Targeting CDC42 Protects Mitochondrial Function through KLF2/HIF-1α/PINK1 Signaling in Acute Kidney Injury
Xue Zhou, Xian Fu, Yi-Wen Meng, Ping Dai, Qing Jiang, Hou-Hua Yin, Qing-Jin Pan, Ai-Zhi Lin, Kai-Di Ni, Zi-Guo Luo, Ru-Yu Liang, Yi-Yu Chen, Hai-Xin Yuan, Jun-Yan Liu

TL;DR
This study shows that targeting CDC42 helps protect kidney cells by improving mitochondrial function and reducing damage in acute kidney injury.
Contribution
The study reveals a novel role of CDC42 in AKI and identifies a new therapeutic strategy through the KLF2/HIF-1α/PINK1 pathway.
Findings
CDC42 inhibition reduces renal injury and preserves mitochondrial function in AKI models.
CDC42 suppression activates a KLF2/HIF-1α/PINK1 cascade that promotes mitophagy and reduces oxidative stress.
The KLF2/HIF-1α/PINK1 pathway is critical for protecting renal tubular epithelial cells from oxidative damage.
Abstract
Acute kidney injury (AKI) is a severe clinical syndrome strongly associated with mitochondrial dysfunction and oxidative stress, yet effective therapies remain elusive. Here, we identify cell division cycle 42 (CDC42) as a critical mediator of AKI. Analysis of human single-cell RNA sequencing (scRNA-seq) dataset revealed marked upregulation of CDC42 in renal tubular epithelial cells (RTECs), which was validated in murine models of cisplatin- and ischemia-reperfusion-induced AKI. Pharmacological inhibition, conditional knockdown, or genetic ablation of CDC42 significantly alleviated renal injury, preserved mitochondrial function, and reduced reactive oxygen species (ROS) both in vivo and in vitro. Mechanistically, transcriptomic analysis, bioinformatic analysis, dual-luciferase reporter assays, ChIP assays and cellular functional validation revealed that CDC42 suppression activated a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
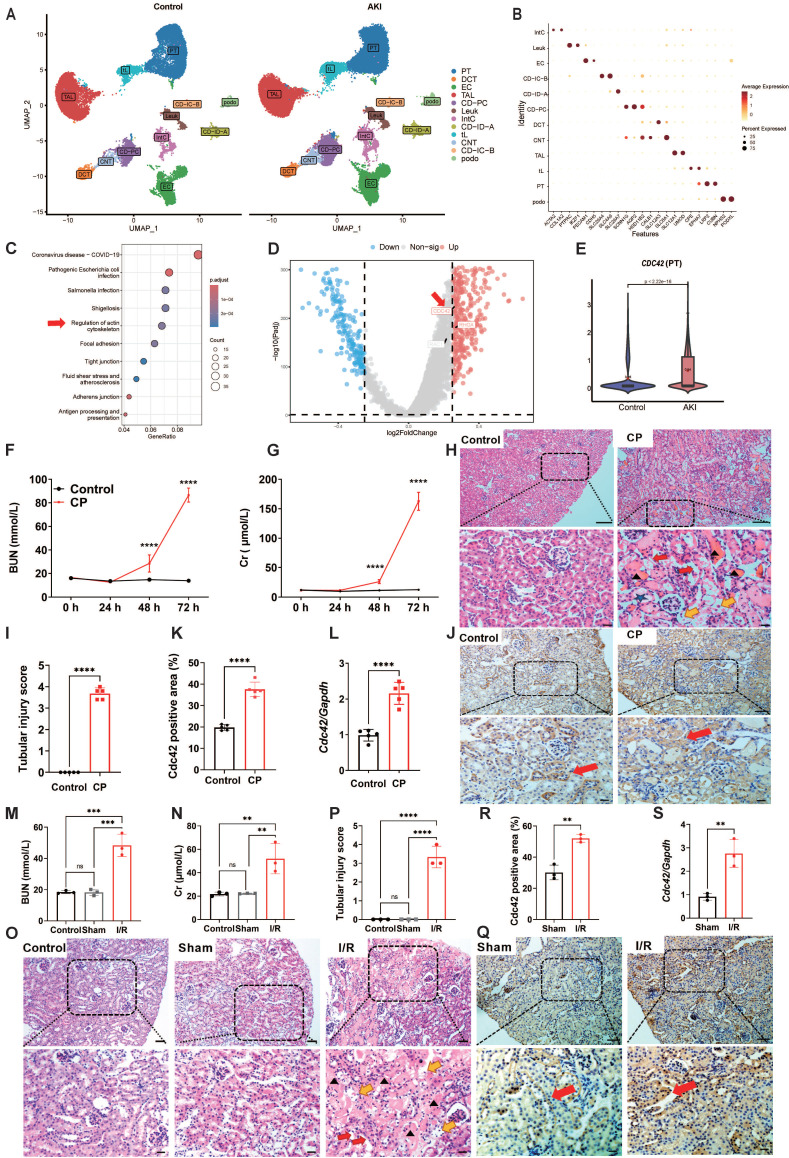 Figure 1
Figure 1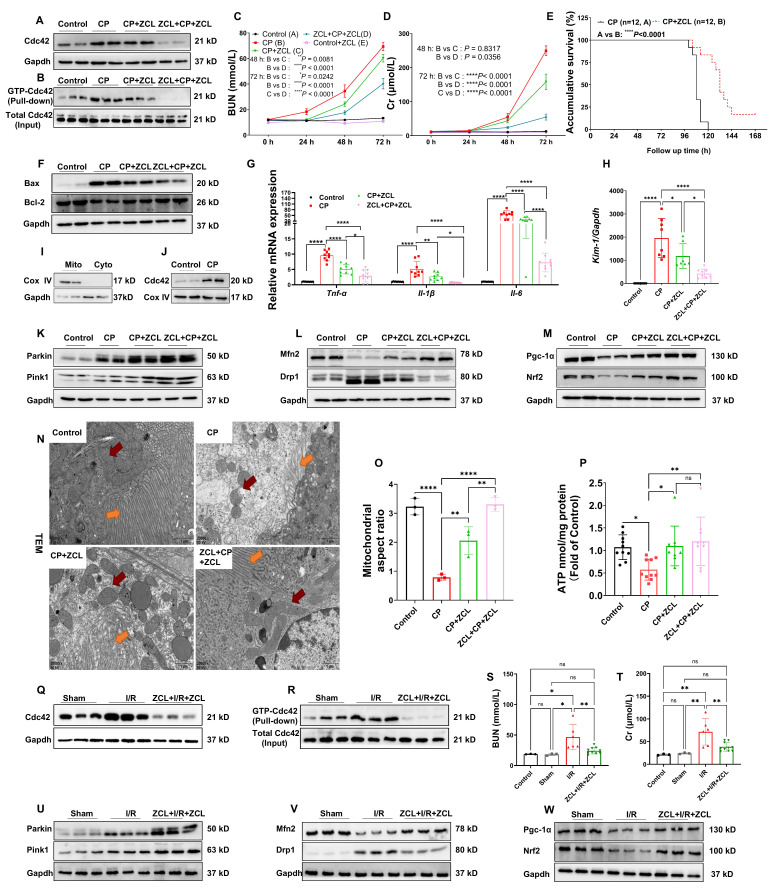 Figure 2
Figure 2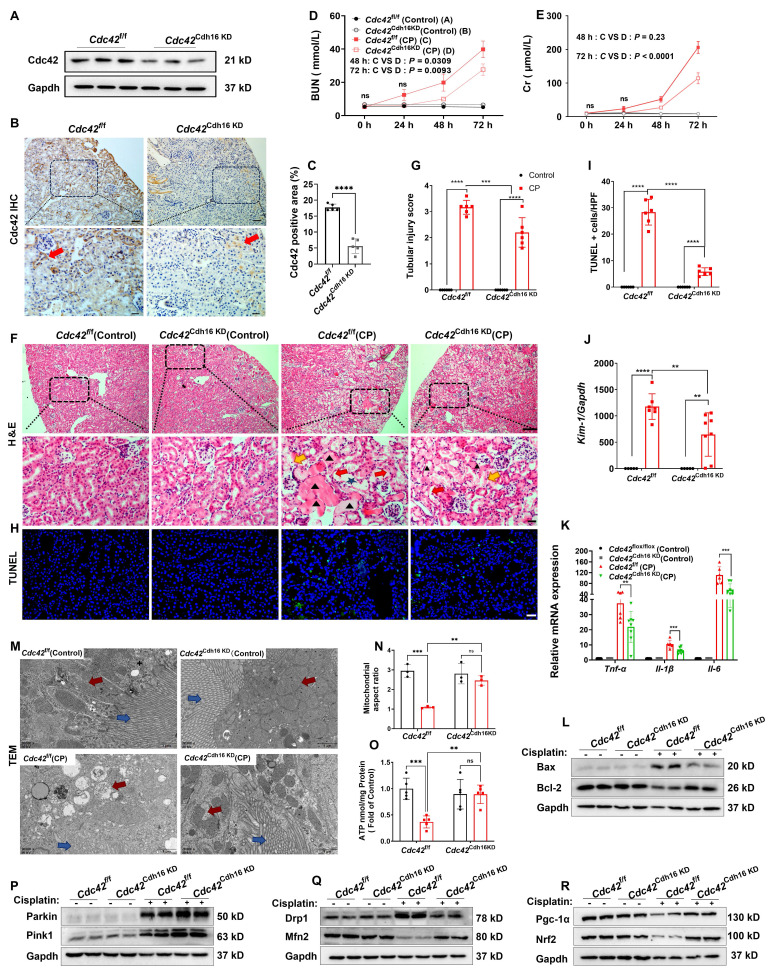 Figure 3
Figure 3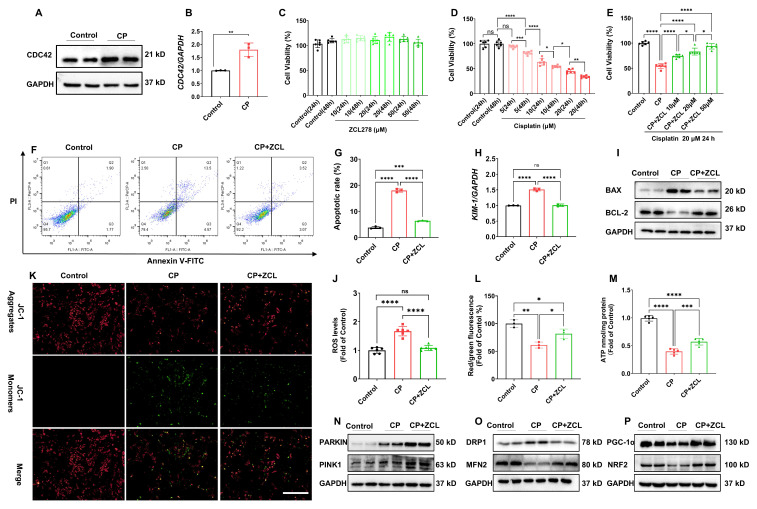 Figure 4
Figure 4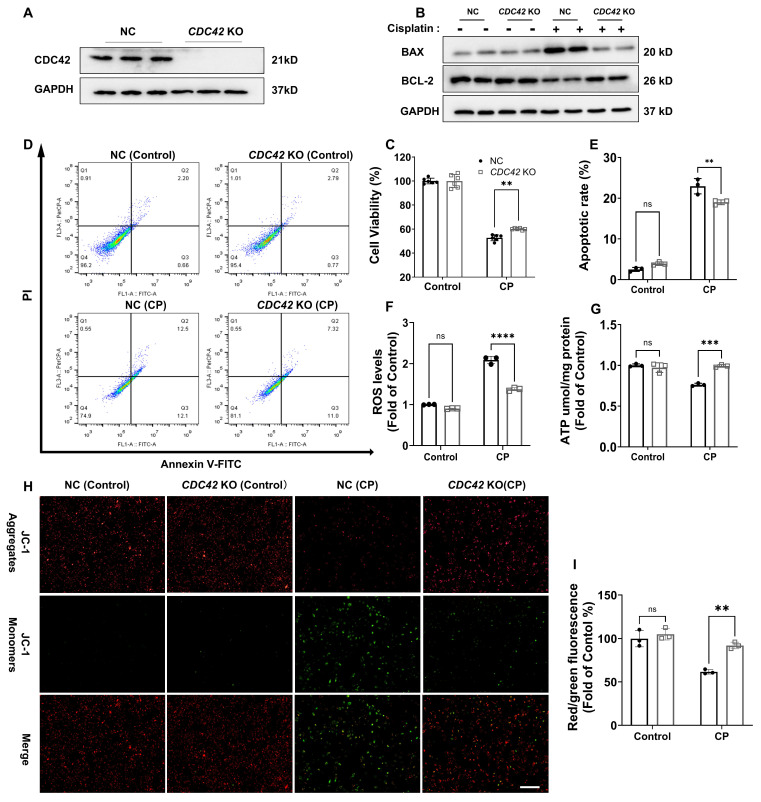 Figure 5
Figure 5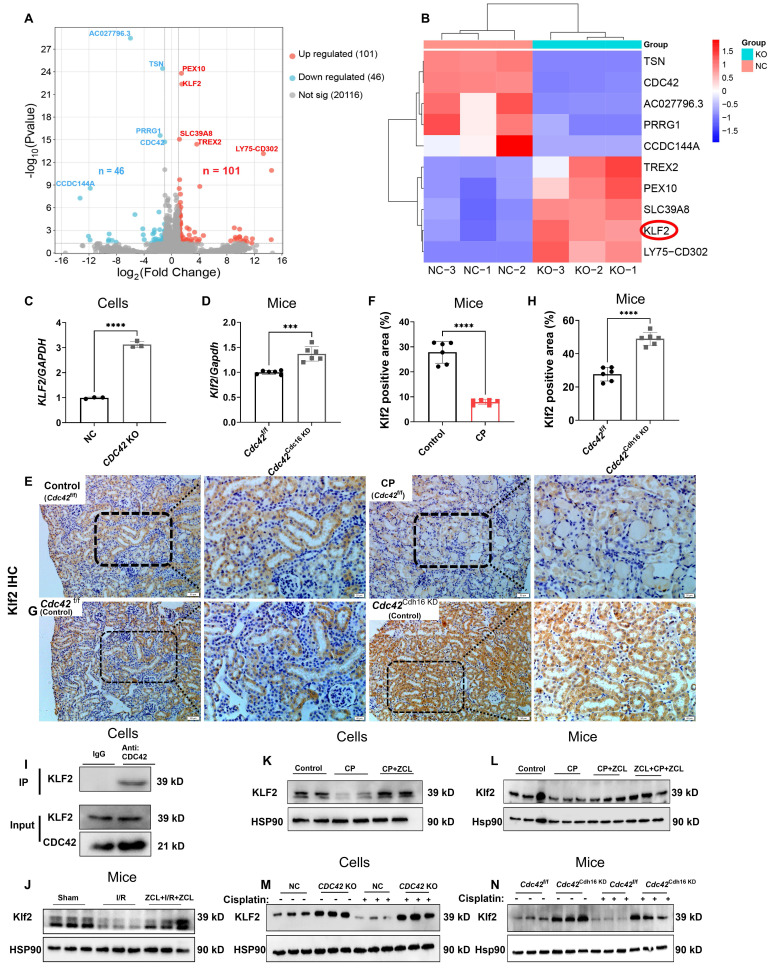 Figure 6
Figure 6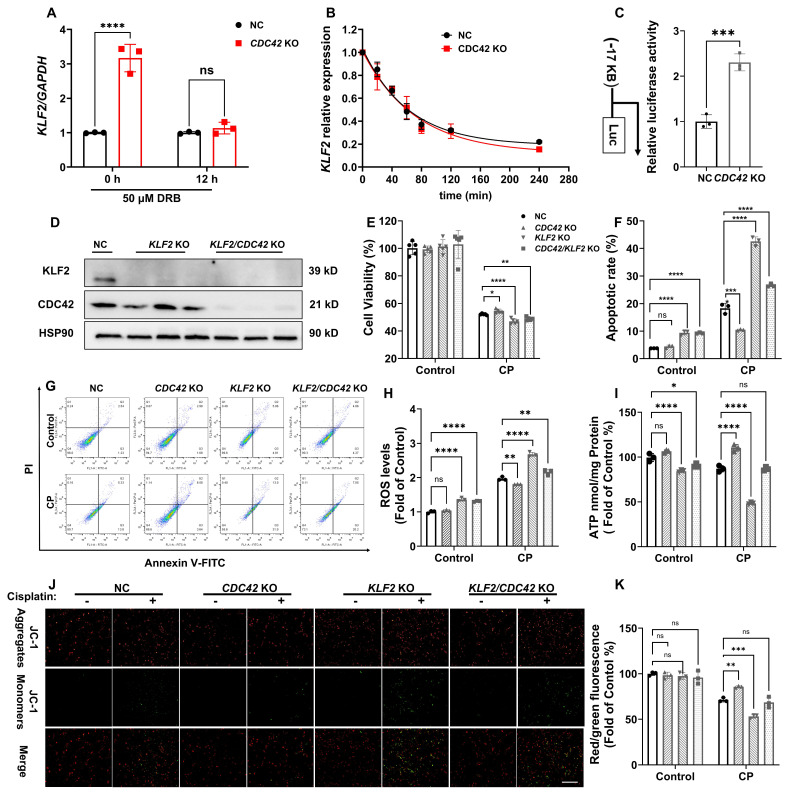 Figure 7
Figure 7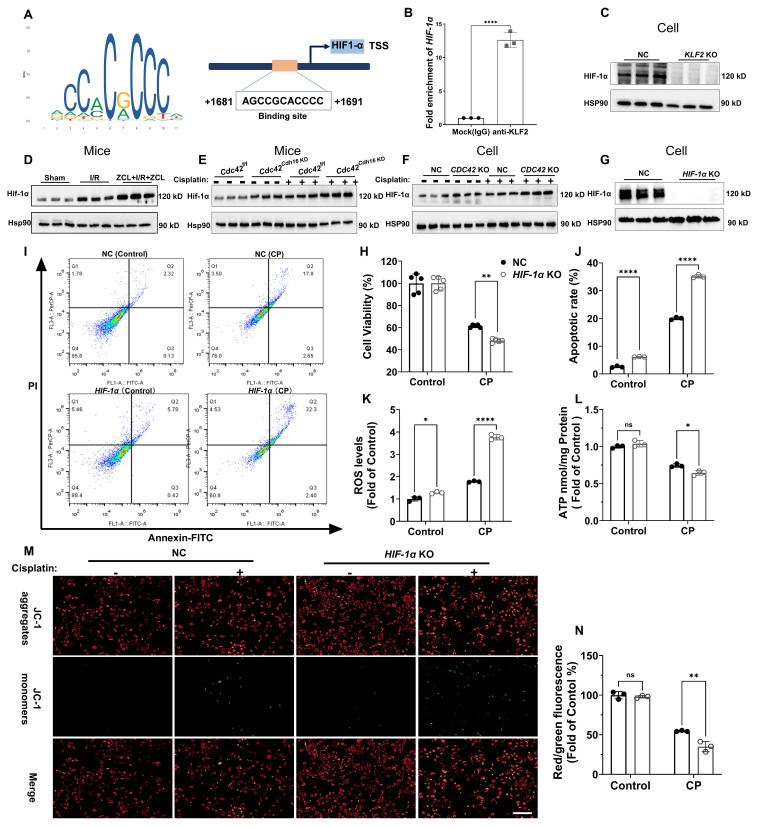 Figure 8
Figure 8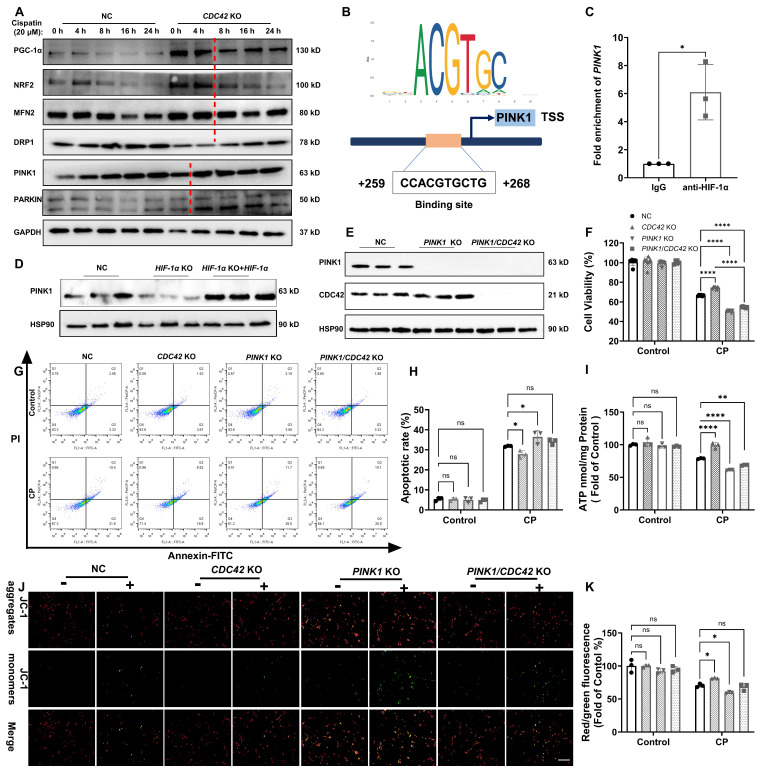 Figure 9
Figure 9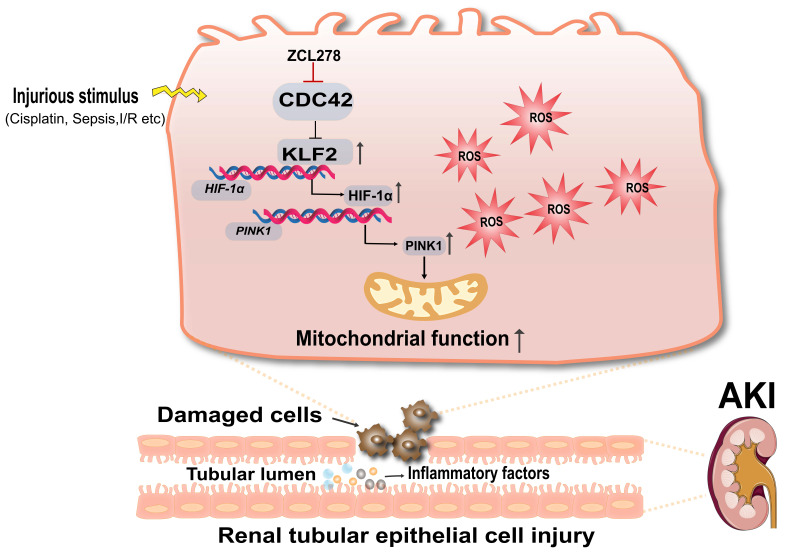 Figure 10
Figure 10Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAcute Kidney Injury Research · Kruppel-like factors research · Chemotherapy-induced organ toxicity mitigation
Introduction
Acute kidney injury (AKI), characterized by rapid deterioration of renal dysfunction, poses a critical healthcare challenge due to its high morbidity, persistently elevated mortality over decades, and substantial socioeconomic burden 1-3. Affecting 10-15% of hospitalized patients and over 50% of ICU admissions, AKI is strongly associated with adverse short-term outcomes, including a 5.6-fold higher 28-day mortality 4, and long-term risks of chronic kidney disease and mortality 5. Current management is largely limited to supportive care, highlighting the urgent need to explore novel prophylactic and therapeutic strategies.
Mitochondrial dysfunction and oxidative stress are central drivers of AKI pathogenesis, particularly in renal tubular epithelial cells (RTECs), which are highly metabolically active and rely on mitochondrial integrity to sustain reabsorption and energy production 6-9. Under AKI-related stress, mitochondria become dysfunctional and generate excessive levels of reactive oxygen species (ROS), which are estimated to contribute up to 90% of cellular ROS production 6,7,10. The resulting cellular ROS surge exacerbates mitochondrial damage and RTECs injury 1,8, creating a self-perpetuating cycle of oxidative damage and organ dysfunction. Therefore, strategy break this vicious cycle and restore mitochondrial homeostasis represent promising therapeutic avenues. However, the molecular mechanisms regulating redox-sensitive mitochondrial maintenance in RTECs remain incompletely understood.
In our ongoing effort to explore the new therapeutic strategies for kidney diseases 11-13, integrated analysis of published human single-cell RNA sequencing (scRNA-seq) data highlighted a potential role of cell division cycle 42 (CDC42). CDC42 is a key regulator of actin cytoskeleton dynamics, controlling essential cellular processes such as shape, migration, cell cycle progression and vesicle trafficking 14-16. Moreover, CDC42 functions as a cellular signaling switch, modulating multiple signaling pathways and governing diverse cellular functions 17. Its broad biological functions make it essential for renal development 18 and an attractive therapeutic target in several malignancies, including gastric 19, colorectal 20, and breast cancers 21,22. While CDC42 has been reported to be involved in mitochondrial fission 23, its role in mitochondrial homeostasis, particularly in redox balance during AKI, remains unknown.
Here, we report for the first time that CDC42 inhibition activates KLF2/HIF-1α/PINK1 signaling axis to promote mitophagy, restore mitochondrial function, and attenuate AKI by reducing oxidative stress. Using murine models of cisplatin- and ischemic-reperfusion (I/R)-induced AKI, as well as cisplatin-caused injury in human RTECs, we show that pharmacological and genetic suppression of CDC42 significantly attenuates mitochondrial dysfunction and renal injury. Collectively, these findings identify CDC42 as a previously unrecognized regulator of redox-sensitive mitochondrial homeostasis and offers mechanistic insights into its therapeutic potential in AKI.
Methods
Study approval
All animal experiments in this study were approved by the Animal Research and Ethics Committee of Chongqing Medical University (IACUC-CQMU-2023-11016) following the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Animals
Male mice (C57BL/6J, 8-weeks old) were purchased from GemPharmatech Co., Ltd. (Chengdu, China). Cdc42^flox/-^ and Cdh16-Cre mice, both with a C57BL/6 background, were purchased from the Shanghai Model Organisms Center, Inc. (Shanghai, China). Cdc42^flox/-^ mice were crossed with Cdh16-Cre mice using the Cre-loxP system to generate renal tubular epithelial-specific Cdc42 knockdown mice Cdc42^+/-^; Cdh16-Cre (Cdc42^Cdh16 KD^), and Cdc42^flox/flox^ mice (Cdc42^f/f^) were generated by mating Cdc42^flox/-^ mice with* Cdc42*^flox/-^ mice. The genotypes of mice were identified by PCR analysis of the DNA of tail tissues with the primer sequences presented in Supplementary Table 1.
Mice were housed in a specific pathogen-free (SPF) facility under controlled temperature (22 ± 1 °C) and humidity (55 ± 10%), with a 12-hour light/dark cycle, and ad libitum access to autoclaved water and food. After a one-week acclimatization, mice were randomly assigned to experimental groups as detailed in Figures 1, 2, and 3. All experimental treatments were performed at consistent times of the day to minimize circadian influences on renal function.
Measurement of blood levels of creatinine and urea
The blood levels of creatinine and urea in mice were measured using a high-performance liquid chromatography-tandem mass spectrometry (HPLC-MS/MS) as previously described 11.
Plasmids, lentivirus short-hairpin RNA construction, and cell transfection
The stable knockout (KO) cell line was constructed using CRISPR-Cas9 technology. LentiCRISPRv2-sgRNA-CDC42 (Sense 5'-CACCGACAGTCGGTACATATTCCGA-3' and anti-sense 5'-AAACTCGGAATATGTACCGACTGTC-3'), lentiCRISPRv2-sgRNA-KLF2 (sense 5'-CACCGCGCGTCGTCGAAGAGACCGA-3' and anti-sense 5'-AAACTCGGTCTCTTCGACGACGCGC-3'), lentiCRISPRv2-sgRNA-HIF-1α (sense 5'-CACCGAGATGCGAACTCACATTATG-3' and anti-sense 5'-AAACCATAATGTGAGTTCGCATCTC-3'), lentiCRISPRv2-sgRNA-PINK1 (sense 5'-CACCGCGTGGACCATCTGGTTCAAC-3' and anti-sense 5'-AAACGTTGAACCAGATGGTCCACGC-3'), and the lentiCRISPRv2-sgRNA-NC were purchased from Bio-rabbit (China, Shanghai). Lentivirus was packaged in HEK293T cells with Lipofectamine 3000 Transfection Reagent (Invitrogen, #L3000008) according to the manufacturer's instructions. After 48 h of infection, the supernatant of HEK293T cells was collected and used to infect HK-2 cells in the presence of polybrene (Beyotime; #C0351) to assist transfection. Transfected cells were then selected with puromycin (Beyotime; #ST551) to generate stable knockout cell lines. The knockout efficiency was evaluated by western blot (WB) analysis.
RNA sequencing (RNA-seq)
RNA sequencing was performed by Sangon Biotech (China, Shanghai) following total RNA extraction from HK-2 cells using TRIzol reagent (Invitrogen, #15596018CN). For samples without biological replicates, read count data were normalized using the TMM method, followed by an analysis of variance analysis with DEGseq. For samples with biological replicates, differential expression analysis was conducted using DESeq. Significantly different genes were identified based on adjusted p value ≤ 0.05 and absolute log_2_ fold change (| log_2_FC |) ≥ 1.
Statistical analysis
Data analysis was performed using SPSS 27.0 (SPSS Inc., Chicago, IL), and figures were generated with GraphPad Prism version 9 (La Jolla, CA). Normality was assessed using the Kolmogorov-Smirnov, Anderson-Darling, D'Agostino-Pearson omnibus, or Shapiro-Wilk test. Differences between two groups were analyzed using an independent-sample t-test, while One-way or two-way analysis of variance (ANOVA), followed by Tukey′s or Games-Howell post-hoc comparison, was used for multiple group comparisons. * p ≤ 0.05, ** 0.001 <* p* ≤ 0.01, *0.0001 < p ≤ 0.001 and **** p ≤ 0.0001 were considered significant difference.
Additional experimental details are presented in the Supplementary Material.
Results
Human scRNA-seq data analysis identified CDC42 as a pivotal gene in AKI pathogenesis
To explore the potential pathogenic mechanisms of AKI, we first downloaded raw data of human AKI from published scRNA-seq databases 24. After performing dimensionality reduction and clustering analysis (Figure 1A), and annotating cell types based on canonical markers (Figure 1B), we identified 12 major clusters from 96527 cells between control and AKI groups. Renal proximal tubule (PT) cells have become a major target for exploring AKI in recent years due to their abundance and heightened susceptibility/responsiveness to external insults 25,26. KEGG pathway enrichment analysis of PT cells from control and AKI groups revealed that differentially expressed genes (DEGs) were significantly enriched not only in infection-related pathways that associated with mortality etiology in AKI patients, but also significantly in actin cytoskeleton signaling pathways, which particularly captured our attention (Figure 1C).
The actin cytoskeleton, one of the most critical intracellular scaffold structures, maintains cell morphology, motility, and division through diverse actin isoforms and regulatory proteins governed by intricate signaling networks 27,28. Researches have demonstrated its involvement in mitochondrial homeostasis regulation 29. Its assembly/disassembly is primarily regulated by the Rho GTPases family, consisting of RHOA, RAC1, and CDC42, which coordinates different cellular processes through downstream effectors 30. Further volcano analysis in PT cells of control and AKI groups revealed that CDC42 is the most significantly altered gene in Rho GTPases family (Figure 1D). Violin plot also showed that* CDC42* expression was significantly upregulated in PT cells of the AKI group (Figure 1E). Taken together, the scRNA-seq data analysis of human AKI suggested CDC42 may be a pivotal gene involved in the pathogenesis of AKI.
CDC42 was upregulated in cisplatin (CP)- and ischemia-reperfusion (I/R)-induced AKI mouse models
To further define the role of CDC42 in AKI, we adopted two most recognized and widely used murine models of CP- and I/R-induced AKI 31. In CP-induced AKI mice, blood urea nitrogen (BUN, Figure 1F) and creatinine (Cr, Figure 1G) were progressively increased and climaxed at 72 h post-exposure. Histologic analysis of kidney sections from the AKI mice (72 h post CP exposure) revealed severe tubular injury, as evidenced by hematoxylin-eosin (H&E) staining (Figure 1H and 1I). Immunohistochemistry (IHC) analysis showed that Cdc42 was abundantly expressed in mice kidney tissues, particularly in the renal tubular epithelial cells (RTECs) (including both PT cells and distal convoluted tubule (DCT) cells) (Figure 1J), which was consistent with the human AKI scRNA-seq results (Figure 1E and Supplementary Figure S1). Moreover, both IHC (Figure 1K) and q-PCR (Figure 1L) analyses indicated a significant upregulation of Cdc42 in the kidneys of CP-AKI mice. In I/R-induced AKI mice, renal injury was evidenced by significantly elevated BUN and blood Cr levels, and severe tubular damage (Figure 1M-1P). In addition, no significant difference in BUN, Cr, or tubular injury scores were observed between the control and the sham groups, suggested that surgical stress did not affect renal function. Similarly, IHC staining and q-PCR analyses revealed a significantly increased Cdc42 expression in the RTECs of I/R-AKI mice when compared with the sham controls (Figure 1Q-1S).
Cdc42 inhibition protected against kidney injury in CP-induced AKI mice
To investigate the therapeutic potential of Cdc42 inhibition in AKI, we administered ZCL278, a Cdc42-selective inhibitor, to AKI mice 32. In CP-induced AKI mouse model, two treatment regimens were designed: 1) post-treatment (CP+ZCL) to evaluate therapeutic efficacy, and 2) combined pre- and post-treatment (ZCL+CP+ZCL) to assess preventive and therapeutic efficacy. Notably, ZCL278 treatment effectively reduced renal Cdc42 protein expression (Figure 2A) and active GTP-Cdc42 levels (Figure 2B), significantly lowering elevated BUN and blood Cr concentrations in CP-AKI mice (Figure 2C and 2D). Importantly, ZCL278 showed no nephrotoxicity in healthy mice, as evidenced by stable BUN and Cr levels (Figure 2C and 2D). Furthermore, ZCL278 treatment prolonged the survival time of CP-induced AKI mice, with a median survival time of 108 hours in the CP group and 132 hours in the CP+ZCL group, indicating that ZCL278 has a sustained protective effect (Figure 2E). Meanwhile, H&E and TUNEL staining analyses (Supplementary Figure S2A-S2D), protein levels of Bax (a pro-apoptotic regulator) and Bcl-2 (an anti-apoptotic regulator) (Figure 2F), mRNA expression of kidney-injury-molecule-1 (Kim-1) and inflammatory cytokines (Il-1β, Il-6, and Tnf-α) (Figure 2G-2H) indicated that ZCL278 greatly attenuated kidney injury in CP-induced AKI mice. Given the concordance between mitochondrial (Supplementary Figure S3) and whole-cell (Figure 2F) Bax expression under CP challenge with or without ZCL278 treatment, subsequent studies used whole-cell Bax expression as an apoptotic marker.
Cdc42 inhibition alleviated mitochondrial dysfunction in CP-AKI mice
Cellular oxidative stress, induced by ROS released from mitochondrial dysfunction, is the central pathological mechanism in AKI 6,7,10. Given the crosstalk between actin cytoskeleton dynamics and mitochondrial function 29, we hypothesize that Cdc42 inhibition may protect against AKI by modulating mitochondrial function. Isolation of mitochondria from mouse kidneys (Figure 2I-2J) revealed that Cdc42 was increased in CP-AKI mouse kidney mitochondria, which indicating its involvement in AKI mitochondrial regulation. Mitochondria homeostasis relies on a dynamic balance among PGC-1α/NFR2-mediated mitochondrial biogenesis, MFN2/DRP1-mediated mitochondrial fusion and fission, and PINK1/PARKIN-mediated mitophagy 9, with impairment of any of these processes causing mitochondrial dysfunction. Notably, ZCL278 treatment effectively maintained mitochondrial homeostasis in CP-induced AKI mice (Figure 2K-2M). To further assess mitochondrial function, we characterized mitochondrial ultrastructure in CP-AKI mice with or without ZCL278 treatment. As illustrated in Figure 2N and 2O, Cdc42 inhibition with ZCL278 treatment significantly improved CP exposure-caused mitochondrial ultrastructural abnormalities. In addition, ZCL278-mediated Cdc42 inhibition rescued the compromised ATP generation (Figure 2P) in CP-challenged mice.
Taken together, pharmacological inhibition of Cdc42 via ZCL278 treatment restored mitochondrial function in CP-induced kidney injury. Both administration routes of ZCL278 provided protective effects, with combined pre- and post-treatment showing superior efficacy.
Cdc42 inhibition protected against kidney injury and mitochondrial dysfunction in I/R-induced AKI
To comprehensively evaluate the reno-protective effects of Cdc42 inhibition across etiologically distinct AKI models, we assessed renal injury and mitochondrial function in I/R-AKI mice following ZCL278 treatment. Consistent with results in CP-AKI mice, ZCL278 reversed I/R-induced increase in Cdc42 protein expression and its activation levels (Figure 2Q-2R), reduced elevated BUN and blood Cr concentrations (Figure 2S and 2T), and alleviated renal tubular injury (Supplementary Figure S2E and S2F). Additionally, ZCL278 treatment effectively preserved mitochondrial function (Figure 2U-2W).
Given the comparable efficacy in both CP- and I/R-AKI mouse models, combined with the higher clinical relevance of nephrotoxic AKI and the mouse mortality associated with I/R surgery, we primarily conducted subsequent animal experiments in CP-induced AKI model.
Conditional knockdown of Cdc42 in RTECs alleviated kidney injury and mitochondrial dysfunction in CP-induced AKI mice
To establish an appropriate renal conditional Cdc42 knockdown (cKD) mouse model to mimic CP-induced kidney injury, we first analyzed publicly available scRNA-seq data from control and CP-AKI mice 33 (Supplementary Figure S4A-S4B). The results revealed that the Nagl (a marker of AKI) 34,35 (Supplementary Figure S4C-S4E) and Cdc42 (Supplementary Figure S4F-S4H) expression in both PT and DCT cells were significantly increased in CP-AKI mice, with DCT cells showing injury severity similar to or greater than that of PT cells. CP exposure is known to impair glomerular filtration rate (GFR) primarily by damaging the distal nephron including DCT, connecting tubule and collecting ducts, and the S3 segment of the PT 36,37. This understanding is supported by previous studies 38,39 and our H&E staining (Figure 1H), which demonstrate that CP causes damage to both PT and DCT cells in AKI mice. Based on these findings, we selected Cdh16-Cre mice to generate a RTEC-specific Cdc42 conditional knockdown mouse model using the Cre-LoxP system, enabling targeted investigation of Cdc42 function in CP-AKI. The knockdown efficiency was verified by identification of mice genotyping (Supplementary Figure S5), WB assay (Figure 3A) and IHC staining (Figure 3B-3C).
Consistent with the nephro-protective profile of ZCL278 treatments, Cdc42 cKD in mice RTECs attenuated CP-induced increases in BUN and Cr levels (Figure 3D-3E), ameliorated histopathological damage (Figure 3F-3G), and decreased renal apoptotic cells in CP-AKI mice (Figure 3H-3I). Furthermore, inflammatory and apoptotic markers further supported these findings, as Cdc42 cKD mice exhibited lower Bax, Kim-1, Il-6, Tnf-α, and Il-1β, alongside higher Blc-2 expression (Figure 3J-3L). Further analyses revealed that Cdc42 cKD AKI mice exhibited improved mitochondrial morphology (Figure 3M-3N), significantly higher ATP levels (Figure 3O) and restored mitochondrial homeostasis (Figure 3P-3R).
Taken together, these results revealed that pharmacological and genetic blockade of Cdc42 improved mitochondrial dysfunction, contributing to its protection against AKI.
Inhibition of CDC42 suppressed CP-induced cell injury and improved mitochondrial dysfunction in vitro
We employed HK-2 cells, an immortalized RTEC as an in vitro model to investigate the direct role of CDC42 in AKI. In accordance with our in vivo findings, CDC42 mRNA and protein expression was significantly upregulated in CP-treated HK-2 cells compared with the controls (Figure 4A-4B). CCK-8 assays showed that ZCL278 treatment (up to 50 μM for 48 h) did not affect cell viability (Figure 4C), indicating its safety profile. As anticipated, CP exposure decreased cell viability concentration- and time-dependently (Figure 4D), with 20 μM CP for 24 h reducing viability to ~50% of control levels. These conditions were set for subsequent experiments to mimic AKI in vitro. Notably, ZCL278 restored CP-induced viability loss in a concentration-dependent manner (Figure 4E), accompanied by reduced apoptosis (Figure 4F-4G) and* KIM-1* expression (Figure 4H). Additionally, ZCL278 treatment reversed CP-induced changes in BAX and BCL-2 protein levels (Figure 4I).
Given that mitochondrial dysfunction is a primary source of ROS in AKI 6,7,10, we assessed ROS levels and found that ZCL278 treatment significantly reduced CP-induced ROS accumulation (Figure 4J). Furthermore, ZCL278 treatment restored ATP synthesis and mitochondrial membrane potential (MMP) (Figure 4K-4M), while WB analyses of mitochondrial homeostasis markers further confirmed that CDC42 inhibition mitigated CP-induced mitochondrial dysfunction (Figure 4N-4P).
Genetical knockout of CDC42 protected HK-2 cells against CP-caused cell injury and mitochondrial dysfunction
To further validate the protective role of CDC42 inhibition in AKI, we constructed a stable CDC42 knockout (KO) HK-2 cell line (Figure 5A). WB analysis revealed that CDC42 KO reversed CP-induced changes in BAX and BCL-2 protein expression in the negative control (NC) cells (Figure 5B). Also, CDC42 KO alleviated the CP-induced decline in cell viability (Figure 5C) and CP-induced increase in cell apoptosis (Figure 5D-5E). Additionally, CDC42 KO markedly mitigated mitochondrial dysfunction and alleviated cellular oxidative stress, as evidenced by a marked reduction in CP-induced ROS accumulation (Figure 5F) and restoration of MMP and ATP levels (Figure 5G-5I).
Taken together, these above in vitro cellular studies showed that inhibition of CDC42 protected HK-2 cells from ROS-induced cellular oxidative stress injury by ameliorating mitochondrial dysfunction, which further supported CDC42 as a potential therapeutic target for AKI.
KLF2 was a downstream target gene of CDC42 in regulating AKI
To elucidate the molecular mechanisms underlying CDC42-mediated renal protection in AKI, we performed RNA sequencing (RNA-seq) on CDC42 KO cells and NC cells. A total of 147 DEGs were identified (Figure 6A), while heatmap analysis (Figure 6B) highlighted the top 10 most significantly altered genes, among which KLF2 emerging as a key candidate due to its well-established roles in mitochondrial function 40,41, oxidative stress-mediated cell damage and apoptosis 42. This hypothesis was then supported by q-PCR, showing that both* CDC42* KO (in vitro) and cKD (in vivo) significantly increased KLF2 mRNA levels (Figure 6C-6D). Furthermore, IHC staining revealed abundant Klf2 expression in RTECs, which was decreased following CP exposure (Figure 6E-6F), but was restored in Cdc42^Cdh16 KD^ mice (Figure 6G-6H).
Critically, co-immunoprecipitation (Co-IP) assays indicated the protein interaction between CDC42 and KLF2 (Figure 6I). Importantly, CDC42 inhibition reversed the CP- and I/R-induced reduction in KLF2 expression in both in vivo (Figure 6J, 6L, 6N) and in vitro (Figure 6K, 6M) models, consistent with IHC results. This regulation axis was also supported by the fact KLF2 KO failed to affect the CDC42 expression (Figure 7D).
These findings identify KLF2 as a critical downstream target of CDC42 inhibition, providing novel insights into its protective role against AKI in mitochondria and oxidative stress.
CDC42 KO increased KLF2 promoter activity and exerted its protective effect against kidney injury by mediating KLF2
To explore the mechanism underlying the regulation of KLF2 by CDC42, we examined KLF2 expression in the presence and absence of DRB (a potent RNA polymerase inhibitor). Notably, the increased KLF2 mRNA level caused by CDC42 KO was completely abolished by DRB treatment (50 μM) (Figure 7A), indicating that CDC42-mediated KLF2 regulation occurs at the transcriptional level. Considering the mature mRNA abundance is influenced by both transcription rate and degradation 43, we first assessed the half-life of KLF2 mRNA in HK-2 cells. We observed that CDC42 KO caused a slight increase in KLF2 mRNA stability (NC: t_1/2_ = 43.6 ± 7.8 min vs CDC42 KO: t_1/2_ = 50.2 ± 11.1 min, p = 0.38) (Figure 7B), demonstrating that CDC42-mediated KLF2 primarily through transcriptional activation rather than mRNA stability. Then, dual-luciferase assay revealed that CDC42 KO significantly increased the KLF2 promoter activity (1.7-kB* KLF2*- Luc) by 2.5-fold, indicating that CDC42 KO promotes KLF2 transcription to increase its mRNA expression (Figure 7C).
To validate that CDC42 inhibition attenuates AKI via KLF2, we established KLF2 KO and KLF2/CDC42 double KO (dKO) cells (Figure 7D). Interestingly, KLF2 KO alone significantly aggravated the CP-induced decrease in cell viability and increase in cell apoptosis. Unexpectedly, the protective effects of CDC42 KO on CP-induced cell injury were virtually counteracted in CDC42/KLF2 dKO cells (Figure 7E-7G). Consistently, KLF2 KO exacerbated CP-induced mitochondrial dysfunction, as evidenced by increased cellular ROS concentrations, and decreased ATP and MMP levels. Strikingly, the ability of CDC42 KO to restore mitochondrial function was nearly abolished in CDC42/KLF2 dKO cells (Figure 7H-7K).
Collectively, these results show CDC42 exerts its protective effects against AKI by transcriptionally upregulating KLF2.
KLF2 regulated AKI mechanism through transcriptional regulation of HIF-1α
To elucidate the mechanistic link between KLF2 and AKI, we performed a transcription factor database search using JASPAR, TRRUST, and KnockTF, identifying hypoxia-inducible factor 1α (HIF-1α) as a key downstream target. HIF-1α encodes the alpha subunit of transcription factor hypoxia-inducible factor-1 (HIF-1), a key regulator of cellular and systemic homeostatic responses to hypoxia 44. JASPAR database revealed conserved KLF2-binding motifs in the HIF-1α promoter (Figure 8A), which was experimentally validated by chromatin immunoprecipitation (ChIP) showing significant HIF-1α promoter enrichment in anti-KLF2 group (Figure 8B). Functionally, KLF2 KO reduced HIF-1α protein expression (Figure 8C), whereas CDC42 inhibition via ZCL278 or genetic approaches (KO/cKD) upregulated HIF-1α in both CP- and I/R-AKI models (Figure 8D-8F).
To define HIF-1α' role in AKI, we generated HIF-1α KO cells and their corresponding rescue model (HIF-1α KO + HIF-1α cells) (Figure 8G and Supplementary Figure S6). As expected, HIF-1α KO worsened CP-induced cellular damage (Figure 8H-8J) and mitochondrial dysfunction (ROS↑, ATP↓, MMP↓; Figure 8K-8N), whereas the aforementioned damage was reversed in* HIF-1α* KO + HIF-1α cells (Supplementary Figure S6). These data collectively show that CDC42 inhibition ameliorates AKI through KLF2-dependent transcriptional activation of HIF-1α.
HIF-1α mediated mitochondrial function via transcriptional regulation of PINK1 in CP-induced AKI
Our study revealed that HIF-1α KO disrupts mitochondrial function, consistent with previous findings 45,46. To elucidate the molecular mechanism underlying HIF-1α's role in mitochondrial homeostasis, we investigated whether mitochondrial biogenesis, fusion/fission, or mitophagy is the primary regulatory process by which CDC42 inhibition against AKI.
Time-course analysis of mitochondrial-related proteins in CDC42 KO and NC cells exposed to CP revealed that mitophagy-related proteins (PINK1, PARKIN) rapidly increased at 4 h, while mitochondrial biogenesis (PGC-1α, NRF2) and fusion/fission (MFN2, DRP1) proteins showed significant changes at 8 h (Figure 9A). This suggests that mitophagy is the initial step in mitochondrial regulation upon CDC42 inhibition. Using the JASPER database, we identified* PINK1* as the most likely downstream gene of HIF-1α in mitochondrial-related genes (Supplementary Figure S7), validated by ChIP assay showing HIF-1α binding to the PINK1 promoter (Figure 9B-9C) and WB results indicating PINK1 expression is positively regulated by HIF-1α (Figure 9D). Collectively, these results support the conclusion that mitophagy is the key initial process in CDC42/KLF2/HIF-1α-mediated mitochondrial regulation.
PINK1 is a crucial sensor of mitochondrial damage, initiating mitophagy to protect cells from external stimulus-induced mitochondrial dysfunction 47. To confirm that CDC42 inhibition provided the protective effect against AKI through regulating KLF2/HIF-1α/PINK1 axis, we examined the impact of CP exposure in PINK1 KO cells and* PINK1/CDC42* dKO cells (Figure 9E). As expected, PINK1 KO further exacerbated CP-induced cell injury and mitochondrial dysfunction, and more importantly, PINK1 KO inhibited the protective effect of CDC42 KO against CP-AKI in vitro (Figure 9F-9K). These findings suggest that PINK1 plays a crucial role in CDC42-mediated mitochondrial protection, supporting the hypothesis that CDC42 inhibition mitigates AKI by promoting mitophagy through KLF2/HIF-1α/PINK1 regulatory axis.
Discussion
This study is the first to reveal that CDC42 inhibition significantly attenuated kidney injury in both CP- and I/R-induced AKI models. Mechanistically, we show that CDC42 inhibition protected RTECs in vivo and in vitro by mitigating mitochondrial dysfunction and oxidative stress via activation of the KLF2/HIF-1α/PINK1-mediated mitophagy pathway (Figure 10).
To identify candidate drivers of AKI, we first analyzed scRNA-seq data from human AKI kidney 24. KEGG enrichment revealed activation pathways related to infection and, notably, actin cytoskeleton dynamics. Given that the AKI patients developed the disease in the context of critical illnesses, severe infections, and systemic inflammation, the enrichment in actin cytoskeleton dynamics signaling pathway was particularly striking. Among the three primary regulators of actin cytoskeletal remodeling 30,* CDC42* exhibited the more pronounced dysregulation in PT cells compared to RHOA and RAC1, suggesting a central role in AKI. It is well known that CDC42 functions as a molecular switch to transduce upstream signals to downstream effectors 14-16,48, which renders it crucial for organ development 49 and disease pathogenesis 50,51, as evident by congenital defects and tumorigenesis in organ-specific knockout models 52. In oncology, CDC42 overexpression has been reported in gastric 19, colorectal 20, breast 21,22, and liver cancers 53-55, correlating with poor clinical outcomes and therapeutic resistance. Beyond cancer, CDC42 is indispensable for kidney development, as it is required for ciliogenesis in RTECs 18. Its deletion in podocyte induces congenital nephrotic syndrome 56, while in chronic kidney disease, it promotes vascular calcification and renal fibrosis 57. Despite these insights, its role in AKI pathogenesis stays unknown.
Our research showed CDC42 was upregulated in human AKI kidneys, murine models of CP- and I/R-induced AKI, and CP-treated HK-2 cells. The induction of Cdc42 was likely driven by the robust increase in inflammatory cytokines, particularly Il-6 58, which was markedly elevated (~60-120 fold) in AKI mice kidneys. Importantly, both pharmacological inhibition and genetic blockage of Cdc42 attenuated renal dysfunction (Cr, BUN, and Kim-1), tubular injury, and inflammatory cytokines (Il-6, Il-1β, Tnf-α) expression. Parallel in vitro studies further validated the reno-protective effects of CDC42 inhibition across experimental systems.
Mitochondrial dysfunction and the ensuing oxidative burst are well-established drivers of RTEC injury in AKI 6-9, however the potential role of CDC42 in this process remains elusive. Notably, accumulating evidences shows that actin cytoskeleton dynamics are essential for modulating mitochondrial morphology and function 59. Furthermore, recent studies indicate that glycogen synthase kinase 3β (GSK3β), a known downstream effector of CDC42 60, acts as a key regulator of mitochondrial dysfunction in kidney injury 61,62. Although the participation of GSK3β in CDC42-mediated AKI remains to be further investigated, the crosstalk between CDC42, cytoskeletal remodeling, and mitochondrial function suggests that CDC42 inhibition likely ameliorates AKI by regulating mitochondrial homeostasis. Our* in vivo* and in vitro studies further validated this hypothesis: CDC42 was upregulated in renal mitochondria during AKI, with its inhibition restoring mitochondrial function by rescuing ATP production, membrane potential, and ultrastructure, along with the coordinated restoration of key homeostasis regulators (PGC-1α, NRF2, MNF2, DRP1, PINK1, and PARKIN).
Transcriptomic analysis identified KLF2 as a key downstream target of CDC42. KLF2 is a zinc finger transcription factor known to mitigate oxidative stress, inflammation, and thrombus activation 42. Previous studies have shown that KLF2 activation preserves mitochondrial function and attenuates ROS-induced damage by promoting mitophagy 40,41,63. In our study, CDC42 inhibition upregulated KLF2 expression via promoter activation, more importantly, KLF2 deficiency abrogated the mitochondria protection afforded by CDC42 inhibition, which establishing KLF2 as an essential mediator of the CDC42-mitochondrial axis.
Further mechanistic exploration revealed that KLF2 regulates HIF-1α transcription. ChIP assay demonstrated HIF-1α is transcriptionally regulated by KLF2 at the predicted promoter binding motif, and WB analyses demonstrated that KLF2-driven HIF-1α activation in response to CDC42 inhibition. HIF-1α is a transcription factor with critical role in hypoxia and mitochondrial protection. Activation of HIF-1α has been found to protect against AKI 64-67 by mitigating mitochondrial dysfunction, oxidative stress, inflammation, apoptosis, and autophagy 46,68,69. In addition, multiple lines of evidence, including transcriptional database prediction, ChIP, WB, and rescue assay, validated PINK1 was transcriptionally modulated by HIF-1α at the predicted promoter binding sites. Functionally, PINK1 deletion exacerbated mitochondrial dysfunction and abrogated the protective effect of CDC42 inhibition, consistent with its role as a key mitophagy regulator 7,70-74. Together, these findings establish a KLF2/HIF-1α/PINK1 axis through which CDC42 inhibition promotes mitophagy and preserves mitochondrial homeostasis.
In addition to mitophagy, CDC42 inhibition dampened inflammatory responses in AKI kidneys, consistent with prior studies linking CDC42 to chronic inflammatory conditions 75-78. However, the loss of protection in KLF2 or PINK1 deficient models indicates that mitophagy-mediated mitochondrial preservation is the predominant protective mechanism, with anti-inflammation and other factors as the secondary outcome.
Our findings also highlight the translational potential of targeting CDC42, which is consistently upregulated in human and experimental AKI, with its inhibition conferring robust renoprotection across models. As a multifunctional signaling hub, CDC42 offers a distinct mechanistic advantage over downstream-specific strategies such as soluble epoxide hydrolase (sEH) inhibitors (anti-inflammatory) 79 or HIF-PH inhibitors (hypoxia adaptation) 80, as well as single-target approaches like PGC-1α agonists 81. By coordinately modulating key processes, including inflammation (Figure 2G, Figure 3K), oxidative stress (Figure 4J, Figure 5F), and mitochondrial function (Figure 2K-2P, Figure 2U-2W, Figure 3M-3R), CDC42 inhibition elicits synergistic effects that enable more comprehensive restoration of mitochondrial homeostasis. The selective inhibitor ZCL278, which disrupts CDC42-Intersectin interaction to suppress CDC42-mediated signaling 32, showed preventive and therapeutic efficacy without nephrotoxicity in CP- and I/R-induced AKI. These effects were further validated in RTEC-specific Cdc42 knockdown mice, minimizing concerns regarding off-target actions and supporting CDC42 as a highly specific therapeutic target. Moreover, ZCL278 showed efficacy through multiple routes of administration, underscoring its preclinical feasibility 82. Although clinically approved CDC42-specific inhibitors are not yet available, several dual Rho GTPases inhibitors, such as R-ketorolac and R-naproxen 50,83,84, mitoxantrone 85, and MBQ-167 86 have progressed into oncology trails. These developments suggest that targeting CDC42 in AKI is both feasible and promising, warranting further exploration.
In addition, we acknowledge that the present study has certain limitations. First, our AKI models are based on rodents, and their translational relevance to human clinical settings needs further verification using more diverse clinical AKI samples. Second, the molecular mechanism underlying CDC42-mediated enhancement of KLF2 promoter activity remains to be fully elucidated, as potential intermediate molecules may be involved.
In conclusion, our study identifies CDC42 as a pivotal regulator of AKI pathogenesis through its impact on mitochondrial function. By activating the KLF2/HIF-1α/PINK1 axis, CDC42 inhibition promotes mitophagy, restores mitochondrial homeostasis, and alleviates oxidative stress and renal injury. These findings provide mechanistic insight and translational rationale for developing CDC42-targeted and mitochondria-directed therapies for AKI.
Supplementary Material
Supplementary methods, figures and tables.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kellum JA Romagnani P Ashuntantang G Ronco C Zarbock A Anders HJ Acute kidney injury Nat Rev Dis Primers 2021 July 15;7(1):5210.1038/s 41572-021-00284-z 34267223 · doi ↗ · pubmed ↗
- 2Moore PK Hsu RK Liu KD Management of Acute Kidney Injury: Core Curriculum 2018 Am J Kidney Dis 2018 July;72(1):136-4810.1053/j.ajkd.2017.11.02129478864 · doi ↗ · pubmed ↗
- 3Uchino S Kellum JA Bellomo R Doig GS Morimatsu H Morgera S Acute renal failure in critically ill patients: a multinational, multicenter study JAMA 2005 Aug 17294781381610600610.1001/jama.294.7.813 · doi ↗ · pubmed ↗
- 4The Beijing Acute Kidney Injury Trial (BAKIT)workgroup Jiang L Zhu Y Luo X Wen Y Du B Epidemiology of acute kidney injury in intensive care units in Beijing: the multi-center BAKIT study BMC Nephrol 2019 Dec 2014683184278710.1186/s 12882-019-1660-z PMC 6915890 · doi ↗ · pubmed ↗
- 5Fortrie G De Geus HRH Betjes MGH The aftermath of acute kidney injury: a narrative review of long-term mortality and renal function Crit Care 2019 Dec 231243067869610.1186/s 13054-019-2314-z PMC 6346585 · doi ↗ · pubmed ↗
- 6Emma F Montini G Parikh SM Salviati L Mitochondrial dysfunction in inherited renal disease and acute kidney injury Nat Rev Nephrol 2016 May 125267802680401910.1038/nrneph.2015.214PMC 5469549 · doi ↗ · pubmed ↗
- 7Tang C Cai J Yin XM Weinberg JM Venkatachalam MA Dong Z Mitochondrial quality control in kidney injury and repair Nat Rev Nephrol 2021 May 1752993183323539110.1038/s 41581-020-00369-0PMC 8958893 · doi ↗ · pubmed ↗
- 8Linkermann A Chen G Dong G Kunzendorf U Krautwald S Dong Z Regulated cell death in AKIJ Am Soc Nephrol 2014 Dec 251226897012492572610.1681/ASN.2014030262 PMC 4243360 · doi ↗ · pubmed ↗