Testing the Island Effect in a Highly Mobile Pollinator: Wing Morphological Divergence in Euglossa mixta from Continental and Insular Panama
Yostin Añino, Jordan Hernández-Martelo, Fernando Moya, Alejandro Piñeiro-González, Laura M. Pérez, Dumas Gálvez, Yosiat Vega-Rovira, Julio Trujillo, Anette Garrido, Danilo Arrocha, Franco Cruz-Jofré, Hugo A. Benítez
TL;DR
This study shows that even highly mobile orchid bees can develop subtle wing shape differences in island versus mainland environments.
Contribution
The study demonstrates that insular isolation can lead to fine-scale morphological divergence in a highly mobile pollinator species.
Findings
Wing shape differences in Euglossa mixta are more pronounced between distant islands and the mainland.
Geometric morphometrics reveal subtle but consistent morphological structuring in insular populations.
Results suggest that isolation can drive early morphological differentiation in highly mobile species.
Abstract
Islands provide unique opportunities to explore how isolation affects the lives of organisms. Although orchid bees are strong fliers, which leads to the assumption that their mobility prevents significant population differences, our study examined the wing shape of the orchid bee Euglossa mixta from the Coiba archipelago islands and a nearby mainland site in Panama. By using geometric morphometrics, we identified subtle yet consistent differences in wing shape across different sites. These variations were more pronounced between the distant islands and the mainland, indicating that even highly mobile pollinators can develop fine-scale morphological differences in insular environments. Islands provide valuable opportunities to study how isolation affects phenotypic variation. Even though orchid bees are highly mobile, their movement can still be restricted by marine barriers. In this…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
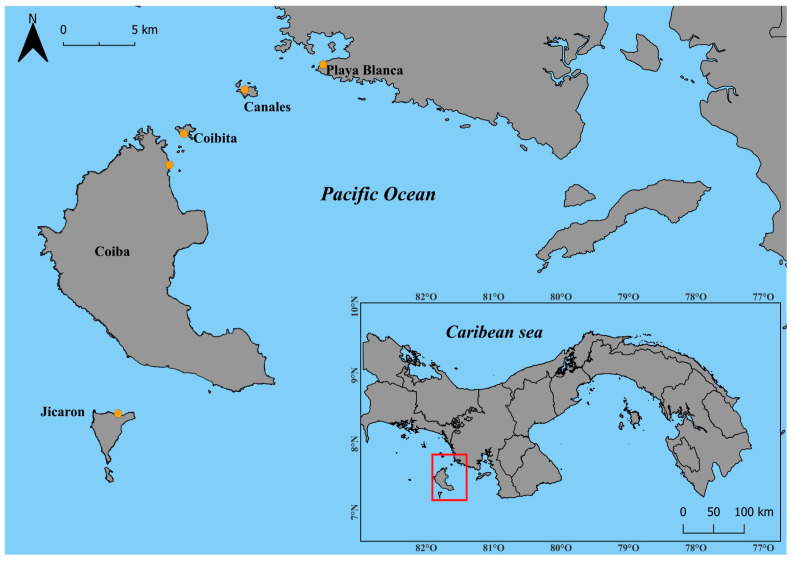 Figure 1
Figure 1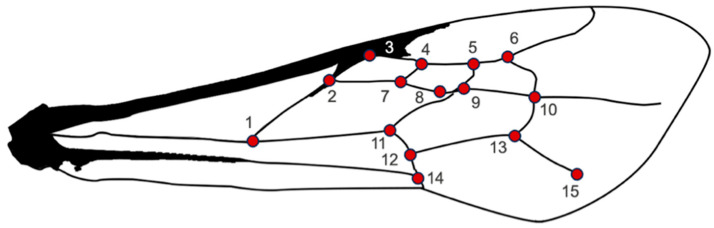 Figure 2
Figure 2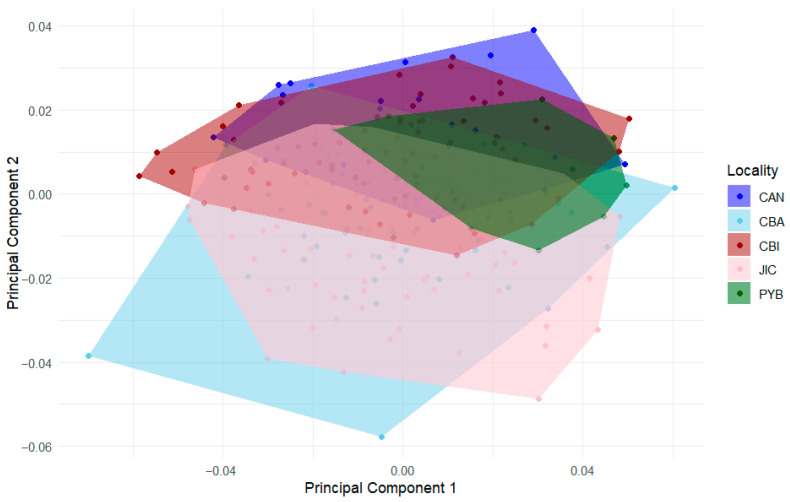 Figure 3
Figure 3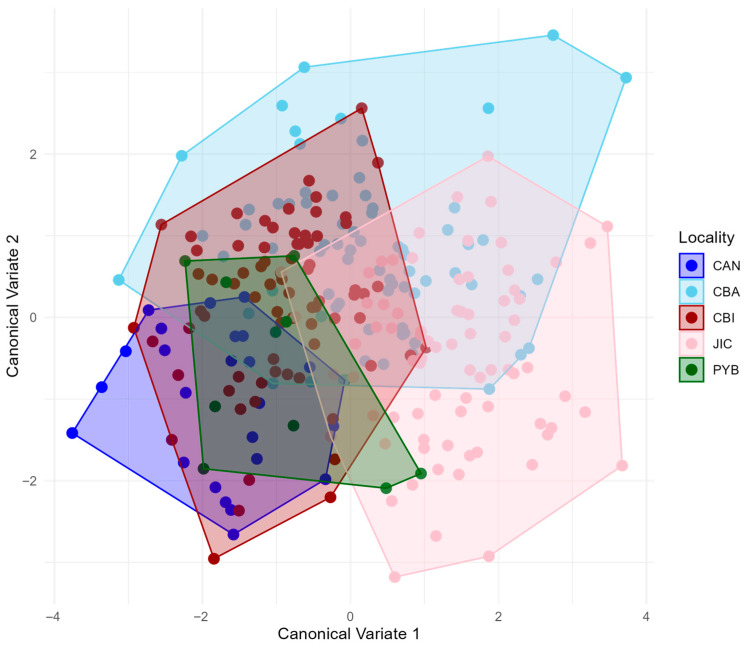 Figure 4
Figure 4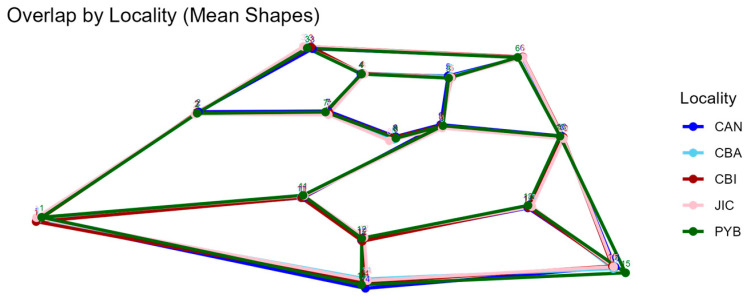 Figure 5
Figure 5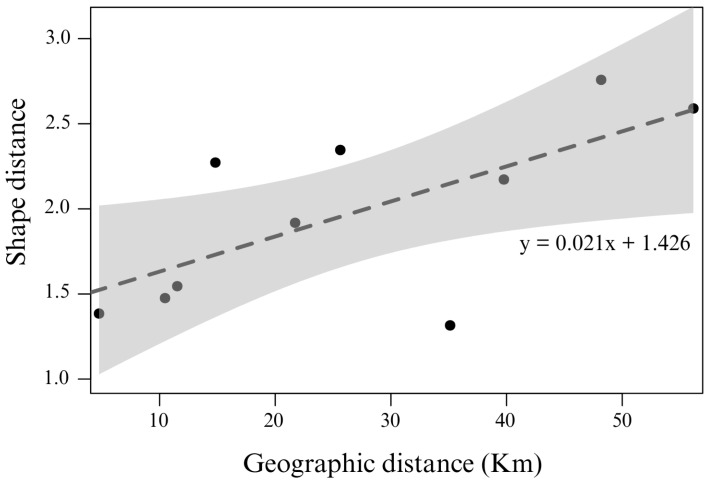 Figure 6
Figure 6- —Convocatoria Interna de Fondos de Investigación de la Estación Científica Coiba AIP
- —Convocatoria Nacional Subvención a Instalación en la Academia Convocatoria Año 2021
- —Millennium Institute BASE and ANID Becas/Magíster Nacional
- —Cape Horn International Center for Global Change Studies and Biocultural Conservation (CHIC)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMorphological variations and asymmetry · Plant and animal studies · Animal Behavior and Reproduction
1. Introduction
The morphological variation in organisms is influenced by both genetic factors and the environmental and ecological conditions under which their development occurs [1,2]. In particular, insular populations provide an ideal framework for studying these processes, as they represent a major source of evidence for the occurrence of rapid adaptive changes in morphological traits [3,4]. The inherent properties of islands, such as geographic isolation, small population sizes, and limited resource availability, among others, contribute to the isolation of evolutionary processes that promote local adaptation and, over time, speciation, a phenomenon commonly referred to as the island effect [5,6,7,8,9]. Consequently, populations inhabiting islands often exhibit phenotypic modifications compared to their continental counterpart, as a result of specific selective pressures associated with resource availability, intraspecific competition, dispersal constraints, or the absence of predators [10,11]. These changes may be expressed through variation in body size, wing proportions, locomotor structures, or feeding-related traits, reflecting the plasticity and adaptive response of species to insular environmental conditions [12,13].
Beyond their biogeographic isolation, islands function as natural laboratories for evolutionary experimentation, where selective pressures operate with greater intensity or independence than in continental systems [8,14]. As a result, insular environments provide valuable opportunities to observe the tempo and mode of evolutionary change, revealing how ecological constraints, resource limitation, and spatial isolation interact to shape phenotypic diversity [3,15]. In this sense, morphological variation serves as a key proxy for understanding the adaptive trajectories and diversification processes that characterize island biotas.
Although the particular conditions of islands may promote phenotypic divergence and the emergence of local adaptations, certain evolutionary processes can, in some cases, lead to the opposite effect by reducing genetic variability and, consequently, morphological variation [16,17]. Factors such as reduced gene flow, recent colonization events, founder effects, and genetic drift tend to limit diversity within insular populations [18,19]. Moreover, when islands are located close to one another or share similar environmental conditions, a certain degree of morphological homogenization may be maintained as a result of the occasional exchange of individuals or partial panmixia [20,21]. Taken together, these processes indicate that insularity does not always promote morphological differentiation; rather, its effect depends on the interaction between the evolutionary history of populations, the degree of connectivity among islands, and local selective pressures.
In this context, several studies have documented patterns of morphological variation across different taxa [22,23,24,25]; however, such processes have been scarcely explored in insular insects [7]. Insects represent a particularly suitable model for studying the effects of insularity on shape, given their wide distribution, high dispersal capacity, and sensitivity to local environmental conditions [7,26,27]. Among these, Euglossa mixta Friese, 1899 stands out as a valuable model species. This bee belongs to the tribe Euglossini, which includes approximately 200–250 species commonly known as orchid bees [28,29]. These bees are native to the Neotropics and are distinguished by their bright metallic coloration, their importance as bioindicators, and the behavior of males, which collect volatile chemical compounds from a wide range of sources, including orchids, but not limited to floral ones [9,30]. Their importance in pollination and their role as bioindicators of ecosystem health, as reflected by changes in composition and abundance, combined with the presence of populations both on the mainland and across oceanic island complexes, make E. mixta an ideal model for exploring how geographic isolation and environmental conditions shape morphology [31,32,33]. This species, therefore, provides an opportunity to test whether classical insular patterns, such as reduced variability or trait shifts associated with isolation, are also expressed in highly mobile pollinators. Because our goal is to test for subtle morphological responses to insularity, an analytical framework that captures fine-scale shape differences is required. In this context, geometric morphometrics provides an ideal analytical framework for assessing subtle morphological variation in E. mixta, as it allows the precise quantification and comparison of biological shapes. By capturing the geometry of anatomical structures through the placement of homologous landmarks, this approach enables the detection of fine-scale shape differences [34,35]. Wing venation variability is not uniform across Apoidea, with some tribes, such as Eucerini, exhibiting markedly higher morphological plasticity than Euglossini. In contrast, the relatively conserved wing architecture of orchid bees makes the detection of population-level shape differences particularly informative, as it reduces the likelihood that observed variation reflects taxon-specific instability rather than spatial structuring.
This methodology enables the capture of the complete geometry of structures, providing a detailed and statistically rigorous description of morphological variation patterns [36,37,38,39].
In insects, wing structures have been widely used in studies of population differentiation and adaptation, as they reflect both genetic components and phenotypic responses to environmental conditions [40]. For example, Ostwald et al. [7] evaluated variation in the wing venation patterns of the social bee Halictus tripartitus Cockerell, 1895, comparing an isolated population from Santa Cruz Island (California) with mainland populations, and found significant morphological differentiation between the two groups. Similarly, Laojun et al. [6] analyzed wing size and shape in the mosquito vectors Aedes albopictus (Skuse, 1894) and Armigeres subalbatus (Coquillett, 1898) across the Ranong and Trat archipelagos (Thailand), reporting significant differences in centroid size, used as a proxy for body size, and slight variations in wing shape. These studies highlight that wing morphology, even in closely related or geographically proximate populations, can reveal evolutionary responses to isolation and local adaptation.
Western Panama constitutes an exceptional natural laboratory for investigating the effects of insularity on morphological variation. This region is characterized by its high biological diversity, elevated endemism, and complex geographical configuration [41,42]. In particular, the Coiba Island National Park System, composed of Coiba Island and 38 smaller islands (including Coibita, Jicarón, and Canales) located in the Gulf of Chiriquí, provides a unique opportunity to evaluate processes of rapid morphological adaptation driven by gradients of isolation and environmental conditions that contrast with those of the mainland [9,43]. The archipelago presents gradients of humidity, temperature, and vegetation cover that may influence flight performance and resource availability, providing an environmental context in which subtle variation in wing shape could arise. Although the bee fauna of the Coiba archipelago has been increasingly documented in recent years, information on population-level structuring and morphological variation across islands remains limited, particularly for highly mobile pollinators. This context is especially relevant for species with high dispersal capacity, such as E. mixta, whose insular and continental populations allow for the exploration of the relationship between geographic isolation and phenotypic differentiation.
Euglossa mixta is a relatively large-bodied orchid bee with strong flight capacity and broad foraging ranges, traits that make it a conservative and biologically informative model for testing insular effects. If morphological differentiation is detectable in a species with high dispersal ability and constrained wing venation, such patterns are unlikely to reflect trivial isolation and instead point to subtle but consistent spatial structuring. Moreover, E. mixta is one of the few orchid bee species consistently recorded across both the mainland and multiple islands of the Coiba archipelago, allowing comparisons across a well-defined gradient of geographic isolation.
Within this framework, this study aims to quantify variation in the wing morphology of E. mixta between insular and continental populations to assess whether geographic isolation influences phenotypic differentiation. Using a geometric morphometric approach, it seeks to determine whether distance and insular isolation are reflected in detectable changes in wing shape, providing evidence for the role of insularity in shaping the morphological structure of this Neotropical species. Therefore, this study hypothesizes that greater geographic isolation is associated with detectable divergence in wing shape, reflecting the influence of insular conditions on morphological differentiation.
2. Materials and Methods
To quantify the morphological variation through shape analysis in E. mixta across the mainland and different islands, 271 male individuals of E. mixta were collected during May and June 2023 using modified traps containing a chemical attractant compound Rigar^®^ Balsam (Laboratorio Rigar S.A., Panamá, Panama). All specimens were described in detail in Vega-Rovira. et al. [9]. Specimens were identified using the taxonomic keys of Roubik and Hanson (2004) [44] and were preserved in 96% ethanol for subsequent processing. Five sites were sampled (Figure 1), Playa Banca (PYB) in the mainland (7°43′33.47″ N, 81°31′45.758″ W) (n = 12), and four islands that varied in area and distance from the mainland: Canales de Afuera (CAN) (7°41′40.995″ N, 81°37′44.651″ W) (n = 26), Coibita (CBI) (7°38′22.062″ N, 81°42′22.057″ W) (n = 76), Coiba (CBA) (7°36′1.89″ N, 81°43′28.76″ W) (n = 78) and Jicarón (JIC) (7°17′22.7040″ N, 081°47′22.4520″ W) (n = 79).
The right forewings were carefully detached, cleaned of excess tissue, air-dried, and mounted flat on microscope slides prior to imaging. The wings preserved on microscope slides are deposited in the G. B. Fairchild Invertebrate Museum, following an alphanumeric registration code that includes locality–individual–wing. Wings were photographed using a stereomicroscope with a calibrated scale. Images were organized into a .tps file using tpsUtil v1.81 [45]. For each wing, 15 landmarks were digitized using the software tpsDig2 v2.31 [45]. The landmarks were placed at the intersections of the wing venation (Figure 2), and their information was extracted by a Procrustes fit, which standardizes the samples, eliminating the effect of size, position and rotation (Rohlf and Slice, 1990) [37].
In order to visualize the wing shape variation, a Principal Component Analysis (PCA) was performed using the Covariance Matrix of Procrustes-aligned coordinates [46]. On the other hand, to reduce the number of shape dimensions and maximize the differentiation between groups (sites), new shape axes were created performing a Canonical Variate Analysis (CVA) [47]. CVA was used as an exploratory tool to maximize between-group shape differences and visualize patterns of population-level structuring, rather than as a predictive classification approach. To statistically evaluate the differences in shape between sites, a permutation test of 10,000 rounds was conducted using the Mahalanobis distances (distances or magnitude of difference between average shapes) extracted from the CVA. Geometric morphometrics analyses were conducted through the software MorphoJ v1.07 [48] and the package “geomorph V. 4.0.10” in R software v 4.04 [49].
Finally, to evaluate whether differences in the shape of the forewings of E. mixta are associated with geographic distances between islands and the mainland, a Mantel test [50] was conducted among sampling points (sites), comparing a matrix of geographic distances with a matrix of Mahalanobis shape distances. The shape distances were extracted from CVA, and the geographical distances between sites were obtained from QGIS software v3.20.2.
3. Results
The Principal Component Analysis (PCA) of the wing shape shows that 33.1% of the variance is explained by the first two components (PC1 = 22.6%; PC2 = 10.5%). The morphospace of these two principal components shows an overlap in the shape of all sites, with a higher variation in the shape along principal component two (Figure 3).
In contrast, the Canonical Variate Analysis shows a subtle differentiation between sites, varying mainly throughout the canonical variate one, and separating the samples into groups composed of Canales, Coibita and Playa Blanca. The two remaining sites (Jicaron and Coiba) individually share a part of their morphospace between them and with the main group (Figure 4).
The superposition of mean wing shapes (Figure 5) reveals subtle but consistent spatial shifts in landmark positions among sites. The most noticeable differences involve the mainland population (Playa Blanca), which shows slight positional displacements relative to insular populations at landmarks associated with vein intersections and inflection points, such as landmark 15. Similarly, landmarks 6 and 1 exhibit small but coherent shifts in position in the Playa Blanca samples compared to the islands. These differences reflect localized changes in wing vein geometry and curvature rather than direct variation in vein length, as geometric morphometric analyses capture relative landmark displacement within the overall wing configuration.
Regarding the shape differences, the permutation test was conducted between the Mahalanobis shape distances, which showed a significant difference in the forewing shape between all sites, except for the wing between individuals of E. mixta from Playa Blanca and Canales de Afuera (Table 1). The highest shape difference was between Jicaron and Canales de Afuera, while the least difference was between Coiba and Jicaron. Finally, the Mantel test shows a positive and significant correlation between the geographical and the Mahalanobis shape distances (r = 0.64; p < 0.05), suggesting that individuals from the most distant sites exhibit higher differentiation in wing shape, while sites closer together have individuals with more similar wing shapes (Figure 6). One pairwise comparison deviates from the overall trend, which is expected given the limited number of sampling sites, and does not affect the significance of the Mantel correlation.
4. Discussion
Beyond their geographic boundaries, islands represent powerful natural laboratories that amplify the effects of spatial structure on evolutionary processes. Islands provide particularly valuable settings for examining evolutionary change because their isolation restricts gene flow among populations and creates opportunities for divergence to accumulate over time [14]. Under such conditions, genetic drift can exert a stronger influence on phenotypic patterns, especially in small or partially isolated populations [51]. These stochastic forces often interact with local ecological pressures, making archipelagos effective arenas for understanding how morphology evolves under limited gene flow and heterogeneous environmental contexts. Geometric morphometrics has proven an effective tool for addressing our central question: whether insular isolation influences wing shape in E mixta. This approach captures small but biologically meaningful variations in shape that traditional linear measurements are less likely to detect [36]. In our dataset, the ability of geometric morphometrics to retain spatial relationships among wing-vein landmarks was essential for uncovering subtle phenotypic differences among populations.
Although the differences observed were subtle, the resulting patterns provide valuable insights into the natural history of E. mixta. In bees, wing venation is a highly conserved trait shaped by aerodynamic demands, developmental constraints, and phylogenetic history, providing mechanical support for flight while allowing limited but informative variation among populations and species (e.g., Cameron [28], Perrard et al. [52]). In Euglossini, despite their strong flight capacity, subtle variation in wing shape and venation has been shown to reflect population-level structuring rather than major functional shifts.
Orchid bees are recognized for their strong flying abilities, wide foraging ranges, and capacity to navigate varied landscapes [28]. These traits often minimize the likelihood of population differentiation in fragmented or isolated systems. Previous studies have shown that high mobility can buffer the effects of isolation by maintaining gene flow across spatially structured environments [19,20]. Under these conditions, morphological divergence among islands would not be expected. Therefore, the consistent shape differences observed here indicate that insular conditions impose ecological or spatial constraints strong enough to generate localized phenotypic changes. This pattern suggests that morphological variation in E. mixta results from an interaction between the regulating effects of dispersal and the diversifying pressures associated with geographic isolation, resource structure, and the environmental characteristics unique to each island.
Patterns of subtle yet statistically robust morphological divergence, such as those observed in E. mixta, have also been reported in other insular insect systems examined using geometric morphometrics. For example, Laojun et al. [6] found that populations of Aedes albopictus and Armigeres subalbatus in the Thai archipelagos showed only partial separation in canonical space, while exhibiting significant Mahalanobis distances between the islands. These findings align with ours, showing that morphological divergence in island insects often results in subtle rather than substantial changes. Similar patterns have been observed in Praocis spinolai (Solier, 1841) from coastal Chile, for which Benítez et al. [27] demonstrated that island populations exhibit subtle but consistent differences in traits related to locomotion and body robustness compared to their mainland counterparts. In the island–mainland comparison of the halictid bee Halictus tripartitus, Ostwald et al. [7] found consistent differentiation in wing venation, despite significant overlap in morphospace. This indicates that population-specific characteristics can emerge even in species with high dispersal potential. Together, these studies support the idea that islands often encourage fine-scale morphological changes in insects. Additionally, geometric morphometrics proves particularly effective for identifying these patterns, even in species with relatively high mobility. There is a strong positive correlation between geographic distance and differences in wing shape, further supporting the idea that spatial isolation influences the phenotypic variation in Euglossa mixta. This pattern aligns with the isolation-by-distance dynamics often observed in insular species, in which even slight reductions in connectivity can lead to noticeable divergence in morphological traits [5].
The differences observed between the most remote island, Jicarón, and the other locations indicate that dispersal between distant islands is either rare or inadequate to fully equalize wing shape. Similar distance-related patterns have been noted in other island-dwelling insects. This pattern aligns with the dynamics of isolation-by-distance that are often observed, as reported by Laojun et al. [6]. They found that the largest Mahalanobis distances, when comparing insular Hymenoptera with mainland populations, occurred between populations separated by wider marine barriers. Similar patterns have also been observed in studies of non-native species, where gradual divergence can occur even in the presence of ongoing gene flow among localities. For example, spatial differentiation has been detected in Trichocera maculipennis Meigen, 1830 across Antarctic stations [23], with greater wing shape divergence observed at the geographically most isolated locality, separated by Fildes Bay and the Collins Glacier.
Although geographic isolation is a primary factor in shaping wing shape in E. mixta, environmental variation between the islands could also contribute to the observed phenotypic patterns. Differences in vegetation structure, availability of floral resources, microclimatic conditions, and wind exposure can all influence flight behavior and aerodynamic demands, which could generate specific functional demands at the island level on wing morphology [53,54]. Similar environmentally driven variations have been documented in other insects, where local habitat conditions affect wing venation and flight dynamics [55,56]. In this context, the subtle but consistent differences observed among islands may indicate an interaction between spatial isolation and the performance demands imposed by the environment. Future studies that incorporate habitat descriptors or directly measure flight behavior would help clarify the extent to which local ecological conditions contribute to the morphological differentiation seen in E. mixta. Finally, our results show that E. mixta exhibits subtle but consistent morphological structuring across the Coiba archipelago, revealing how insular environments can shape phenotypic variation even in species with high dispersal capacity. Although the detected differences in wing shape are smaller, the agreement across multivariate analyses, the significant Mahalanobis distances, and the positive association between geographic distance and shape variation indicate that these patterns reflect more than sampling noise. Instead, they point to a landscape of partial connectivity in which both spatial isolation and local environmental conditions contribute to fine-scale morphological differentiation. While our data do not allow us to determine whether these shifts are genetically based or environmentally induced, the patterns are consistent with early-stage divergence observed in other island insects. In Hymenoptera, males are haploid, which may allow genetic differences among populations to be more directly expressed at the phenotypic level [57,58]. However, because our analyses rely exclusively on wing shape data, we cannot disentangle the relative contributions of genetic differentiation and phenotypic plasticity. Future work integrating genomic data, environmental variables and functional measures of flight performance will be essential for assessing the relative contributions of drift, selection and plasticity. Nonetheless, this study demonstrates that geometric morphometrics is a sensitive and informative tool for detecting subtle phenotypic divergence in E. mixta, providing a foundation for understanding how insular systems influence the evolutionary trajectories of highly mobile pollinators.
5. Conclusions
This study shows that the wing shape of Euglossa mixta remains consistent mainly across the Coiba archipelago and the mainland. However, subtle morphological variations have emerged due to insular isolation. These patterns suggest that even highly mobile pollinators can experience spatial constraints that allow for fine-scale differences in their traits. Instead of showing either substantial divergence or complete uniformity, the morphology of E. mixta represents a balance between its ability to disperse and the geographic layout of the island systems. This underscores how insular environments can influence pollinators’ functional traits, highlighting the interplay between mobility, environmental conditions, and spatial structures in shaping phenotypic variation. In the future, integrating genomic, ecological, and functional data will be critical for uncovering the mechanisms behind these patterns and evaluating their significance in the context of pollinator ecology and island biogeography.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kingsolver J.G. Huey R.B. Introduction: The evolution of morphology, performance, and fitness Integr. Comp. Biol.20034336136610.1093/icb/43.3.36121680444 · doi ↗ · pubmed ↗
- 2Miner B.G. Sultan S.E. Morgan S.G. Padilla D.K. Relyea R.A. Ecological consequences of phenotypic plasticity Trends Ecol. Evol.20052068569210.1016/j.tree.2005.08.00216701458 · doi ↗ · pubmed ↗
- 3Losos J.B. Ricklefs R.E. Adaptation and diversification on islands Nature 200945783083610.1038/nature 0789319212401 · doi ↗ · pubmed ↗
- 4Thompson J.N. Rapid evolution as an ecological process Trends Ecol. Evol.19981332933210.1016/S 0169-5347(98)01378-021238328 · doi ↗ · pubmed ↗
- 5Emerson B.C. Evolution on oceanic islands: Molecular phylogenetic approaches to understanding pattern and process Mol. Ecol.20021195196610.1046/j.1365-294X.2002.01507.x 12030975 · doi ↗ · pubmed ↗
- 6Laojun S. Sontigun N. Chaiphongpachara T. Influence of insular conditions on wing phenotypic variation in two dominant mosquito vectors, Aedes albopictus and Armigeres subalbatus (Diptera: Culicidae), in the border archipelagos of Thailand Med. Vet. Entomol.20243834936010.1111/mve.1272238641881 · doi ↗ · pubmed ↗
- 7Ostwald M.M. Thrift C.N. Seltmann K.C. Phenotypic divergence in an island bee population: Applying geometric morphometrics to discriminate population-level variation in wing venation Ecol. Evol.202313 e 1008510.1002/ece 3.1008537181201 PMC 10172614 · doi ↗ · pubmed ↗
- 8Whittaker R.J. Fernández-Palacios J.M. Matthews T.J. Borregaard M.K. Triantis K.A. Island biogeography: Taking the long view of nature’s laboratories Science 2017357 eaam 832610.1126/science.aam 832628860356 · doi ↗ · pubmed ↗