type [31] and other factors [32,33]. Among WT donor cells, Pietrain and Duroc donor cell-derived SCNT embryos yielded the highest total cloning efficiency, while Large White and Yorkshire breeds showed lower performance. This trend persisted with GM donor cells, where GM-Duroc and GM-Yorkshire demonstrated better efficiency than GM-Large White and GM-Bama. These findings collectively suggest that specific pig breeds possess breed-specific characteristics that facilitate more effective nuclear reprogramming. Furthermore, we specifically investigated the impact of GM donor cells on cloning outcomes. Our large-scale data clearly indicate that the use of GM donor cells does not exert a significant negative impact on the efficiency of pig cloning compared to their wild-type counterparts. These findings collectively indicate that the genetic background of donor cells from different breeds directly impact the outcome of nuclear reprogramming during SCNT.
The number of embryos transferred is widely recognized as a key factor influencing the outcomes of pig cloning. In 2013, our team identified the transfer of 200–249 embryos per recipient as an optimal number range [31]. However, after more than a decade of large-scale data tracking, our current investigation refines this understanding. Our current analysis reveals that increasing the number of embryos transferred from 100–150 to 251–300 had no significant impact on the pregnancy rate, delivery rate, or average litter size of surrogate sows, but was negatively correlated with the overall cloning efficiency. This relationship is definitively supported by our controlled experiment demonstrating superior cloning efficiency and higher litter size in recipients receiving 150 versus 200 embryo transfers. These findings demonstrate that exceeding an optimal embryo number does not improve cloning outcomes, despite the established minimum requirement of four embryos for pregnancy maintenance in pigs. Notably, compared with data from a decade ago, pregnancy rates in the 200–299 embryo transfer groups have risen to exceed 70% (from below 60%), and cloning efficiency showed a >4-fold increase, indicating substantial improvements in SCNT embryo quality. However, excessive embryo development leads to mutual exclusion because the uterine capacity in sows limits the number of fetuses developing to term, resulting in the loss of supernumerary fetuses [34,35]. Thus, with the improved quality of individual cloned embryos, transferring excessive numbers becomes counterproductive for embryonic development to term. Our findings establish that surrogates receiving 100–150 cloned embryo transfers maintain comparable pregnancy rates, delivery rates, and average litter sizes while achieving significantly higher cloning efficiency than those receiving larger numbers of embryos.
This study retrospectively analyzed the embryo transfer records of our team rather than prospective new experiments. Thus, the experimental conditions, such as surrogate sows (including age, weight, parity, and breeds), passages of donor cell lines, the embryo transfer operation, the number of embryos transferred, and season, were not tightly controlled and their variations may have influenced the outcomes. Additionally, our data cannot determine whether transferring fewer than 100 embryos would lead to improved cloning outcomes in surrogate sows, which warrants further investigation. Furthermore, the lack of systematically recorded environmental data precludes a direct correlation between climatic conditions and the observed seasonal variations in cloning efficiency. Future studies integrating real-time environmental monitoring are warranted to address this. Moreover, our statistical approach, employing direct group comparisons, did not account for the simultaneous effects of multiple variables; thus, it cannot distinguish their individual and interactive influences on the outcomes. However, the total number of embryos transferred (367,701) and the number of surrogate sows (2019) were large, and data collected were for a longer duration. Therefore, this study provides valuable information for further research on porcine cloning.
5. Conclusions
Our large-scale retrospective analysis demonstrates that the genetic background of donor cells significantly influences the efficiency of pig cloning. SCNT embryos derived from Duroc and Pietrain donor cells yielded significantly higher production efficiency compared to those from Large White and Yorkshire breeds. Furthermore, our findings indicate that Spring is the optimal season for cloned pig production in South China, and transferring 100–150 embryos per surrogate sow represents the most efficient strategy to maximize overall cloning outcomes. These insights provide valuable guidance for optimizing large-scale SCNT protocols in swine.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Galli C. Lazzari G. 25th ANNIVERSARY OF CLONING BY SOMATIC-CELL NUCLEAR TRANSFER: Current applications of SCNT in advanced breeding and genome editing in livestock Reproduction 2021162 F 23F 323385243010.1530/REP-21-0006 · doi ↗ · pubmed ↗
- 2Dai Y. Vaught T.D. Boone J. Chen S.H. Phelps C.J. Ball S. Monahan J.A. Jobst P.M. Mc Creath K.J. Lamborn A.E. Targeted disruption of the alpha 1,3-galactosyltransferase gene in cloned pigs Nat. Biotechnol.20022025125510.1038/nbt 0302-25111875425 · doi ↗ · pubmed ↗
- 3Yan S. Tu Z.C. Liu Z.M. Fan N.N. Yang H.M. Yang S. Yang W.L. Zhao Y. Ouyang Z. Lai C.D. A Huntingtin Knockin Pig Model Recapitulates Features of Selective Neurodegeneration in Huntington’s Disease Cell 2018173989100210.1016/j.cell.2018.03.00529606351 PMC 5935586 · doi ↗ · pubmed ↗
- 4Niemann H. Lucas-Hahn A. Somatic cell nuclear transfer cloning: Practical applications and current legislation Reprod. Domest. Anim.20124721010.1111/j.1439-0531.2012.02121.x 22913555 · doi ↗ · pubmed ↗
- 5Park J.-K. Lee Y.-K. Lee P. Chung H.-J. Kim S. Lee H.-G. Seo M.-K. Han J.-H. Park C.-G. Kim H.-T. Recombinant human erythropoietin produced in milk of transgenic pigs J. Biotechnol.200612236237110.1016/j.jbiotec.2005.11.02116460825 · doi ↗ · pubmed ↗
- 6Onishi A. Iwamoto M. Akita T. Mikawa S. Takeda K. Awata T. Hanada H. Perry A.C.F. Pig cloning by microinjection of fetal fibroblast nuclei Science 20002891188119010.1126/science.289.5482.118810947985 · doi ↗ · pubmed ↗
- 7Vajta G. Gjerris M. Science and technology of farm animal cloning: State of the art Anim. Reprod. Sci.20069221123010.1016/j.anireprosci.2005.12.00116406426 · doi ↗ · pubmed ↗
- 8Liu T. Dou H. Xiang X. Li L. Li Y. Lin L. Pang X. Zhang Y. Chen Y. Luan J. Factors Determining the Efficiency of Porcine Somatic Cell Nuclear Transfer: Data Analysis with Over 200,000 Reconstructed Embryos Cell. Reprogram.20151746347110.1089/cell.2015.003726655078 PMC 4677548 · doi ↗ · pubmed ↗