The Influence of Acute Cold Stress on Intestinal Health of the Juvenile Chinese Soft-Shelled Turtle (Pelodiscus sinensis)
Xiaona Ma, Qing Shi, Zhen Dong, Chen Chen, Junxian Zhu, Xiaoli Liu, Xiaoyou Hong, Chengqing Wei, Xinping Zhu, Weijia Song, Wei Li, Liqin Ji

TL;DR
This study explores how sudden cold stress affects the intestines of Chinese soft-shelled turtles, revealing changes in gut health and molecular responses.
Contribution
The study provides a comprehensive analysis of physiological, microbial, and molecular responses to acute cold stress in Chinese soft-shelled turtles.
Findings
Acute cold stress at 7°C caused severe intestinal damage, including inflammation and tissue necrosis.
Cold stress altered gut microbiota, with Helicobacter and Citrobacter dominating at 7°C.
Transcriptomic and metabolomic analyses identified key pathways like Toll-like receptor signaling and linoleic acid metabolism affected by cold stress.
Abstract
As a poikilothermic species, the Chinese soft-shelled turtle is highly sensitive to changes in environmental temperature. Sudden drops in temperature often lead to mass mortality, causing substantial economic losses to the aquaculture industry. This study aims to uncover the mechanisms of damage induced by low-temperature stress in the Chinese soft-shelled turtle. Through integrated analysis of intestinal microbiota, molecular expression, metabolite profiles, and related signaling pathways, we further elucidate the physiological and molecular responses of this species to cold stress, providing a scientific basis for understanding its low-temperature adaptability. Sharp declines in temperature pose a significant risk for mass mortality events in the Chinese soft-shelled turtle (Pelodiscus sinensis). To assess the effects of acute cold stress on intestinal health, turtles were exposed to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
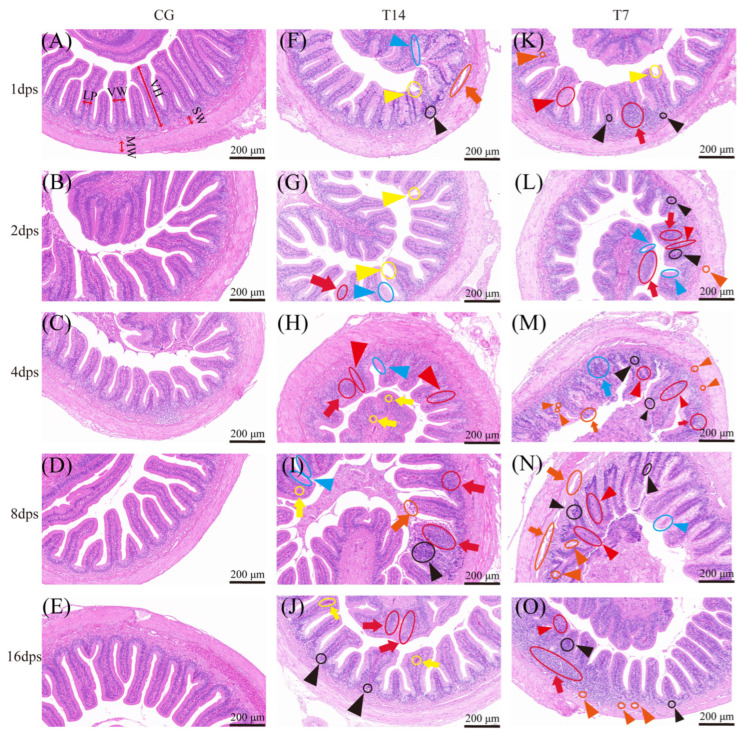 Figure 1
Figure 1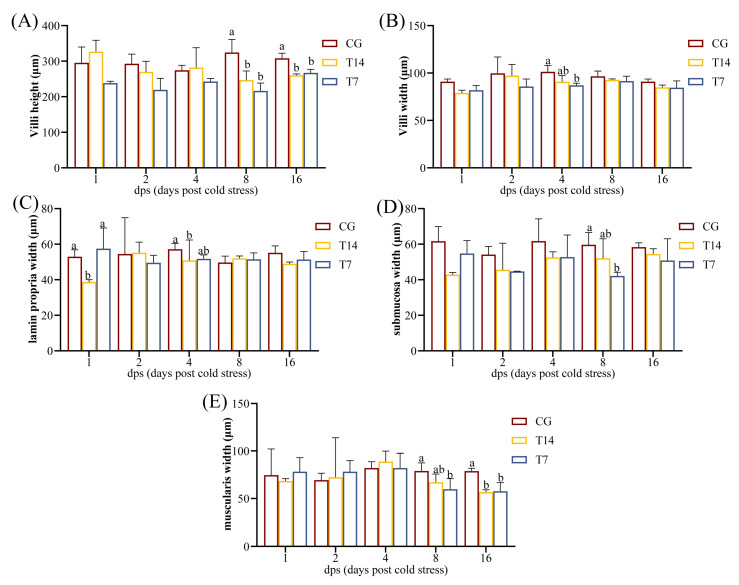 Figure 2
Figure 2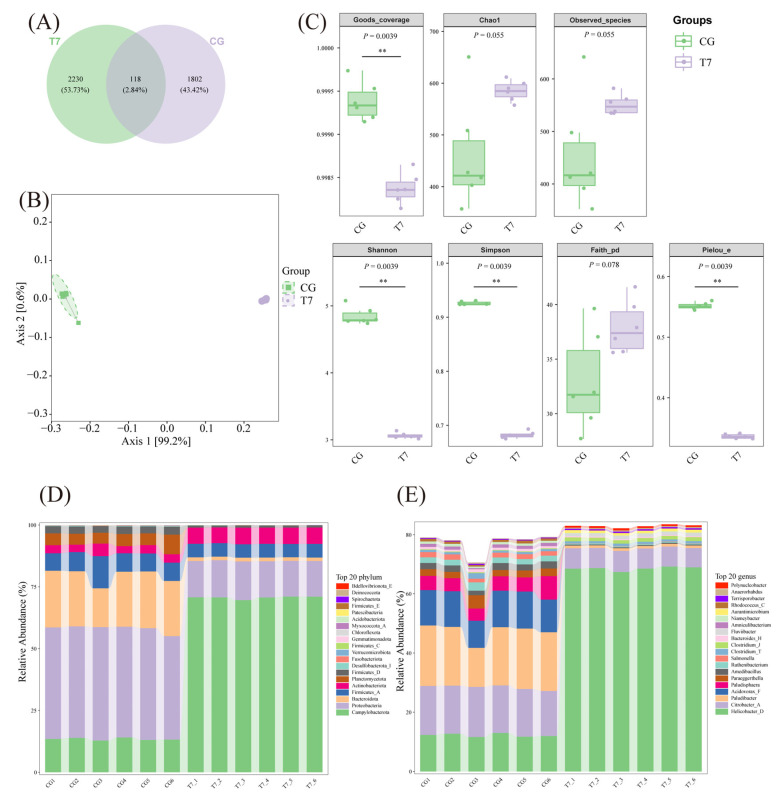 Figure 3
Figure 3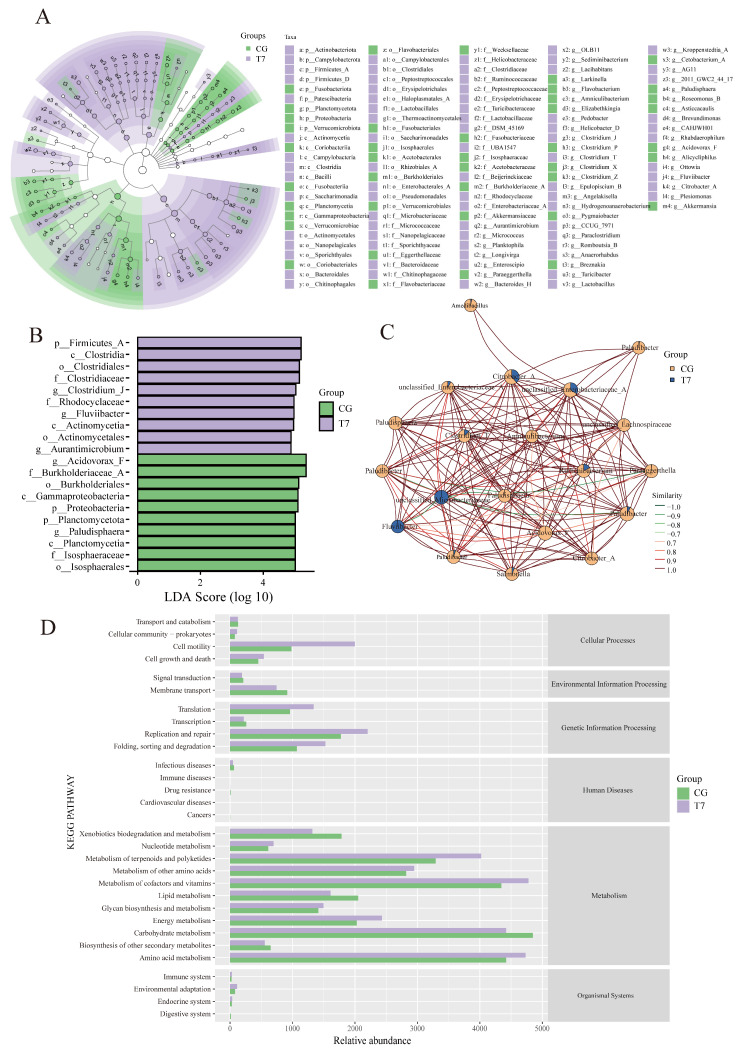 Figure 4
Figure 4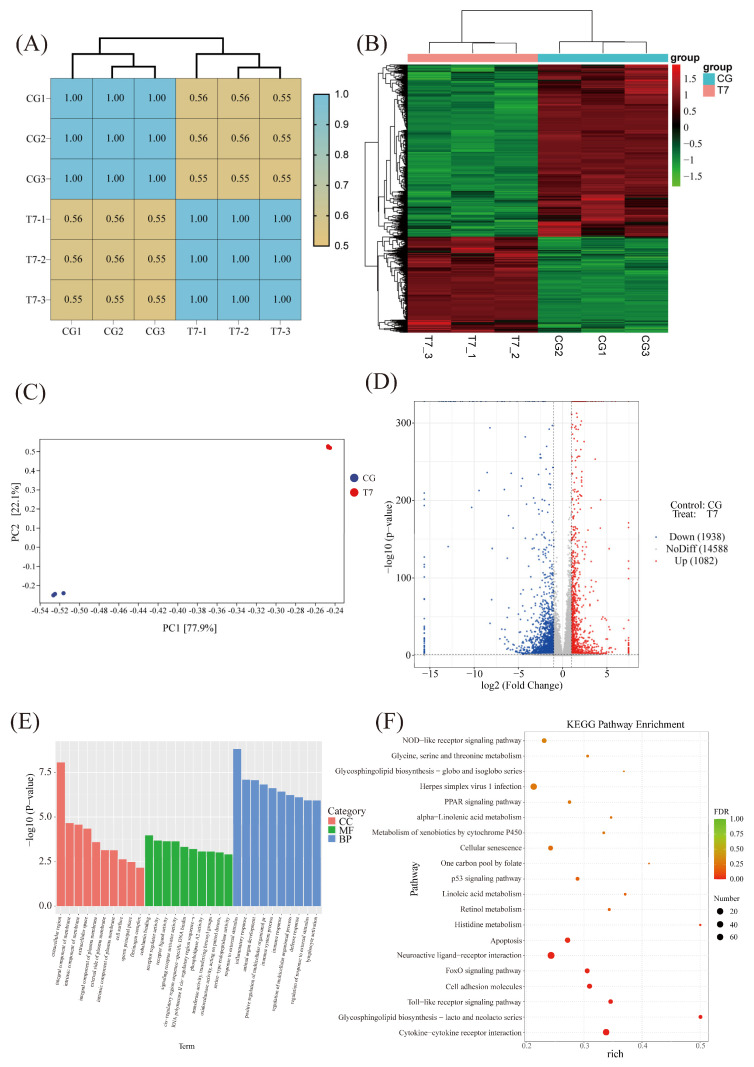 Figure 5
Figure 5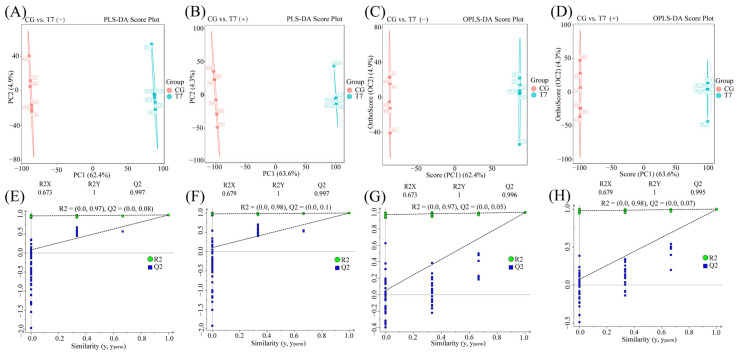 Figure 6
Figure 6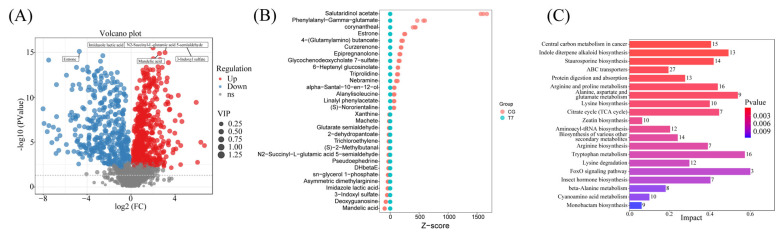 Figure 7
Figure 7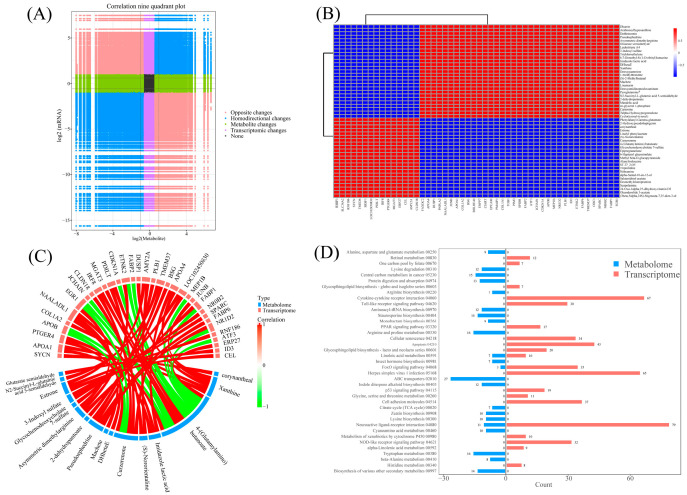 Figure 8
Figure 8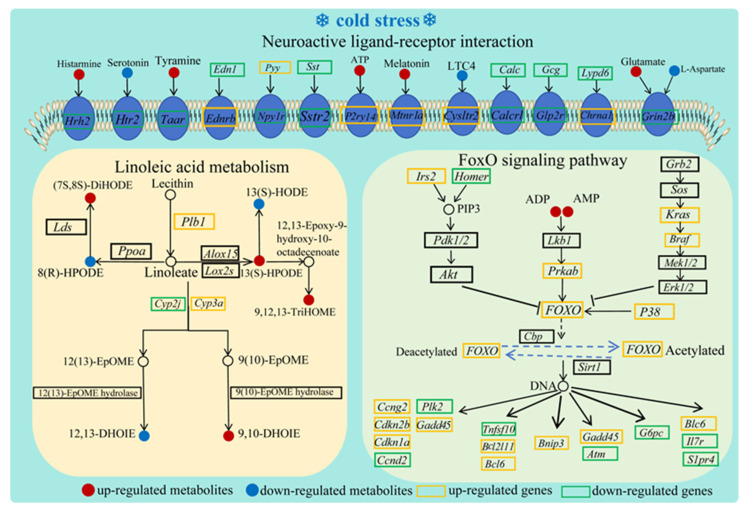 Figure 9
Figure 9- —National Natural Science Foundation of China
- —Science and Technology Program of Guangzhou
- —Central Public-interest Scientific Institution Basal Research Fund
- —Key Laboratory of Environment Controlled Aquaculture (Dalian Ocean University) Ministry of Education
- —Key Laboratory of Tropical and Subtropical Fishery Resources Application and Cultivation, Ministry of Agriculture and Rural Affairs
- —Selection Breeding of the high-quality fast-growing Chinese Soft-Shelled Turtle and Studies on Efficient and Healthy Farming Technology
- —the Key Laboratory of Marine Environment Exploration Technology and Application of the Ministry of Natural Resources
- —the China─ASEAN Maritime Cooperation Fund
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTurtle Biology and Conservation · Physiological and biochemical adaptations · Aquaculture disease management and microbiota
1. Introduction
The Chinese soft-shelled turtle (Pelodiscus sinensis), as a member of the class Reptilia, order Testudines, and family Trionychidae, is a traditional and economically important aquaculture species in Southeast Asia. It is popular in Chinese markets for its high protein content and nutritional value, particularly its richness in bioactive peptides [1]. By 2023, national production of the Chinese soft-shelled turtle had reached nearly 0.5 million tons [2]. As an ectotherm with an optimal temperature range of 25–32 °C, this species is highly susceptible to water temperature fluctuations, which can cause mass mortality [3]. However, the adverse effects of acute cold stress on the Chinese soft-shelled turtle remain less comprehensively studied in comparison with other aquatic species.
Given the increasing frequency of extreme low-temperature events occurring from climate change and seasonal variation [4], the impact of cold stress on aquatic animals has been widely studied, especially in fishes [5]. A drastic reduction in temperature can profoundly affect physiological processes, metabolic traits, and immune response [6]. For instance, cold stress can alter biochemical parameters, cause DNA damage, and interfere with immune processes in the orange spotted grouper [7]. Research on Takifugu fasciatus has revealed that cold exposure leads to hepatic structural damage, fibrosis, oxidative stress, and lipid peroxidation [8]. Further research on hybrid sturgeon has found that acute cold stress suppresses the immune response by regulating apoptosis and the Toll-like receptor signaling pathway [9]. In contrast, the influence of low temperature on the Chinese soft-shelled turtle remains less well understood [10]. Therefore, investigating the adverse effects of cold stress is crucial for developing effective strategies to mitigate cold-stress-induced damage or death.
Seasonal cooling is a slow and gradual process, providing aquatic animals sufficient time to mobilize physiological compensation mechanisms—including the regulation of metabolic pathways and gene expression—for successful cold adaptation [5]. Conversely, acute cold stress occurs when water temperatures decline within hours, eliciting intensive physiological responses and exerting oxidative stress [9]. Although seasonal temperature variation can influence aquatic health, it typically enhances long-term survival through physiological acclimation. However, acute cold stress commonly surpasses an organism’s capacity to maintain homeostasis, leading to behavioral imbalances, incapacitating shock, and high mortality rates [11].
The intestine is a vital organ responsible for nutrient absorption and immune response [12]. Its physiological functions are highly sensitive to environmental stimuli [13], particularly temperature variations [14]. Studies on largemouth bass have found that an acute decrease in temperature alters gut morphology, characterized by increased villus length and reduced goblet cell numbers [15]. Temperature fluctuation leads to intestinal dysbiosis, subsequently leading to metabolic disorders and impaired immune function [16,17]. Similarly, research on tsinling lenok trout shows that heat stress results in gut microbial disturbance and metabolic dysregulation [14].
Our previous work revealed that acute cold stress triggers energy metabolic disorders, tissue damage, and apoptosis in the liver of the Chinese soft-shelled turtle [18]. The liver as the metabolic center and the intestine as the primary immune and barrier organ are functionally linked via the gut–liver axis, which facilitates bidirectional functional communication. To understand the organism’s systemic response to acute cold stress, the present study was thus conducted to evaluate the effects on the intestine, aiming to elucidate the distinct and coordinated physiological responses in the two organs. Our findings will provide novel insights into the intestinal adaptive mechanisms against acute cold stress, while offering potentially practical strategies for maintaining intestinal homeostasis in the Chinese soft-shelled turtle under acute cold stress.
2. Materials and Methods
2.1. Animal Ethics
The animal trials in this research complied with Chinese guidelines for laboratory animal care and use. The animal trial procedures were approved by the Ethics Committee of the Pearl River Fisheries Research Institute, Chinese Academy of Fishery Sciences (Approval No. LAEC-PRFRI-2023-10-15).
2.2. Experimental Design
The Chinese soft-shelled turtles were kindly gifted by the Huizhou Wealth Xing Industrial Co., Ltd. (Huizhou, China). A total of 180 juvenile turtles, aged 3 months with an average weight of 12 ± 3 g, were randomly selected for this research. Sex could not be determined visually at this juvenile stage and was therefore not considered. All of the experimental turtles were in good health, displaying no signs of disease or abnormal behavior before the experiment. The trial was conducted in the Pearl River Fisheries Research Institute (Guangzhou, China). Before initiating the cold stress challenge, the turtles were acclimated in polyethylene boxes (1 m × 1 m × 0.25 m) for 14 days. During acclimation, they were fed commercial pellets gifted by Guangdong Nutriera Group Co., Ltd. (Guangzhou, China) twice daily at 9:00 and 17:00 until visibly satiated. The water quality parameters were maintained as follows: temperature 28 ± 1 °C, pH 8.2 ± 0.4, NO_2_^−^ 1.0 ± 0.3 mg/L, NH_4_^+^-N 1.1 ± 0.5 mg/L, and dissolved oxygen 5.8 ± 1.5 mg/L. One-third of the farming water was changed once a week to maintain stable water conditions.
After acclimation, the 180 Chinese soft-shelled turtles were randomly and evenly distributed into 18 polyethylene boxes (48 cm × 35 cm × 17 cm, 10 turtles per box). These boxes were randomly assigned to three groups with three replicates: a 28 °C control group (CG) and 14 °C and 7 °C acute cold stress groups (T14 and T7). After a 24 h starvation period, the turtles were transferred to RXZ-436 cooling incubators (Ningbo Jiangnan Instrument Factory, Ningbo, China) for the acute cold stress challenge. The water temperature in the incubators was gradually lowered from 28 °C to the target temperatures at a rate of 1 °C/h and then kept at 28 °C, 14 °C, and 7 °C for 1, 2, 4, 8, and 16 days.
2.3. Sample Collection
The beginning of the acute cold stress experiment was defined as the time point when the water reached the target temperature. Sampling was conducted at 1, 2, 4, 8, and 16 days post cold stress (dps) with 12 replicates at each time point for each group. Before sampling, the turtles were anesthetized by immersion in a 1 g/L solution of tricaine methanesulfonate (MS-222). Afterwards, the turtles were quickly dissected on ice to collect the intestine. The small intestine, defined as the proximal 65% of the total intestinal length (regions S1–S5, Supplementary Figure S1) [19], was chosen for its primary roles in nutrient absorption and immune function. After fixation in Bouin’s solution overnight, the intestinal tracts were washed three times (10 min each) with 70% ethanol to remove residual fixative. The remaining small intestinal tracts from 12 individuals per group were pooled into 6 tubes (intestines from two turtles within the same tank were pooled into one sample), snap-frozen in liquid nitrogen, and stored at −80 °C for subsequent analysis of the microbiome, transcriptome, and metabolome.
2.4. Intestinal Histological Analysis
After overnight fixation in Bouin’s solution, the intestinal tracts were washed with 70% ethanol three times (10 min each) to remove residual Bouin’s solution. The intestinal tracts were then dehydrated through a graded ethanol series (80%, 90%, 95%, and 100%), equilibrated with xylene, and embedded in paraffin. Tissue sections (4 μm) were prepared using a Leica RM 2016 microtome (Leica Biosystems Co., Ltd., Wetzlar, Germany), mounted on slides, dewaxed in xylene, and stained with hematoxylin and eosin (H&E). After staining, sections were sealed with resin, and intestinal morphology was observed and captured using a NIKON ECLIPSE 100 upright optical microscope (Nikon Corporation, Tokyo, Japan). The morphological parameters, including villus height, villus width, lamina propria width, submucosal width, and muscularis layer width, were measured using ImageJ 1.44 software (National Institute of Health, Bethesda, MD, USA). For histological analysis, three individuals per group (n = 3) were assessed at each time point. The value for each individual was derived from the mean of five measurements per parameter. Consequently, statistical analysis was performed using three mean values as independent data points (n = 3) for each group at each time point.
2.5. Intestinal Microbiota Analysis
Six mixed intestinal samples (n = 6) per group were analyzed for microbiota analysis, with each sample representing a blend of intestines from two turtles within the same tank. The Fast DNA SPIN kit (MP Biomedicals, Irvine, CA, USA) was utilized for microbial genomic DNA extraction. After assessing the DNA quality and concentration, the V3-V4 hypervariable region of the bacterial 16S rDNA gene was amplified using primers 338F: 5′-ACTCCTACGGGAGGCAGCA-3′ and 806R: 5′-GGACTACHVGGGTWTCTAAT-3′ with the diluted DNA as a template. PCR amplification was conducted in a final volume of 25 µL. The reaction mixture comprised 5 µL of 5 × Q5 Reaction Buffer, 5 µL of 5× Q5 High GC Buffer, 0.25 µL of Q5 High-Fidelity DNA Polymerase, 2 µL of dNTPs, 1 µL each of forward and reverse primers, 2 µL of template DNA, and nuclease-free water to volume. The PCR program was set as follows: an initial denaturation at 98 °C for 5 min, followed by 25 cycles of 98 °C for 30 s, 52 °C for 30 s, and 72 °C for 45 s, with a final extension at 72 °C for 5 min. All samples were run in triplicate, and a no-template control (NTC) was included for quality assurance. The PCR product was purified using the Omega DNA purification kit (Omega Inc., Norcross, GA, USA) and sequenced on the Illumina NovaSeq 6000 platform by Suzhou PANOMIX Biomedical Tech Co., Ltd., China. The final clean reads were obtained by removing chimeric sequences from raw data.
Sequences were assigned to the same operational taxonomic unit (OTU) when their similarity exceeded 97%, as implemented in the Uparse software (version 7.0.1001) [20]. Alpha diversity indices (Good’s coverage, Chao1, Observed species, Shannon, Simpson, Faith’s PD, and Pielou’s evenness), β diversity (principal coordinate analysis), and bacterial abundance at the phylum and genus levels were analyzed with QIIME2 software. The linear discriminant analysis (LDA) effect size (LEfSe) was employed to identify differentially abundant taxa between CG and T7. The potential function of the microbial communities was predicted from the 16S rDNA data using PICRUSt2. The resulting gene families were then annotated against the KEGG database (http://www.genome.jp/kegg/pathway.html, (accessed on 24 November 2024)). The abundances of KEGG pathways were compared between the CG and T7 groups to assess differences in metabolic functions.
2.6. Intestinal Transcriptomic Analysis
Small intestines from the CG and T7 groups at 8 days post stress (dps) were collected and subjected to transcriptomic analysis. Three pooled samples (n = 3), each representing a mix from four individuals in the same box, were analyzed. Total RNA was isolated from the intestine using TRIzol reagent (Invitrogen Life Technologies, Waltham, MA, USA). RNA integrity was initially assessed via electrophoresis on 1% agarose gels. RNA concentration and purity were determined using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Furthermore, the RNA Integrity Number (RIN) was precisely quantified with an Agilent 2100 Bioanalyzer (Agilent Technologies, Waltham, MA, USA). Only RNA samples with an RIN greater than 7 were used to construct the cDNA library.
Raw data were processed to remove adapters and low-quality reads. The clean data were subsequently mapped to the Chinese soft-shelled turtle genome (https://www.ncbi.nlm.nih.gov/datasets/genome/GCF_000230535.1/ (accessed on 24 July 2012)). Differentially expressed genes (DEGs) between CG and T7 were identified using the DESeq2 R package (v1.20.2), with screening thresholds set at |log_2_(fold change)| ≥ 1.0 and adjusted p < 0.05. Subsequently, these DEGs were functionally annotated using the Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) databases.
2.7. Intestinal Metabolomic Analysis
Six small intestines per group from CG and T7 at 8 dps were used for metabolomic analysis (n = 6). Metabolites were extracted from 100 mg of intestine with a 1 mL methanol/chloroform/water mixture (27:3:1, v/v/v), which was then homogenized, ultrasonicated, and centrifuged at 12,000 rpm for 10 min. After vacuum drying, the supernatant was redissolved in 400 µL of methanol-water (1:1) containing 4 ppm 2-chloro-L-phenylalanine as the LC-MS sample. Chromatographic separation was achieved on a Thermo Vanquish ultra-high-performance liquid chromatography system equipped with an ACQUITY UPLC^®^ HSS T3 column (Waters, Milford, MA, USA). System stability was monitored by analyzing a quality control (QC) sample, which was a pool of equal aliquots from all experimental samples, alongside the study samples using a Thermo Q Exactive HF-X mass spectrometer (Thermo Fisher Scientific, Waltham, MA, USA) in both positive and negative ionization modes. Raw data were converted to mzXML format using ProteoWizard (ver. 3.0), and the XCMS package (ver. 3.1) was used for peak identification, filtering, and alignment [21]. Metabolites were annotated by mapping with public databases, including the standard BioNovoGene database (http://www.bionovogene.com), Metlin (http://metlin.scripps.edu), Mzclound (https://www.mzcloud.org), LipidMaps (http://www.lipidmaps.org), Massbank (http://www.massbank.jp), and the Human Metabolome Database (http://www.hmdb.ca). Multivariate statistical analysis, including partial least squares discriminant analysis (PLS-DA) and orthogonal partial least squares discriminant analysis (OPLS-DA), was performed. Differentially expressed metabolites (DEMs) between groups were screened by the criteria of VIPs > 1 and p < 0.05. The DEMs were functionally annotated with the KEGG database.
2.8. Integrated Transcriptomic and Metabolomic Analysis
An integrative analysis of the DEGs and DEMs was conducted to identify significantly changed biological pathways. Pearson’s correlation analysis was performed to evaluate the relationships between the DEGs and DEMs. Pairs with a Pearson’s correlation coefficient (|PCC|) > 0.80 and p < 0.05 were considered significantly correlated. Furthermore, DEGs and DEMs were matched with the KEGG database to reveal their common pathways.
2.9. Validation of Transcriptomic Data
To validate the RNA-seq findings, quantitative real-time PCR (qRT-PCR) was conducted using the same RNA samples prepared for sequencing. Nine DEGs were selected for verification. Each sample was run with three technical replicates. The primers used in qRT-PCR are listed in Table S1. β-actin was selected as the reference gene for normalization. The 2^−△△CT^ method was applied to calculate the fold changes in the mRNA levels in T7 relative to CG [22].
2.10. Statistical Analysis
The experimental unit used in this research was the tank. Histological parameters (n = 3) were analyzed with the Kruskal–Wallis H test, followed by Dunn’s post hoc test for multiple comparisons. The qRT-PCR validation results and α diversity indices of the intestinal microbiota, including Chao1, Shannon, Good’s coverage, Observed species, Simpson, Faith’s PD, and Pielou’s evenness, were evaluated using the Mann–Whitney U test. All statistical analyses were conducted with GraphPad Prism 9 software (GraphPad Software Inc., La Jolla, CA, USA). Data were expressed as mean ± standard error of the mean (SEM). Statistical significance was defined as p < 0.05.
3. Results
3.1. Histopathological Injury and Intestinal Morphology
The histopathological analysis revealed cold-stress-induced microstructural damage in the intestine (Figure 1). The small intestine in the CG maintained normal structure throughout the experiment, with intact, evenly arranged villi and mucosal epithelium. Mild injuries appeared in both T14 and T7 from 1 to 2 dps (Figure 1F,G,K,L), including inflammatory cell infiltration, lymphoid hyperplasia, mucosal epithelial degeneration, and villus defects such as villus splitting and fusion. Lesions worsened from 4 to 8 dps (Figure 1H,I,M,N), particularly in T7, where severe villus fusion, erosion, extensive epithelial necrosis and desquamation, and obvious fissures in the muscle layer and submucosa were observed.
By 16 dps (Figure 1J,O), histological damage was remarkably ameliorated in both T14 and T7. The villi exhibited well-defined borders, largely intact architecture, and uniform distribution, although mild inflammatory cell infiltration and lymphoid hyperplasia persisted.
To evaluate the effect of acute cold stress on intestinal morphology, villus height, villus width, lamina propria width, submucosal width, and muscularis layer width were measured (Figure 2). Compared with the CG, the villus height (Figure 2A) in the T14 and T7 groups was significantly reduced at 8 and 16 dps (p < 0.05). T7 exhibited significantly narrower villus width (Figure 2B) than the CG at 4 dps (p < 0.05). The lamina propria width (Figure 2C) was also reduced in the T14 group at 1 and 4 dps (p < 0.05). The submucosal width (Figure 2D) in T7 was smaller than in CG at 8 dps (p < 0.05). Additionally, the muscularis layer width (Figure 2E) in T14 showed significant shortening at 16 dps (p < 0.05), while that of T7 showed significant reductions after both 8 and 16 dps (p < 0.05).
Histopathological injuries and morphological changes in the intestine were observed in a time-dependent manner under both 14 °C and 7 °C stress conditions. Notably, the intestinal damage caused by 7 °C low-temperature stress was more severe than that in the 14 °C group. Given the substantial damage caused by 7 °C exposure at 8 dps, small intestinal samples from T7 and CG at 8 dps were selected for comprehensive multi-omics analysis.
3.2. Intestinal Microbial Composition
A total of 1802 and 2230 OTUs were sequenced in CG and T7, respectively, with 118 OTUs shared between the two groups (Figure 3A). The alpha diversity analysis (Figure 3B) consisted of multiple indices: Good’s coverage (representing sequencing depth), Observed species and Chao1 (indicating species richness), Simpson and Shannon (reflecting species diversity), Faith’s PD (indicating phylogenetic diversity), and Pielou’s evenness. The Good’s coverage values exceeded 99.50% in both groups, demonstrating comprehensive sample coverage. Compared with CG, the Simpson, Shannon, and Pielou’s evenness in T7 showed lower values (p < 0.05). The p values of Chao1 index (p = 0.055), observed species index (p = 0.055), and Faith pd index (p = 0.078) were close to 0.05. Given the small sample size and the limited statistical power of non-parametric tests employed, these results suggest that these parameters might reach statistical significance in a larger-scale trial. PCoA based on OUT levels revealed clear separation between the two groups, with axes 1 and 2 accounting for 99.2% and 0.6% of the variations, respectively (Figure 3C). The assessment of the microbial community structure found distinct composition at both the phylum (Figure 3D) and genus levels (Figure 3E). The T7 group was dominated by Campylobacterota, Proteobacteria, and Actinobacteriota at the phylum level, and by Helicobacter (68.56%) and Citrobacter (6.83%) at the genus level. In contrast, the CG was mainly composed of Proteobacteria, Bacteroidota, and Campylobacterota, with Paludibacter (18.87%) and Citrobacter (16.17%) as the major genera. These results indicate that acute cold stress significantly altered the structure and diversity of the intestinal microbial community.
3.3. Alterations in Phenotypic Characteristics of Intestinal Bacteria
To better characterize the microbial composition variations under acute cold stress, cladogram and LEfSe analyses were performed (Figure 4A). T7 showed enrichment of five phyla, five classes, 16 orders, 18 families, and 32 genera, while the CG exhibited the dominance of four phyla, five classes, seven orders, eight families, and 17 genera. The LDA effect size distribution (Figure 4B) further identified the top 10 dominant taxa (LDA score > 4). In T7, the predominant taxa were the genera Clostridium J, Fluviibacter, and Aurantimicrobium, the families Clostridiaceae and Rhodocyclaceae, the orders Clostridiales and Actinomycetales, the classes Clostridia and Actinomycetia, and the phylum Firmicutes A.
The bacterial correlation networks (Figure 4C) revealed distinct network properties between groups: the T7 exhibited fewer nodes, lower density, and reduced clustering coefficients compared to CG, indicating disrupted bacterial connectivity and weakened community stability. Functional prediction showed that acute cold stress significantly altered metabolic pathways (Figure 4D). The pathways “cell motility”, “metabolism of terpenoids and polyketides”, and “folding, sorting and degradation” were enhanced, whereas the “Xenobiotic biodegradation and metabolism”, “lipid metabolism”, and “carbohydrate metabolism” pathways were suppressed. These findings demonstrated that acute cold stress restructured microbial community architecture and reprogrammed metabolic capacity, potentially affecting host energy homeostasis and stress adaptation.
3.4. Transcriptomic Profiling of Intestine Under Acute Cold Stress
3.4.1. Sequencing and Functional Enrichment of DEGs
Transcriptomic sequencing revealed approximately 52 million raw reads per group (Table S2). After quality control, over 50 million clean reads were retained in each group, with more than 83% successfully mapped to the reference genome. All samples exhibited high sequencing quality, with Q20 and Q30 values over 96% and GC contents averaging approximately 48%. These data confirmed the RNA-seq data were reliable for subsequent analysis.
Three biological repeats with PCCs over 0.95 in all groups showed high reproducibility (Figure 5A). Both clustering (Figure 5B) and principal component analysis (PCA) (Figure 5C) revealed obvious distinctions between CG and T7. The volcano plot exhibited 3020 DEGs, including 1082 upregulated and 1938 downregulated genes (Figure 5D). GO analysis uncovered significantly enriched terms in biological process (BP), molecular function (MF), and cellular component (CC) (Figure 5E). The key DEGs in BP included “response to external stimulus” (Go:0009605) and “inflammatory response” (Go:0006954). Enriched terms in CC were involved in “extracellular region” (Go:0005576) and “integral component of membrane” (Go:0016021), while MF was dominated by “cobalamin binding” (Go:0031419) and “receptor regulator activity” (Go:0030545).
The KEGG enrichment analysis revealed 20 significantly enriched pathways (Figure 5F), 10 of which were involved in immune function (Table S3), such as “cytokine–cytokine receptor interaction” (including Edar, Cntf, Il17b, Epor, Cxcr5, Ccr6, Bmpr1b, and Ccr7), “FoxO signaling pathway” (including Cdkn1a, Igf1, Bcl6, Bnip3, Tnfsf10, S1pr4, Egf, and Plk2), and “apoptosis” (including Tuba1c, Ctsz, Ctsb, Kras, Hras, Pik3ca, Bcl2l11, Gadd45a, Pmaip1, Ddit3, and Fos).
3.4.2. qRT-PCR Verification
The reliability of the transcriptomic data was validated by quantifying the mRNA expression levels of nine randomly selected genes using qRT-PCR. (Figure S2). The qRT-PCR results showed consistent expression patterns with transcriptomic data, confirming the accuracy and reproducibility of RNA-seq analysis.
3.5. Metabolomic Profiling of Intestine
3.5.1. Multivariate Statistical Analysis
Multivariate statistical models effectively distinguished the metabolic profiles between CG and T7. Both PLS-DA (Figure 6A,B,E,F) and OPLS-DA models (Figure 6C,D,G,H) demonstrated clear separation in POS and NEG. The contribution rate and predictability of two models respectively evaluated by R2Y and Q2 parameters indicated their reliability and separating capacity for group discrimination, with R2Y = 1 cum and Q^2^ > 0.995 cum.
3.5.2. Screening for Differential Metabolites
A comparative metabolomic analysis identified 886 DEMs between CG and T7, with 495 upregulated and 391 downregulated (Figure 7A). Z-score plots (Figure 7B) highlighted the most significantly decreased metabolites of salutaridinol acetate, phenylalanyl-γ-glutamate, corynantheal, estrone, and 4-(glutamylamino) butanoate, as well as elevated mandelic acid, deoxyguanosine, 3-indoxyl sulfate, imidazole lactic acid, and asymmetric dimethylarginine in T7. The KEGG analysis revealed predominantly enriched pathways (Figure 7C), including “indole diterpene alkaloid biosynthesis”, “ABC transporters”, “arginine and proline metabolism”, “lysine biosynthesis”, and “citrate cycle”.
3.6. Integrated Analysis of Transcriptome and Metabolome
Pearson correlation analysis identified significant relationships between DEGs and DEMs (Figure 8A,B), with 50 gene–metabolite pairs showing strong correlations (|PCC| > 0.80 and p < 0.05).
Genes such as faxdc2, apoa4, dusp1, pnpla2, and naaladl1 exhibited positive correlations with metabolites, such as dhurrin, arabinosylhypoxanthine, dothistromin, pseudoephedrine, and asymmetric dimethylarginine, but negative correlations with phenylalanyl-gamma-glutamate, 2-hydroxypseudobaptigenin, corynantheal, estrone, and linalyl phenylacetate. Conversely, reciprocal patterns were found for rspr1, slc34a2, rnf186, sycn, and tmed6 expression. A chord diagram (Figure 8C) further delineated specific correlations between top-ranked DEGs and DEMs. For instance, 4-(Glutamylamino)-butanoate was positively associated with Pdilt and Mgat3 but negatively related to Apob and Egr1. Three critical pathways—“linoleic acid metabolism”, “FoxO signaling pathway”, and “neuroactive ligand–receptor interaction”—were enriched in both DEGs and DEMs, suggesting their pivotal role in acute cold stress response (Figure 8D and Figure 9).
4. Discussion
Exposure to low temperature triggers significant physiological, immunological, and metabolic dysregulation in aquatic species, often leading to observable morphological alterations and tissue damage of the intestine [23]. While the detrimental effects of extreme temperatures on intestinal health are known in several aquatic species [15,24,25], the intestinal response of Chinese soft-shelled turtles to acute cold stress remains poorly characterized. Consequently, the histology, microbiome, transcriptome, and metabolome analyses were integrated to elucidate the adaptive mechanisms of the intestine under acute cold exposure.
4.1. Effects of Low Temperature on Intestinal Structure
Histopathological injury and intestinal morphology are key indicators for evaluating intestinal health under environmental stress [26,27]. Under normal conditions, the intestinal epithelium of Pelodiscus sinensis is characterized by a pseudostratified columnar structure with tightly connected enterocytes and abundant mucous cells [28]. In this study, exposure to 14 °C and 7 °C induced progressive intestinal damage in a temperature- and time-dependent manner. Early stages (from 1 to 2 dps) featured inflammatory cell infiltration and lymphoid hyperplasia, implying the onset of inflammatory lesions [29]. By 4–8 dps, severe villus detachment, necrosis, and submucosal and muscular layer damage were observed. A notable amelioration of intestinal morphology was observed at 16 dps. As a species capable of winter dormancy, Chinese soft-shelled turtles appear to activate intrinsic adaptive mechanisms to restore homeostasis, such as regulating stress-protective genes and mobilizing cellular repair systems [5]. Previous studies on hibernation in this species show that the intestinal villi remain structurally intact during dormancy, with the up-regulation of intestinal barrier proteins (ZO-1 and Cx43) and innate immune genes (Tlr2 and Tlr4) [30]. We deduced that similar adaptive mechanisms possibly contributed to the histological repair at 16 dps observed in our study. Generally, these results demonstrated the intestinal capacity to restore homeostasis following initial cold-induced damage.
Quantitative morphological analysis demonstrated significant reductions in several intestinal parameters during cold stress, including villus height and width, as well as submucosal width and muscularis layer width. These morphological alterations likely impaired intestinal barrier function, nutrient absorption, and immune defense, consistent with previous observations in yellow catfish [31] and zebrafish [24]. Notably, 7 °C treatment elicited more severe damage than 14 °C, suggesting a temperature-dependent nature of acute cold stress injury.
4.2. Effects of Low Temperature on the Intestinal Microbiome
The intestinal microbiome plays a pivotal role in maintaining intestinal functions, including digestion, nutrient absorption, and immune regulation. In aquatic species, temperature is a key environmental factor shaping microbial composition [32]. In the current research, α-diversity indices (Simpson, Shannon, and Pielou’s evenness) were significantly reduced, indicating decreased microbial diversity and evenness. Further analysis at the phylum level found that cold stress reduced the abundance of Proteobacteria and Bacteroidota—typical dominant phyla in healthy turtles—but increased Actinobacteriota. This result is similar to previous reports in teleosts [15] and the same turtle [33], implying that acute cold stress profoundly influenced the gut microbiota architecture of the Chinese soft-shelled turtle. At the genus level, potentially pathogenic taxa such as Helicobacter [34] and Citrobacter [35] were enriched in the 7 °C group, implying elevated disease risk under acute cold stress. The enrichment of Helicobacter and Citrobacter coincided with severe intestinal damage and metabolic disturbances, suggesting a possible pathological association that needs further validation. Functional analyses showed that microbiota lipid metabolism and carbohydrate metabolism were suppressed at 7 °C, while the energy and amino acid metabolism pathways were elevated, indicating a metabolic shift toward supporting host energy demands.
The relationship between host physiology and gut microbiota under cold stress might be bidirectional. On one hand, intestinal damage induced by acute cold stress, such as epithelial necrosis and inflammation, can alter the gut microenvironment, which promotes the growth of opportunistic pathogens and impairs host metabolic and immune function. This is consistent with seasonal studies of Chinese soft-shelled turtles, where winter restructuring of the gut microbiota enriches conditional pathogens such as Aeromonas and Edwardsiella. These alterations potentially contribute to post-hibernation disease susceptibility [36]. On the other hand, microbial changes may also reflect adaptation to cold stress. Intestinal villus atrophy under low temperatures reduces nutrient absorption capacity. Meanwhile, the microorganisms involved in “energy metabolism” and “amino acid metabolism” increased, suggesting that the microbiota may reallocate metabolic resources to restore host energy homeostasis. Similar adaptive metabolic alterations have been reported in cold-stressed Micropterus salmoides [15].
4.3. Functional Gene and Metabolite Screening Under Acute Cold Stress
Aquatic animals can modulate their immune systems in response to temperature fluctuations. As the first line of defense against pathogens [37], the innate immune system utilizes pattern recognition receptors (PRRs) to identify molecular signatures associated with pathogens or cellular damage, thereby triggering downstream immune responses [38]. Our findings demonstrated the significant down-regulation of multiple Toll-like receptor signaling pathway genes (Tlr2, Tlr4, Tlr5, Tlr7, and Tlr8) and NOD-like receptor signaling pathway genes (Traf6, Traf2, Casr, Rnasel, Pstpip1, Plcb2, Atg5, and Mfn2) in the 7 °C group, indicating that the pathogen recognition ability may be impaired. These results aligned with previous reports in tilapia, where genes encoding TLRs and NLRs are suppressed after 10 °C exposure [39]. In addition, genes involved in apoptosis (Tuba1c, Ctsz, Ctsb, Kras, Hras, Pik3ca, Bcl2l11, Gadd45a, Pmaip1, Ddit3, and Fos) and the p53 signaling pathway (Serpine1, Sesn2, Ccng2, Igf1, Mdm2, Gadd45a, Pmaip1, and Cdkn1a) were significantly upregulated in this research, indicating that cold stress induces intestinal apoptosis, corroborating the aforementioned histopathological damage.
Metabolomic analysis further revealed disturbances in lysine metabolism, with altered levels of key intermediates such as oxoglutaric acid, L-lysine, and L-aspartic acid. As a nutritionally important amino acid, lysine plays an important role in growth regulation and immune function in aquatic species [40]. Lysine supplementation is able to enhance organismal resilience to high-temperature stress [41]. Our study results indicate that lysine supplementation has the potential to mitigate acute cold-stress-induced metabolic disturbances. In addition, multiple intermediates of the TCA cycle showed significant changes, such as oxoglutaric acid, oxaloacetate, succinic acid, fumaric acid, DL-malic acid, citric acid (CA), and aconitate. CA is known to enhance gut health and alleviate oxidative stress in aquatic species [42,43,44]. Its decrease in our results indicates that restoring TCA cycle homeostasis through CA supplementation could be a potential strategy to mitigate intestinal damage under cold stress.
4.4. Crucial Pathways Responding to Low-Temperature Stress
By integrating DEG and DEM data, we identified three key pathways that mediate the intestinal response of the Chinese soft-shelled turtle to acute cold stress, including “linoleic acid metabolism”, “neuroactive ligand–receptor interaction”, and the “FoxO signaling pathway”.
In response to cold stress, poikilothermic vertebrates utilize several key strategies: modulating cell membrane fluidity, producing antifreeze proteins, and increasing the synthesis of molecular chaperones [45]. Linoleic acid, a key component of membrane phospholipids, contributes to maintaining membrane fluidity. This study revealed significant alterations in genes (Plb1, Cyp2j, and Cyp3a) and metabolites ((12Z)-9,10-Dihydroxyoctadec-12-enoic acid (9,10-DHOME), 13S-Hydroperoxy-9Z,11E-octadecadienoic acid (13(S)-HPODE), (12Z)-9,10-Dihydroxyoctadec-12-enoic acid (9,10-DHOME), (13S)-Hydroxyoctadecadienoic acid (13(S)-HODE), and 9(S),12(S),13(S)-Trihydroxy-10(E)-octadecenoic acid (9,12,13-TriHOME)) related to linoleic acid metabolism. Plb1 activation leads to the down-regulation of Cyp2j expression and the up-regulation of Cyp3a expression, resulting in a decrease in 12,13-DHOME and an increase in 9,10-DHOME. On the other hand, Plb1 can specifically catalyze the conversion of 13(S)-HPODE to 9,12,13-TriHOME, rather than 13(S)-HODE. This result is similar to previous reports on Hemiphyllodactylus yunnanensis [46] and Eremias argus [47], suggesting that the activation of linoleic acid metabolism to maintain cell membrane fluidity may represent a universal physiological mechanism in poikilothermic vertebrates for coping with cold stress.
Organisms eliminate injured cells via apoptotic processes to maintain normal physiological function [48]. The FoxO signaling pathway is a critical regulator of apoptosis [49]. In Brachymystax lenok tsinlingensis, FoxO suppresses apoptosis through the up-regulation of the anti-apoptotic gene Bcl-6. Similar regulatory patterns have been reported in cold-challenged carps [50] and tsinling lenok trout [51]. Our study observed increased mRNA levels of pro-apoptotic genes (Foxo4, Prkab, Bcl2l11, Bcl6, Bnip3, and Cdkn1a) and metabolites such as adenosine diphosphate (ADP) and adenosine monophosphate (AMP). ADP and AMP, as key intracellular energy-sensing molecules, regulate the expression of Prkab. The activation of Prkab regulates the expression of the Foxo4 gene, further initiating the transcription of pro-apoptotic genes such as Bcl6, Bnip3, and Bcl2l11. This pathway was also identified in the spleen of Chinese soft-shelled turtles under similar conditions [52], highlighting its systemic role in stress adaptation. Neuroactive ligand–receptor (NLR) interaction, essential for intercellular communication and stress signal transduction, significantly changed. Similarly, NLR activation has been documented in the brains of yellow drum [53], Coilia nasus [54], and gynogenetic mrigal carp [55] under cold stress. Additionally, intestinal NLR regulation has been observed in yellow pond turtles [56]. Key genes (Adcyap1, Adora2a, Adra2a, Agtr1, and Agtr2) and metabolites (serotonin, L-glutamic acid, L-aspartic acid, melatonin, and taurine) were remarkably altered, suggesting that NLR signaling might regulate neuronal activity to respond to acute cold stress in Chinese soft-shelled turtles.
In general, multi-omics data indicate that cold stress leads to the disruption of intestinal microbiota homeostasis, enrichment of potentially pathogenic bacteria, and suppression of microbial metabolic functions, particularly in lipid and carbohydrate metabolism. This dysbiosis, together with direct cold stress, triggers host metabolic reprogramming, characterized by altered TCA cycle intermediates, activated linoleic acid metabolism, and modified neuroactive ligand–receptor interactions. Concurrently, cold stress activates apoptosis and p53 pathways to clear and repair damaged epithelium, while suppressing immune recognition via TLR/NLR signaling. These coordinated responses ultimately lead to intestinal morphological damage, metabolic dysfunction, and weakened immunity.
4.5. Impact of Acute Cold Stress on Systemic Metabolism via the Gut–Liver Axis
Cold stress not only disrupts intestinal flora imbalance, but also synergistically disrupts intestinal and hepatic metabolism and function through the gut–liver axis. The intestinal microbiota and liver communicate bidirectionally via metabolites, immune signals, and barrier integrity, playing a pivotal role in low-temperature adaptation [57]. Spearman correlation analysis (Figure S3) between liver metabolites from our published paper [18] and intestinal microbiota revealed correlations between specific gut bacterial taxa and hepatic metabolites, suggesting the microbiota-mediated regulation of liver function.
Notably, Helicobacter abundance correlated strongly with uric acid, a biomarker and pathogenic driver of non-alcoholic fatty liver disease (NAFLD). It can promote reactive oxygen species (ROS) generation by NADPH oxidase activation, inducing oxidative stress and activating the NF-κB signaling pathway to upregulate inflammatory factors [58]. Similar mechanisms have also been reported in gut–liver axis injury models [59]. Citrobacter enrichment was correlated with elevated hepatic pro-inflammatory metabolites such as hypoxanthine and lysophosphatidic acid (LysoPA). Hypoxanthine accumulation reflects purine metabolic dysregulation and exacerbates hepatocyte oxidative damage by enhancing ROS production [60]. LysoPA, at high local concentrations, disrupts cell membranes and induces cytotoxicity [61]. These microbiota–metabolite–liver regulatory patterns align with observations in murine models [59,62]. Consistent with our previous histopathological damage in the liver, these findings suggest that specific bacteria may aggravate intestinal barrier damage and hepatic inflammation by releasing endotoxins or pro-inflammatory factors.
5. Conclusions
This study comprehensively investigated the effects of acute cold stress on intestinal health in Chinese soft-shelled turtles. Exposure to 14 °C and 7 °C induced significant intestinal damage, with more severe pathology at 7 °C. Cold stress also disrupted the gut microbiota and altered key genes and metabolites. Enriched pathways included “linoleic acid metabolism”, “neuroactive ligand–receptor interaction”, and the “FoxO signaling pathway”. Furthermore, cold-induced dysbiosis, marked by the enrichment of Helicobacter and Citrobacter, might exacerbate systemic injury via metabolite-mediated gut–liver axis crosstalk, highlighting this axis as central to the host’s cold stress response.
The findings of this study offer promising insights, yet certain limitations should be considered. These findings are preliminary due to limited omics sample sizes (transcriptomics n = 3, metabolomics n = 6 per group) and controlled laboratory conditions. Future studies should validate mechanisms in larger, field-relevant cohorts and address sex-specific adaptations in mature turtles. The inferred functional data from omics tools also require experimental confirmation. In addition, our findings reflect responses from a mixed-sex juvenile population without considering potential sex-specific adaptations. Future studies should investigate sex-dependent adaptive mechanisms to cold stress in Chinese soft-shelled turtles.
Our work provides a basis for three potential future applications: (1) developing feed additives from identified metabolites (citric acid and succinic acid) to alleviate cold stress; (2) using key differentially expressed genes in marker-assisted selection for cold-tolerant strains; and (3) targeting the gut–liver axis through microbiota modulation or metabolite supplementation to mitigate systemic cold injury in aquaculture.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Liang Y.X. Tian P. Lu Y.K. Qin Q. Wang Z.A. Xiong G. Wang X.Q. Hu Y.Z. Establishment and population genetic analysis of SNP fingerprinting of Chinese soft-shelled turtle (Pelodiscus sinensis)Aquac. Rep.20243810234010.1016/j.aqrep.2024.102340 · doi ↗
- 2Wang D. Gao H.Q. China Fishery Statistical Yearbook China Agriculture Press Beijing, China 202524
- 3Zhang Z. Chen B. Yuan L. Niu C. Acute cold stress improved the transcription of pro-inflammatory cytokines of Chinese soft-shelled turtle against Aeromonas hydrophila Dev. Comp. Immunol.20154912713710.1016/j.dci.2014.10.01025450906 · doi ↗ · pubmed ↗
- 4Pirhalla D.E. Sheridan S.C. Ransibrahmanakul V. Lee C.C. Assessing Cold-Snap and Mortality Events in South Florida Coastal Ecosystems: Development of a Biological Cold Stress Index Using Satellite SST and Weather Pattern Forcing Estuaries Coasts 2015382310232210.1007/s 12237-014-9918-y · doi ↗
- 5Reid C.H. Patrick P.H. Rytwinski T. Taylor J.J. Willmore W.G. Reesor B. Cooke S.J. An updated review of cold shock and cold stress in fish J. Fish Biol.20221001102113710.1111/jfb.1503735285021 · doi ↗ · pubmed ↗
- 6Scharsack J.P. Franke F. Temperature effects on teleost immunity in the light of climate change J. Fish Biol.202210178079610.1111/jfb.1516335833710 · doi ↗ · pubmed ↗
- 7Sun Z.Z. Tan X.H. Liu Q.Y. Ye H.Q. Zou C.Y. Xu M.L. Zhang Y.F. Ye C.X. Physiological, immune responses and liver lipid metabolism of orange-spotted grouper (Epinephelus coioides) under cold stress Aquaculture 201949854555510.1016/j.aquaculture.2018.08.051 · doi ↗
- 8Chu P. Wang S. Yu W. Wang A. Zong Y. Yin S. Zhao C. Wang T. The impact of extremely low-temperature changes on fish: A case study on migratory bony fishes (Takifugu fasciatus)Aquaculture 202457974015510.1016/j.aquaculture.2023.740155 · doi ↗