Identification of OCT Family Genes in Tomato (Solanum lycopersicum) and Function of SlOCT20 Under Cold Stress
Rui Lv, Fulei Mo, Yuxin Liu, Huixin Zhang, Mingfang Feng, Peiwen Wang, Mozhen Cheng, Shusen Liu, Zhao Liu, Xiuling Chen, Aoxue Wang

TL;DR
This study identifies the SlOCT20 gene in tomatoes and shows it helps plants tolerate cold stress by reducing oxidative damage.
Contribution
The study discovers a new gene, SlOCT20, that positively regulates cold stress tolerance in tomatoes.
Findings
SlOCT20 is localized in the cell membrane and plays a role in cold stress resistance.
Silencing SlOCT20 reduces cold tolerance and increases oxidative stress in tomatoes.
Transcriptome and qRT-PCR analyses confirmed SlOCT20's key role in cold stress response.
Abstract
Tomato, as a vegetable crop that prefers warmth, has relatively high requirements for planting temperature. Cold stress often occurs in tomato cultivation, especially in early spring and late autumn. Cold stress not only affects the growth, development and yield of tomatoes, but in severe cases, it can also lead to the direct death of tomato plants. Screening cold-resistant genes and analyzing their regulatory mechanisms of cold resistance are of great significance for enhancing the pertinence of stress-resistant breeding. This study identified the cold-resistant gene SlOCT20 in cultivated tomatoes and analyzed the molecular mechanisms by which it regulates tomato cold resistance, providing a new regulatory mechanism of plant cold resistance. Plant organic cation transporters (OCTs) are involved in a variety of beneficial biological processes, such as cadaverine transfer in plants and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
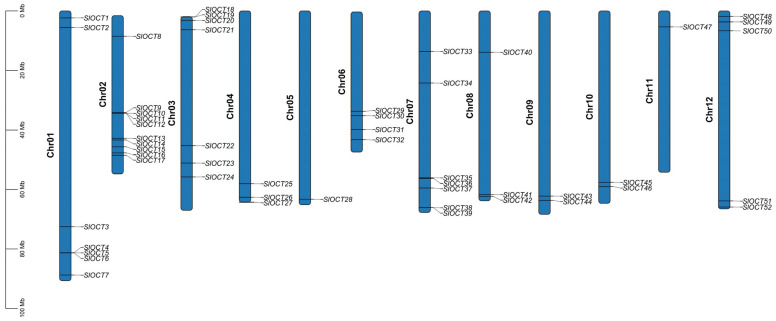 Figure 1
Figure 1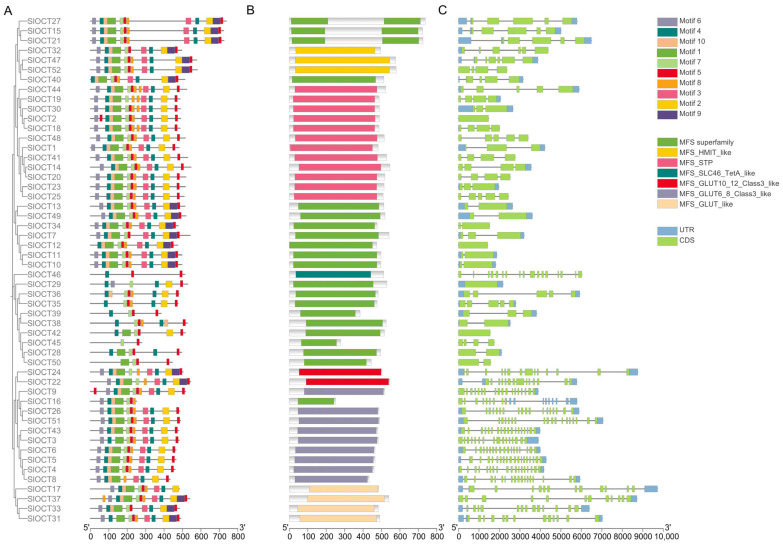 Figure 2
Figure 2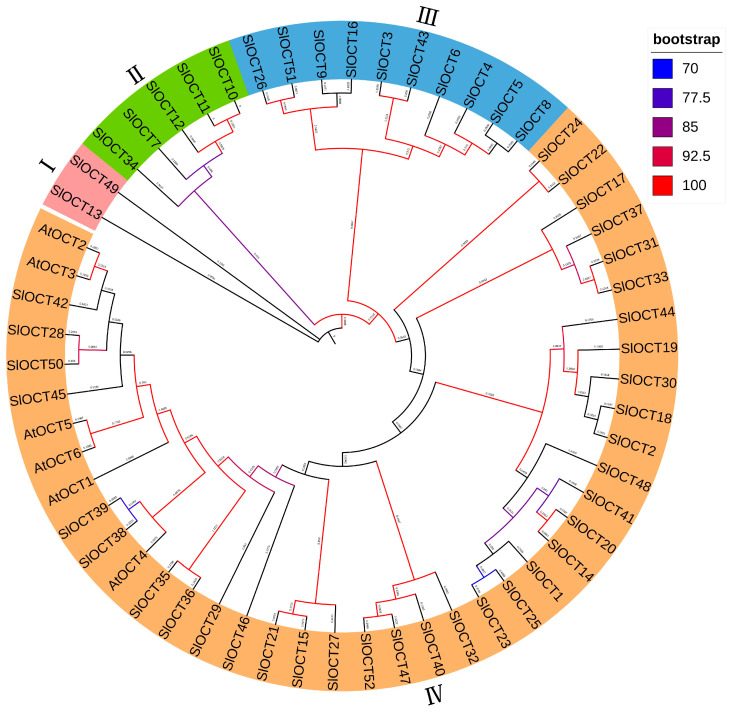 Figure 3
Figure 3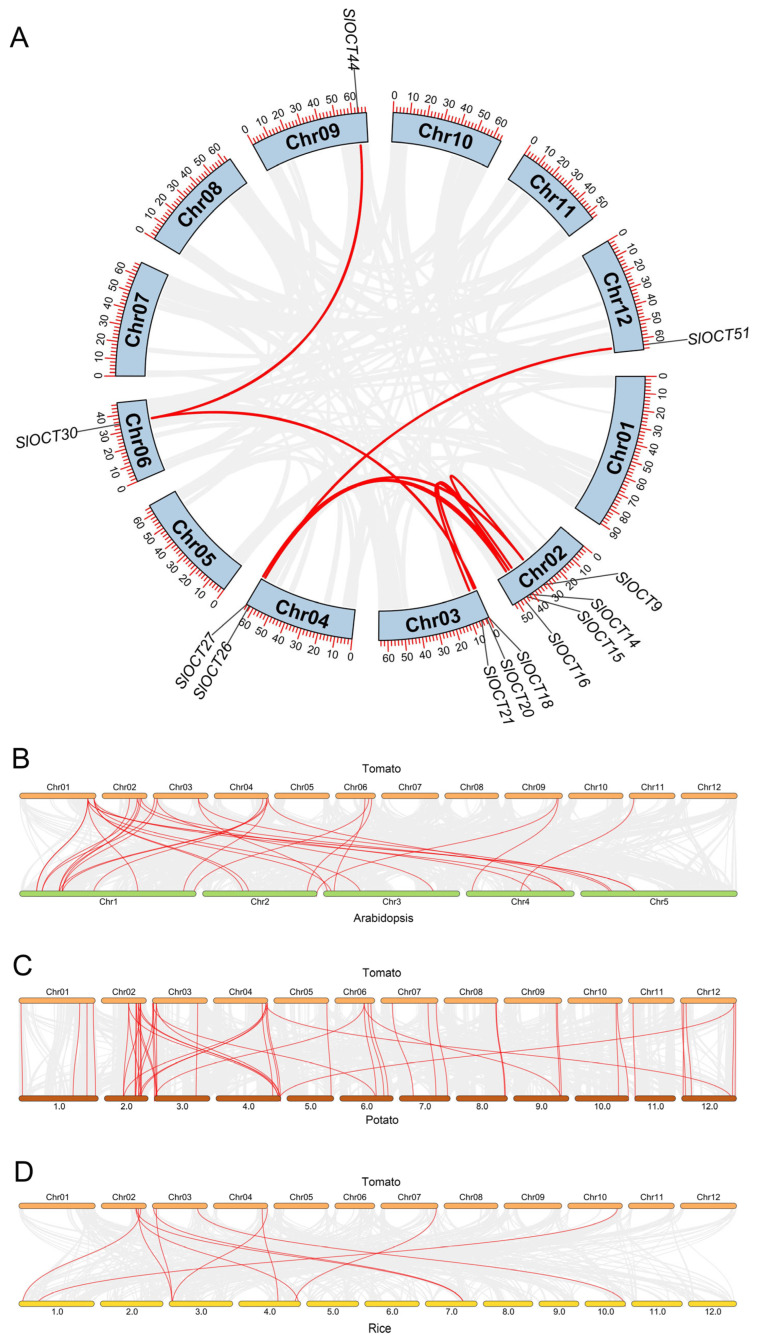 Figure 4
Figure 4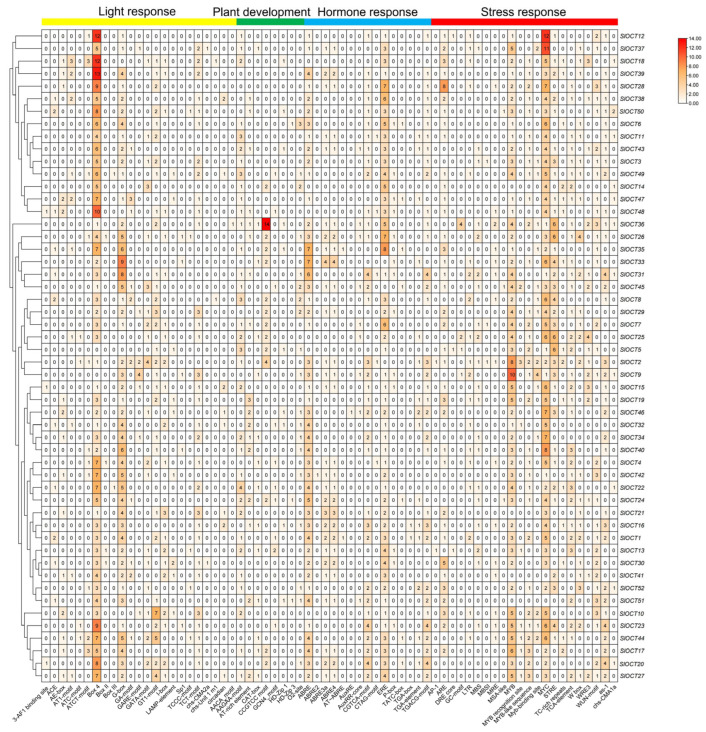 Figure 5
Figure 5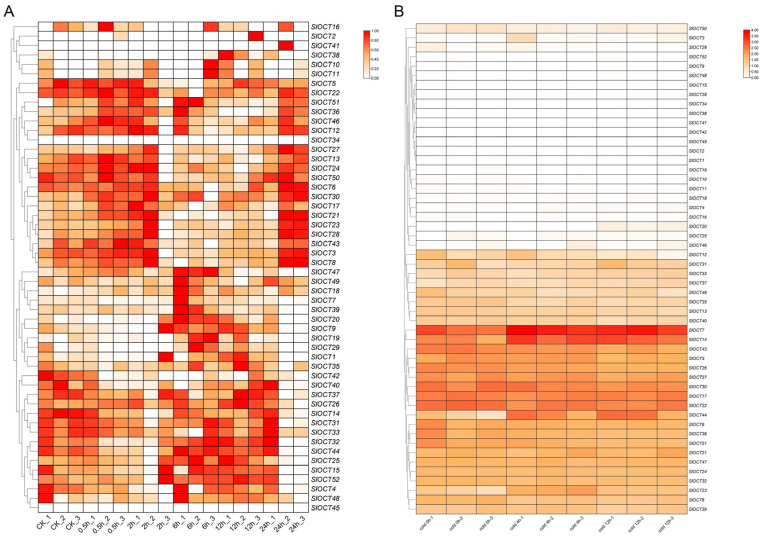 Figure 6
Figure 6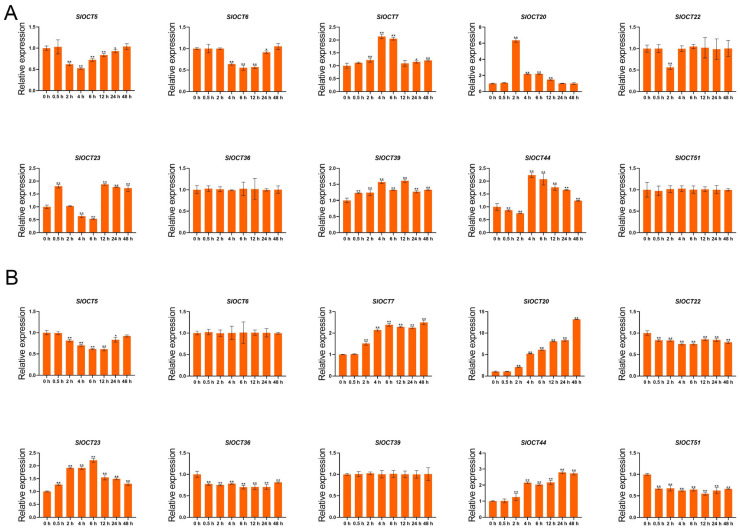 Figure 7
Figure 7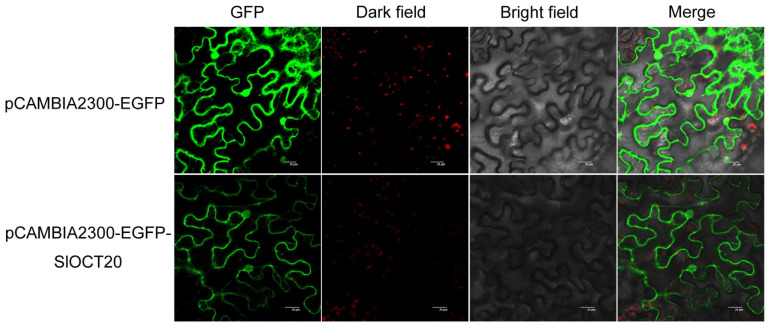 Figure 8
Figure 8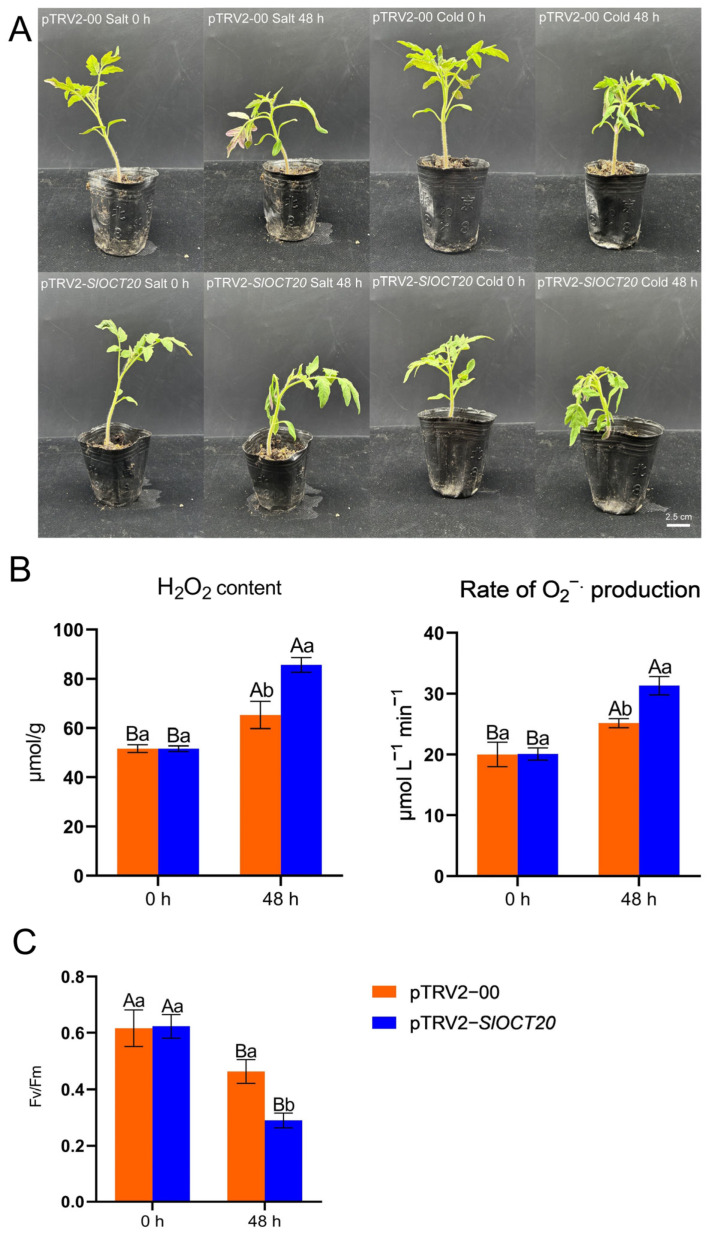 Figure 9
Figure 9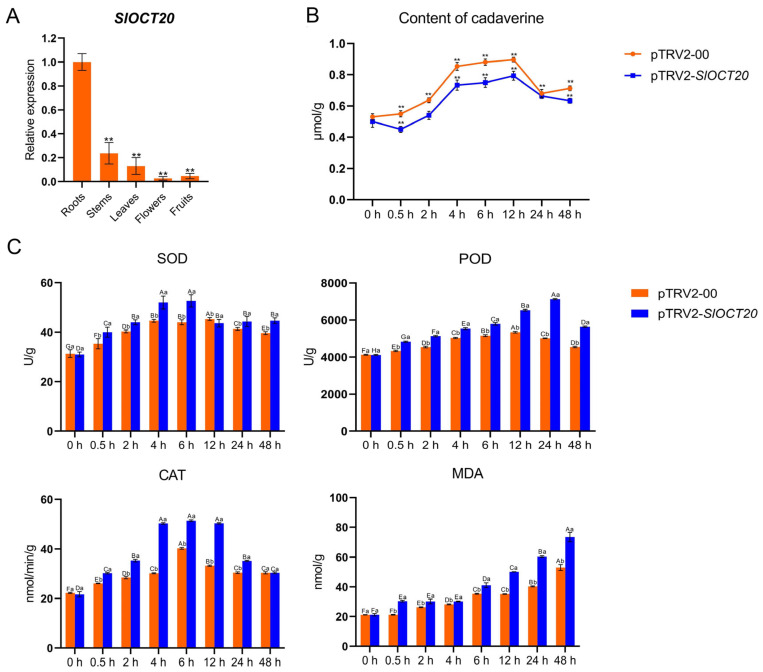 Figure 10
Figure 10- —National Key R&D Program of Shandong
- —National Natural Science Foundation of China
- —Taishan Industry Leading Talents Project Special Funds
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Stress Responses and Tolerance · Plant-Microbe Interactions and Immunity · Plant responses to water stress
1. Introduction
In complex environments, plants have evolved multiple mechanisms to adapt to stress. The transmembrane transport of cations plays a significant role in this process, including mineral absorption, osmotic pressure regulation, cell expansion, environmental and cellular signal transduction and stress tolerance [1]. OCTs are involved in a variety of biological processes. OCT proteins are not only present in animals but also widely found in higher plants.
OCT family proteins belong to the Solute Carrier 22A (SLC22A) superfamily and are involved in the transmembrane transport of organic cations [2]. The first reported OCT gene in plants was identified in Phasolus vulgaris, which is induced by drought and other stresses, but its substrate remains unknown [3]. In Arabidopsis thaliana, BOU is proposed to encode a mitochondrial acetylcarnitine transporter, and OCT1 has been shown to repress BOU transcription. The Arabidopsis oct1 mutant showed an altered root sensitivity to carnitine, with higher lateral root density in the oct1 mutant than in the wild type. Yeast mutant transformation experiments revealed that AtOCT1 may function as a carnitine transporter [4]. AtOCT1 protein is localized to the cell membrane, and is expressed in vascular tissues and lateral root initiation sites. AtOCT1 promoter activity has also been detected in the vascular tissues of roots, leaves and flowers, suggesting that AtOCT1 may play a role in transport in plants [5].
Soil contains organic cations such as cadaverine cation and carnitine zwitterion, and plants can synthesize carnitine and cadaverine under stress conditions [6,7]. The above organic cations in plants play an active role in plant development and stress adaptation [8]. Studies on the absorption and transport mechanism of beneficial organic cations in plants can deepen the theory of plant stress resistance. In this study, 52 OCT family genes were identified using the tomato genome version 4.0. The intron and exon distribution of all OCT family genes in tomato was visualized. The conserved structures and features of all OCT proteins were characterized. By constructing the OCT family protein phylogenetic tree and analyzing the collinearity relationship among OCT family genes, the evolutionary laws of OCT family genes in plants can be understood. In addition, the cis-acting elements of tomato OCT family genes in their promoters were statistically analyzed, and the expression patterns of tomato OCT family genes under salt and cold stress were clarified by transcriptome data analysis and qRT-PCR experiments. The function of SlOCT20 in cold stress was resolved by silencing the SlOCT20 gene. This study elucidated the evolutionary and functional differentiation of OCT family genes in tomato and clarified the association between OCT family genes and cold stress tolerance in tomato.
2. Materials and Methods
2.1. Planting Environment and Different Stress Treatments
The tomato variety used in this study is ‘Ailsa Craig’, and the tobacco (Nicotiana benthamiana) seeds were stored and provided by our laboratory. Seeds were sown in fertile soil and grown in the plant incubator. The soil composition consisted of 80% mass of uncontaminated field soil (obtained from Xiangyang Farm, Northeast Agricultural University, Harbin, China) and 20% mass of perlite, which was mixed and sterilized at high temperature for use. The sterilized soil was loaded into a 200 mL capacity plastic flowerpot, and the seeds were soaked in distilled water for 12 h before sowing. Plant incubator parameters: (illumination time: 15.5 h, illumination intensity: 25,000 lux, parameter of temperature: 25.5 °C; time of night: 8.5 h, parameter of temperature: 19.5 °C). When tomatoes reached 4 weeks of age, they were subjected to salt and cold stress treatments. The concentration of NaCl in the salt stress treatment was 100 mmol/L, the treatment was carried out by root irrigation (single irrigation, 200 mL per tomato plant) and the control group was treated with the same volume of distilled water. Cold stress treatment parameters: (illumination time: 15.5 h, illumination intensity: 25,000 lux, parameter of temperature: 4.0 °C; time of night: 8.5 h, parameter of temperature: 4.0 °C). The temperature of the control group was 25.5 °C during the day and 19.5 °C during the night, and other parameters were the same as those of the cold treatment group. The cooling mode of the cold stress treatment was direct cooling, that is, the plants were directly transferred to 4 °C. The control group of the VIGS experiment was tomato transfected with pTRV2-00 empty vector, and the stress treatment was the same as above. Tomato tissue samples were removed and snap frozen in liquid nitrogen for three biological replicates at 0 h, 0.5 h, 2 h, 4 h, 6 h, 12 h, 24 h and 48 h of stress, and these samples were used for subsequent experiments.
2.2. Identification of OCT Family Members in Tomato and Characters of OCT Proteins
Known OCT protein sequences in Arabidopsis were obtained from the TAIR database (https://www.arabidopsis.org/, accessed on 17 January 2025). The tomato genome file (version 4.0; from cultivated tomato Heinz 1706) and its annotation file were obtained from the Phytozome database (http://phytozome.jgi.doe.gov/pz/portal.html, accessed on 19 January 2025). All tomato DNA sequences were translated, and the resulting protein file was used as a library, in which BLAST proteins similar to the Arabidopsis OCT protein were identified using the BLASTp plugin in TBtools (v2.138), E value = 1 × 10^−5^. The HMM profile (PF00083) for the OCT protein family, retrieved from the Pfam database (accessed on 26 January 2025), was employed to screen the tomato protein sequences using TBtools software (v2.138) [9]. The threshold parameter is the default of the software. The intersection of the search results of BLAST and HMM was taken as the OCT family members in tomato. Tomato OCT family protein sequences were extracted and submitted to Expasy (https://web.expasy.org/, accessed on 5 February 2025) to analyze their characteristics.
2.3. Localization of OCT Family Genes on Tomato Chromosomes
According to the annotation file of the tomato genome, details of all tomato chromosomes were obtained. The relative position of SlOCT genes on chromosomes was visualized using TBtools software.
2.4. Gene Structure, Protein Conserved Motifs and Domains
Based on the annotation file of the tomato genome, the information on whether the SlOCT genes contain introns, the length and number of introns were obtained. For conserved motifs, the MEME database was selected for identification (accessed on 11 February 2025), protein sequences were submitted to the database and the number of motifs was set to the default. Conservative structure domain identification of choice NCBI databases (accessed on 11 February 2025), TBtools software was used to visualize the results together with conserved motifs and gene structures.
2.5. Phylogenetic Tree Construction of OCT Family Proteins in Tomato and Arabidopsis
All currently known sequences of OCT proteins in Arabidopsis were obtained from the TAIR database [4,10,11]. The sequences of the above proteins were input into the phylogenetic tree construction plug-in of TBtools software, and the Clustal W method was selected to align the sequences (clustalw, version 2.0.10, software parameters were set to default). The maximum likelihood method was selected for tree construction, and the boot-strap value was set to 10,000. After the program, the data of the phylogenetic tree were submitted to the ITOL website for graphical drawing (accessed on 5 March 2025).
2.6. Collinearity Between OCT Family Genes
Genomic data for Arabidopsis (TAIR10), rice (Oryza sativa v7.0) and potato (Solanum tuberosum v3.0) were obtained from the TAIR and Phytozome databases. The genome assembly and annotation files were analyzed using TBtools software to investigate the collinearity of SlOCT genes. The genomes and corresponding annotation files of different plants were submitted to the One Step MCScanX-Super Fast plugin of TBtools software to analyze the synteny between the OCT genes of different plants (tomato vs. Arabidopsis/rice/potato).
2.7. Identification and Classification of cis-Acting Elements in Promoter
The promoter regions (2000 bp upstream) of SlOCTs were screened for cis-acting elements using the PlantCARE database (accessed 4 March 2025). The resulting data were graphed using the heatmap plug-in in the TBtools software.
2.8. Expression of SlOCT Family Genes Under Cold and Salt Stresses Based on Publicly Available Transcriptome Data
The transcriptome data used in this study were all open-access data obtained from the NCBI GEO database (https://www.ncbi.nlm.nih.gov/gds/, accessed on 4 March 2025). The data download numbers are GSE148887 (cold stress; cultivated tomato Micro-TOM) and PRJNA888477 (salt stress; cultivated tomato M82). FPKM values of SlOCT genes were extracted, and TBtools software was used to draw visual heat maps.
2.9. qRT-PCR Experiments
Total RNA was extracted from tomato tissues using the Trizol method. Tomato tissues at 0 h, 0.5 h, 2 h, 4 h, 6 h, 12 h, 24 h and 48 h of stresses as described in 2.1 were used in qRT-PCR experiments to define the expression of SlOCT family genes under cold and salt stresses; tomato roots 21 days after transfection with the SlOCT20-silencing vector (the recombinant vector pTRV2-SlOCT20 mentioned in 2.11) were used to detect SlOCT20 silencing efficiency. To analyze the tissue-specific expression of the SlOCT20 gene, samples of roots, stems, leaves, flowers and fruits were collected from 90-day-old tomato seedlings. cDNA was synthesized from total RNA using the HiScript III RT SuperMix for qPCR (+gDNA wiper) kit (Vazyme Biotech Co., Nanjing, China). All qRT-PCR primers were designed using the NCBI website online tool, and all primers used in this study are listed in Table S1. The qRT-PCR experiments were performed using the ChamQ Universal SYBR qPCR Master Mix kit (Vazyme Co., Nanjing, China); the reaction temperature and time referred to the instructions of the kit. Three replicates were performed for each sample. After the qRT-PCR reaction, the specificity of the primers was verified based on the presence or absence of a single, sharp peak in the melting curve. β-actin in tomato was chosen as the reference gene [12,13], and the relative expression was calculated using the 2^−∆∆Ct^ method [14].
2.10. Analysis of Subcellular Localization
PCR primers were designed according to the CDS sequence (does not contain a stop codon) of the SlOCT20 gene and the sequence of the pCAMBIA2300-EGFP vector [15]. The restriction sites were KPN I and BamH I, and the EGFP protein was fused to the C-terminus of SlOCT20; primers were listed in Table S1. The CDS of SlOCT20 with the addition of a homology arm was amplified using tomato cDNA as a template and ligated to the linearized pCAMBIA2300-EGFP vector using the homologous recombinase product from Vazyme Biotech Co., Ltd. (Nanjing, China). The recombinant vector was transferred into Agrobacterium tumefaciens after Sanger sequencing verification by Sangon BioEngineering Co., Ltd. (Shanghai, China). Agrobacterium tumefaciens carrying the vector was injected into Nicotiana benthamiana, and fluorescence signals in transiently transformed Nicotiana benthamiana leaf cells were observed using laser confocal microscopy (1–1000×, Nikon Co., Tokyo, Japan). EGFP was excited using a 488 nm luminescence wavelength.
2.11. Silencing of the Tomato SlOCT20 Gene
VIGS targets and primers were designed according to the sequences of the SlOCT20 gene and vector (Table S1), and PCR was performed. The VIGS target was located in the CDS region of the SlOCT20 gene (1–300 bp), with a total length of 300 bp. The uniqueness of the target was verified by the BLAST target sequence in the whole tomato genome sequence. PCR amplification system: cDNA from Ailsa Craig 1 μL, upstream and downstream primers 1 μL each, KOD One^TM^ PCR Master Mix (TOYOBO Co., Ltd., Osaka, Japan) 25 μL, ddH_2_O 22 μL. Thirty cycles of 98 °C for 10 s, 56 °C for 5 s and 68 °C for 10 s were performed. The products were subjected to 1% agarose gel electrophoresis to purify the correct bands. The target sequences were ligated into the pNC-pTRV2 empty vector using the Nimble Cloning commercial kit [16]. pTRV2-SlOCT20, empty vector pTRV2-00 and pTRV1, which were verified by Sanger sequencing, were transformed into Agrobacterium tumefaciens. Transient transformation was performed using the same method as Chen et al.; bacteria were resuspended using MgCl_2_-IM liquid medium, OD600 was adjusted to 0.6 before injection into tomato seedlings and the seedling age of the tomatoes used was 3 weeks [17]. After 21 days of transformation, the efficiency of SlOCT20 silencing was identified by qRT-PCR experiments.
2.12. Analysis of Physiology
H_2_O_2_ content was measured using the commercial kit H_2_O_2_/2/Y (Keming Co., Suzhou, China). The detection was carried out according to the instructions. The rate of O_2_^−.^ production was detected according to the method proposed by Elstner and Heupel [18]. The PSII maximum photochemical quantum yield (Fv/Fm) was detected using a Plant fluorescence imager (WALZ company, Effeltrich, Germany). The intact tomato leaves were removed and placed on the test table, and the Fv/Fm value was directly detected using the built-in software of the Plant fluorescence imager after switching on.
SOD, POD, CAT activities and MDA content in tomato were detected by kits (SOD/1/Y, POD/1/Y, CAT/1/Y and MDA/1/Y; Keming Co., Suzhou, China). Cadaverine content was detected by Plant Cadaverine Elisa Kit (BLL105866E; Shanghai Baililai Biotechnology Co., Shanghai, China). Three biological replicates were performed for each sample. Individual tomato plants were sampled for each biological replicate. Three parallel technical replicates were performed for each sample.
2.13. Statistical Analyses
The experimental data generated in this study were tested for significance; the data of a single independent variable were tested for significance by one-way ANOVA, and the data of two independent variables were tested for significance by two-way ANOVA using IBM SPSS software (Version 22). The means were separated using Fisher’s protected LSD test at the 5% level of probability in two-way ANOVA, and the t-test was used in a one-way ANOVA.
3. Results
3.1. Identification of OCT Family Genes in Tomato
The tomato genome and its annotation files were retrieved from the Phytozome database, while Arabidopsis OCT protein sequences were obtained from the TAIR database. The coding sequences (CDS) of tomato were extracted from the genome file and converted into corresponding protein sequences. These sequences were then subjected to a BLAST search against the Arabidopsis OCT protein sequences. Additionally, HMM files for the OCT proteins were sourced from the Pfam database, and these files were used for HMM-based identification within the tomato protein sequences. By integrating the results from both the BLAST and HMM analyses, a total of 52 OCT genes were successfully identified in tomato (Table S2).
3.2. Chromosomal Localization of Tomato OCT Genes and Characters of Encoded Protein
The tomato OCT genes were assigned names from SlOCT1 to SlOCT52 based on their positions on the chromosomes, with their chromosomal distribution visualized using TBtools software (Figure 1). Analysis revealed that the SlOCT genes are dispersed across all tomato chromosomes, with chromosome 2 containing the most SlOCT genes (ten genes) and chromosomes 5 and 11 containing the fewest SlOCT genes (only one gene). Most of the SlOCT genes were distributed at both ends of the chromosome. Regarding the encoded proteins, the amino acid lengths varied from 251 (SlOCT16) to 738 (SlOCT27), molecular weights ranged from 27.60 kDa (SlOCT16) to 79.33 kDa (SlOCT27), and the isoelectric points varied between 4.60 (SlOCT45) and 10.08 (SlOCT19) (Table S2).
3.3. Structure of Tomato OCT Family Genes and Conserved Motifs and Domains of Their Encoded Proteins
The tomato OCT family protein sequences were submitted to the MEME and NCBI databases to identify conserved motifs and domains, respectively. These results, along with the tomato genome annotation file, were processed using TBtools software for structural analysis and visualization of the tomato OCT genes. A total of ten conserved motifs (Figure 2A) and seven conserved domains were identified in the tomato OCT family members. A total of 21 SlOCT proteins contained the MFS superfamily domain, 12 SlOCT proteins contained the MFS_-STP-like domain, 9 SlOCT proteins contained the MFS-GLUT6-8-Class3-like domain, 4 SlOCT proteins contained the MFS-GLUT-like domain, 2 SlOCT proteins contained the MFS-GLUT10-12-Class3-like domain, 3 SlOCT proteins contained the MFS-HMIT-like domain and only 1 SlOCT protein contained the MFS-SLC46-TetA-_like domain (Figure 2B). In addition, all SlOCT genes contained introns (Figure 2C).
3.4. Phylogenetic Analysis of OCT Family Proteins
To reconstruct the phylogenetic relationship, OCT protein sequences from both Arabidopsis (AtOCT1-AtOCT6, TAIR10) and tomato were aligned and used for constructing the phylogenetic tree. The proteins in the phylogenetic tree were classified into four distinct branches. Group I contains two tomato OCT proteins, and the homology of these two proteins is low. Group II contains 5 tomato OCT proteins, Group III contains 10 tomato OCT proteins and Group IV contains 35 tomato OCT proteins and six Arabidopsis OCT proteins (Figure 3), suggesting that some Arabidopsis and tomato OCT proteins may have come from the same ancestor.
3.5. Collinearity Analysis of Tomato OCT Family Genes
By analyzing the genome-wide collinearity of the 52 identified SlOCT genes, duplication events within the OCT family were investigated. The results revealed nine pairs of collinear tomato OCT genes, such as SlOCT30 and SlOCT44 (Figure 4A), suggesting that these genes underwent repeated duplication events during evolution. Additionally, synteny analysis was performed to explore the evolutionary relationships between tomato OCT genes and those from other plants. The analysis identified 32 syntenic relationships between tomato OCT genes and Arabidopsis OCT genes (Figure 4B), 54 syntenic relationships with potato OCT genes (Figure 4C), and 11 syntenic relationships with rice OCT genes (Figure 4D). Notably, the highest number of syntenic relationships was observed between tomato and potato OCT genes, likely due to the close evolutionary relationship between these two solanaceous species.
3.6. Analysis of cis-Acting Elements in the Promoters of Tomato OCT Family Genes
To investigate the types, quantities and functions of cis-acting elements in the promoter regions of tomato OCT family genes, the sequences of the promoter regions of all OCT genes were extracted for identification. The identification results were statistically analyzed, and a visual heatmap was generated using TBtools software. In the promoter regions of tomato OCT genes, 68 cis-acting elements were identified and predominantly assigned to four functional groups: light response, plant development, hormone response, and stress response. Light response and plant development elements, such as Box 4 and CCGTCC motifs, were commonly found in plant gene promoters and were also prevalent in the promoters of tomato OCT genes. Several hormone response elements, including ABRE and AuxRE, were detected, suggesting that tomato OCT genes may be regulated by certain hormones. Importantly, the promoters of tomato OCT genes were particularly rich in cis-acting elements associated with stress response, such as MYB and MYC transcription factor binding sites and stress response motifs like the WUN-motif (Figure 5).
3.7. Transcriptome Data and qRT-PCR Analysis of Tomato OCT Family Genes Under Salt and Cold Stress
To gain an initial understanding of the expression patterns of OCT family genes in tomato under various stresses, we retrieved publicly available transcriptome data from the NCBI database, focusing on representative environmental stresses (salt and cold stress). The FPKM values for the OCT family genes were extracted, and a heatmap was generated using TBtools software. Except for SlOCT34 and SlOCT45, all other tomato OCT family genes had detected FPKM values in the tomato transcriptome data under salt stress (Figure 6A). Most of the tomato OCT family genes showed changes in expression under salt stress, indicating that OCT family genes could respond to salt stress. Under cold stress, FPKM values of SlOCT1/2/15/28/34/42/45 were not detected in transcriptome data, and some of the other SlOCT family genes whose FPKM values were detected also showed changes in gene expression (Figure 6B), implying functional diversity of SlOCT family genes.
To further understand the expression patterns of tomato OCT family genes under different stresses and to explore the potential key genes that function under salt or cold stress, we selected genes with stable FPKM values and relatively regular expression in the salt and cold stress transcriptome data for stress qRT-PCR analysis (SlOCT5/6/7/20/22/23/36/39/44/51). The results showed that salt stress up-regulated the expression of SlOCT7, SlOCT20 and SlOCT39 and down-regulated the expression of SlOCT5, SlOCT6 and SlOCT22. The expression of SlOCT23 was first up-regulated and then down-regulated, and the expression of SlOCT44 was first down-regulated and then up-regulated. There was no significant change in the expression of SlOCT36 and SlOCT51 (Figure 7A). Under cold stress, SlOCT7, SlOCT20, SlOCT23 and SlOCT44 were up-regulated, SlOCT5, SlOCT22, SlOCT36 and SlOCT51 were down-regulated, and SlOCT6 and SlOCT39 expressions did not change significantly (Figure 7B). Notably, the expression of SlOCT20 was significantly up-regulated under both salt and cold stress, and the up-regulation was the highest. These findings led to the hypothesis that SlOCT20 is involved in salt and cold stress responses in tomato, leading to its selection as a prime candidate for functional validation.
3.8. Analysis of SlOCT20 Protein Subcellular Localization
The SlOCT20 was ligated to the vector pCAMBIA2300-EGFP, and the EGFP tag was fused to the C-terminus of the SlOCT20 protein. The validated vector pCAMBIA2300-EGFP-SlOCT20 and the empty vector pCAMBIA2300-EGFP were transiently transformed into epidermal cells of Nicotiana benthamiana leaves, respectively. Laser confocal microscopy was used to visualize fluorescence. The green fluorescence signal of EGFP and chloroplast autofluorescence in the dark field could be clearly observed in the transformed empty vector group, indicating that the transient transformation was successful. The green fluorescence signal of EGFP in the transformed recombinant vector pCAMBIA2300-EGFP-SlOCT20 group mainly appeared in the cell membrane, indicating that the SlOCT20 protein was mainly localized in the cell membrane (Figure 8).
3.9. Silencing of SlOCT20 Reduced the Cold Tolerance of Tomato
VIGS was used to silence SlOCT20 in tomato to understand the function of tomato SlOCT20 under salt and cold stresses. The silencing efficiency of SlOCT20 was identified 21 days after transient transformation. The expression of SlOCT20 in most (8/10) SlOCT20-silenced tomatoes was significantly lower than that in the control (Figure S1), indicating that SlOCT20 has been silenced in tomatoes. Tomatoes with higher silencing efficiency were chosen for subsequent experiments (pTRV2-SlOCT20#4, 7, 8, 9).
Phenotypic differences between control and SlOCT20-silenced tomatoes were analyzed after 48 h of salt and cold stresses. After 48 h of salt and cold stresses, both control and SlOCT20-silenced tomatoes showed a wilting phenotype, but there was no significant difference in phenotype between the control and SlOCT20-silenced tomatoes after salt stress. However, compared with the control, SlOCT20-silenced tomatoes exhibited a less cold-tolerant phenotype after cold stress (Figure 9A). Reactive oxygen species accumulation (H_2_O_2_ and O_2_^−.^) and PSII maximum photochemical quantum yield (Fv/Fm) are often used to evaluate the degree of plant damage under cold stress. H_2_O_2_ content, rate of O_2_^−.^ production, and Fv/Fm were measured in control and SlOCT20-silenced tomato leaves after cold stress. H_2_O_2_ content and rate of O_2_^−.^ production in SlOCT20-silenced tomatoes were higher than that in the control (Figure 9B), and the Fv/Fm value was lower than that in the control (Figure 9C). This suggests that SlOCT20-silenced tomatoes were more severely damaged and less tolerant to cold after cold stress.
3.10. Tomato SlOCT20 May Positively Regulate Cold Tolerance by Regulating Cadaverine Transport from Soil to Roots
As mentioned above, OCT family proteins can transport some organic cations, such as carnitine and cadaverine, and cadaverine has been reported to enhance cold tolerance by mitigating oxidative damage in plants. We examined the expression of the SlOCT20 gene in different tomato organs, and although the SlOCT20 gene is expressed in different tomato organs, the highest expression level is found in the root system (Figure 10A). After cold stress, the expression of SlOCT20 was upregulated in tomato roots, and the silencing of SlOCT20 had a negative impact on this response pattern (Figure S2). The results of cadaverine detection in the roots of control and SlOCT20-silenced tomatoes showed that cadaverine content in the roots of SlOCT20-silenced tomatoes was significantly lower than that of control tomatoes under cold stress (Figure 10B). SOD, POD, CAT activities and MDA content in the roots of control and SlOCT20-silenced tomatoes were increased under cold stress. In addition, SOD, POD, CAT activities and MDA content in SlOCT20-silenced tomatoes were significantly higher than those in control tomatoes (Figure 10C), indicating that the oxidative damage of SlOCT20-silenced tomatoes was more serious under cold stress. In conclusion, tomato SlOCT20 may enhance cold tolerance by regulating cadaverine accumulation in roots and reducing oxidative damage.
4. Discussion
Tomatoes often suffer from various environmental stresses during their growth and development, such as cold stress and saline-alkali stress [19]. Tomatoes prefer warmth, and cold stress is extremely harmful to their growth and development. The regulation mechanism of plants to cold stress is mainly through Ca^2+^, hormones and other signaling pathways to activate the expression of cold resistance genes, such as CBFs and ICEs. In addition, other indirect mechanisms also regulate plant tolerance to cold stress [20]. The transmembrane transport of cations not only plays a role in mineral absorption, osmotic pressure regulation, cell expansion, and signal transduction, but also relates to plant stress tolerance [3]. In this study, all of the OCT family genes were identified in the tomato genome version 4.0 (Table S2), and the structures of these OCT genes and encoded proteins were characterized (Figure 2). Most of the SlOCT genes are unevenly distributed at both ends of the chromosome (Figure 1). This phenomenon is frequently seen in other similar studies [21,22,23,24,25] and may be related to gene evolutionary expansion. Phylogenetic and collinearity analysis of OCT proteins and OCT family genes in tomato and different plants showed that OCT genes in tomato and OCT genes in other plants (such as Arabidopsis, rice and potato) may come from the same ancestor, and SlOCT family genes were repeatedly generated during evolution (Figure 3 and Figure 4). In addition, the identification of cis-acting elements in the promoter of tomato OCT family genes also implies a correlation between gene function and environmental stress in this family.
Under stress, plants will adjust their gene expression to adapt to stress and reduce stress damage. We found that many tomato OCT family genes could respond to salt and cold stress using publicly available omics data and qRT-PCR validation under salt and cold stresses (Figure 6 and Figure 7). We selected the SlOCT20 gene, whose expression was significantly up-regulated and with the highest up-regulation fold under salt stress and cold stress, as the hypothetical key gene for later functional studies. We cloned the full-length CDS of SlOCT20 in tomato and found that the encoded protein was mainly localized to the cell membrane (Figure 8). The localization of tomato OCT protein obtained in this study is consistent with the localization of Arabidopsis OCT protein [5]. The expression of SlOCT20 protein on the plasma membrane suggests that it may have an ion transport function. We treated the constructed SlOCT20-silenced tomatoes with salt and cold stress, and the phenotype of tomato plants treated with stress for 48 h showed that under cold stress, SlOCT20-silenced tomatoes exhibited a phenotype that was less cold-tolerant compared to the control tomatoes (Figure 9A). We also tried to demonstrate the reduction of cold tolerance in SlOCT20-silencing tomatoes from other physiological indicators. It has been shown that plants will produce a strong burst of reactive oxygen species in cells after stress [26], and excessive bursts of reactive oxygen species in a short period of time can impair cell function [27]. Therefore, tissue-reactive oxygen species content is often used to assess the severity of tissue damage caused by stress. The results of H_2_O_2_ content and the rate of O_2_^−.^ production showed that the above indicators in SlOCT20-silenced tomatoes were higher than those in the control (Figure 9B), indicating that the oxidative stress of SlOCT20-silenced tomatoes was more serious after cold stress. In addition, the Fv/Fm can reflect photosynthetic capacity and is often used to assess plant health under cold stress. Lower values of Fv/Fm indicate weaker photosynthetic capacity and poorer health of the plant [28]. The Fv/Fm of SlOCT20-silenced tomato plants under cold stress was significantly lower than that of the control plants (Figure 9C). The PSⅡ maximum photochemical quantum yield also showed that the silencing of the SlOCT20 gene reduced the cold tolerance of tomato plants.
We tried to understand the underlying mechanism of SlOCT20 in regulating tomato cold tolerance. By detecting the expression levels of SlOCT20 in different organs of tomato, we found that SlOCT20 was most highly expressed in tomato roots (Figure 10A). After cold stress, the expression of SlOCT20 was upregulated in tomato roots, and the silencing of SlOCT20 had a negative impact on this response pattern (Figure S2). Combined with the results of localization of SlOCT20 protein on the cell membrane, we hypothesized that the tomato SlOCT20 protein may play a role in the uptake of some beneficial organic cations in soil by the root system. Cadaverine, as an organic cation widely present in soil, plays an active role in plant stress resistance and antioxidant processes [7,8]. Our results showed that cadaverine content in the roots of SlOCT20-silenced tomato was significantly lower than that of the control under cold treatment (Figure 10B). In addition, silencing of SlOCT20 significantly increased SOD, POD, CAT activities and MDA content in tomato roots under cold stress (Figure 10C). The complete metabolic pathway formed by the above-mentioned antioxidant enzymes in plants can convert reactive oxygen species into biologically harmless substances. Their activities increase rapidly in the early stage of stress to remove harmful substances, and the more severe the oxidative damage to the cells, the more intense the burst of enzyme activity [29,30,31]. Previous studies have shown that cadaverine absorption, transport, and accumulation in plants are positively correlated with antioxidant systems under stress [32]; therefore, we speculate that silencing SlOCT20 may lead to a reduction in cadaverine accumulation in roots, thereby reducing the antioxidant capacity of tomato under cold stress.
5. Conclusions
In this study, 52 OCT family genes were identified in tomato for the first time, and a detailed bioinformatics analysis of these genes was conducted. A key gene regulating cold stress tolerance, SlOCT20, was identified. SlOCT20 was mainly localized in the cell membrane and positively regulates cold tolerance in tomato.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Igor P. Sergey S. Polyamines control of cation transport across plant membranes: Implications for ion homeostasis and abiotic stress signaling Front. Plant Sci.2014515410.3389/fpls.2014.0015424795739 PMC 4006063 · doi ↗ · pubmed ↗
- 2Zhou S. Zeng S. Shu Y. Drug-Drug interactions at organic cation transporter 1Front. Pharmacol.20211262870510.3389/fphar.2021.62870533679412 PMC 7925875 · doi ↗ · pubmed ↗
- 3Torres G.A. Lelandais-Briere C. Besin E. Jubier M.F. Roche O. Mazubert C. Corre-Menguy F. Hartmann C. Characterization of the expression of Phaseolus vulgaris OCT 1, a dehydration-regulated gene that encodes a new type of phloem transporter Plant Mol. Biol.20035134134910.1023/A:102201422989912602865 · doi ↗ · pubmed ↗
- 4Lelandais-Briere C. Jovanovic M. Torres G.A. Perrin Y. Lemoine R. Corre-Menguy F. Hartmann C. Disruption of At OCT 1, an organic cation transporter gene, affects root development and carnitine-related responses in Arabidopsis Plant J.20075115416410.1111/j.1365-313X.2007.03131.x 17521409 · doi ↗ · pubmed ↗
- 5Strohm A.K. Vaughn L.M. Masson P.H. Natural variation in the expression of ORGANIC CATION TRANSPORTER 1 affects root length responses to cadaverine in Arabidopsis J. Exp. Bot.20156685386210.1093/jxb/eru 44425403917 PMC 4321547 · doi ↗ · pubmed ↗
- 6Hulya T. Serkan E. Rahmi D. Carnitine-induced physio-biochemical and molecular alterations in maize seedlings in response to cold stress Arch. Agron. Soil Sci.20206692594110.1080/03650340.2019.1647336 · doi ↗
- 7Jancewicz A.L. Gibbs N.M. Masson P.H. Cadaverine’s functional role in plant development and environmental response Front. Plant Sci.2016787010.3389/fpls.2016.0087027446107 PMC 4914950 · doi ↗ · pubmed ↗
- 8Paul E.A. The nature and dynamics of soil organic matter: Plant inputs, microbial transformations, and organic matter stabilization Soil Biol. Biochem.20169810912610.1016/j.soilbio.2016.04.001 · doi ↗