Porcine FRZB (sFRP3) Negatively Regulates Myogenesis via the Wnt Signaling Pathway
Jingru Nie, Yu Fu, Xin Hao, Dawei Yan, Bo Zhang, Hao Zhang

TL;DR
This study shows that the FRZB gene limits muscle growth in pigs by blocking Wnt signals, which could help improve meat production through selective breeding.
Contribution
The study identifies FRZB as a negative regulator of myogenesis in pigs via the Wnt pathway, linking it to breed-specific muscle growth differences.
Findings
FRZB expression is higher in slow-growing indigenous pigs compared to fast-growing commercial breeds.
Reducing FRZB in muscle cells increases proliferation, migration, and formation of larger muscle fibers.
FRZB silencing upregulates myogenic and fusion markers while promoting hypertrophy-related genes.
Abstract
Skeletal muscle growth is the primary factor determining meat production in the pig industry. A gene called FRZB is known to affect bone development, but its specific role in muscle formation is not well understood. This study aimed to investigate how FRZB influences muscle growth and whether it contributes to the size differences between pig breeds. We compared slow-growing indigenous pigs with fast-growing commercial pigs and found that the slow-growing breeds had significantly higher levels of FRZB in their loin muscles during development. To understand its function, we reduced the levels of FRZB in muscle cells in the laboratory. We observed that cells with lower FRZB levels multiplied faster and formed larger, thicker muscle fibers. Our results suggest that FRZB acts like a natural “brake” on muscle growth by blocking specific growth signals. This research is valuable to society…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
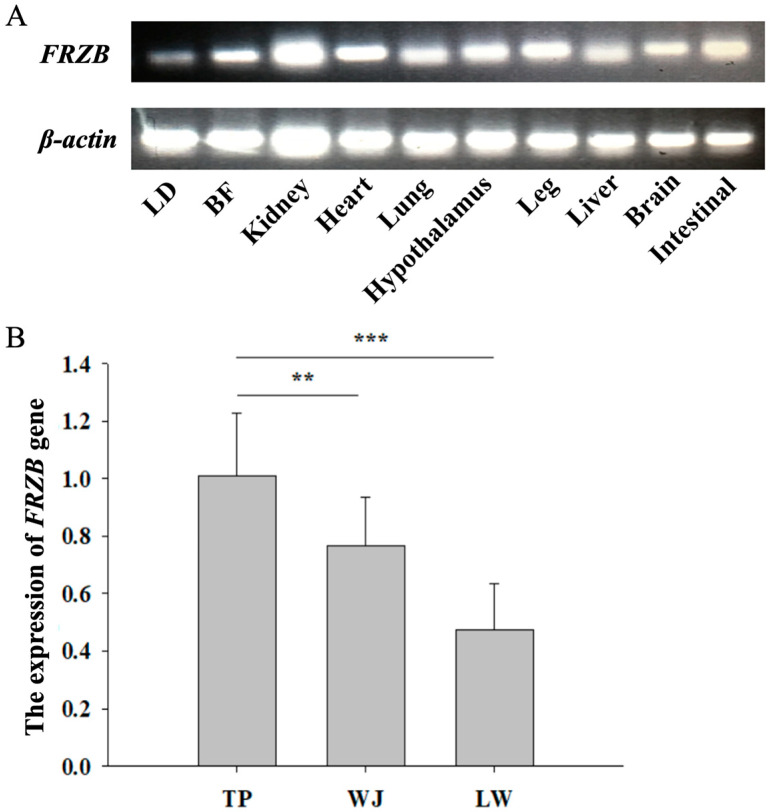 Figure 1
Figure 1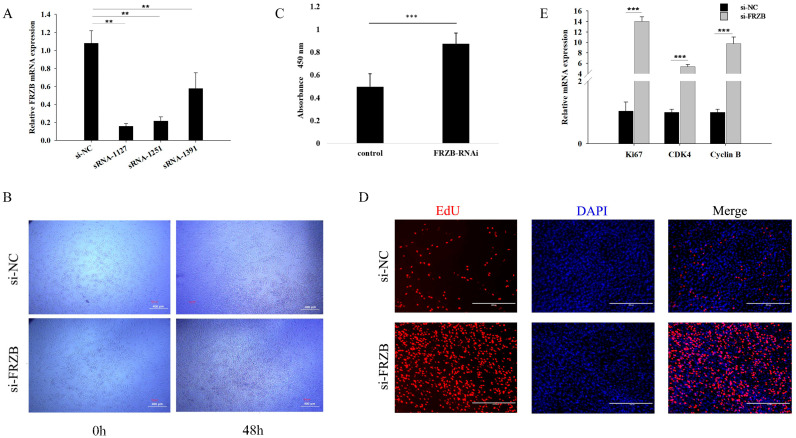 Figure 2
Figure 2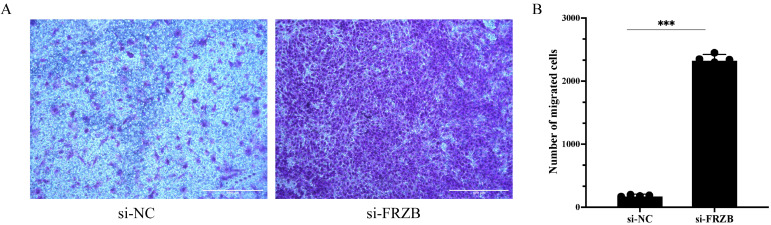 Figure 3
Figure 3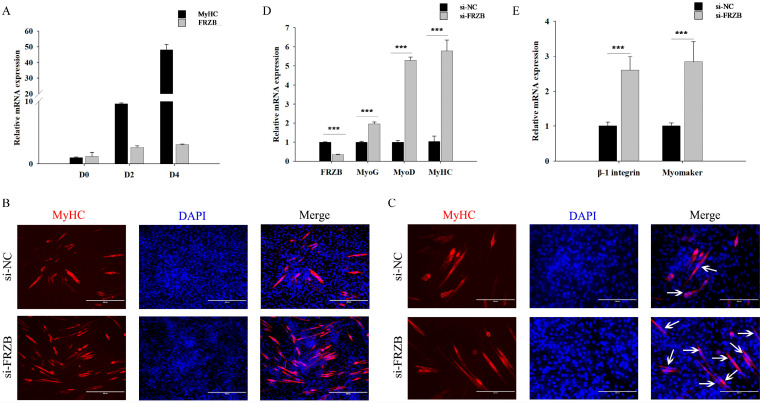 Figure 4
Figure 4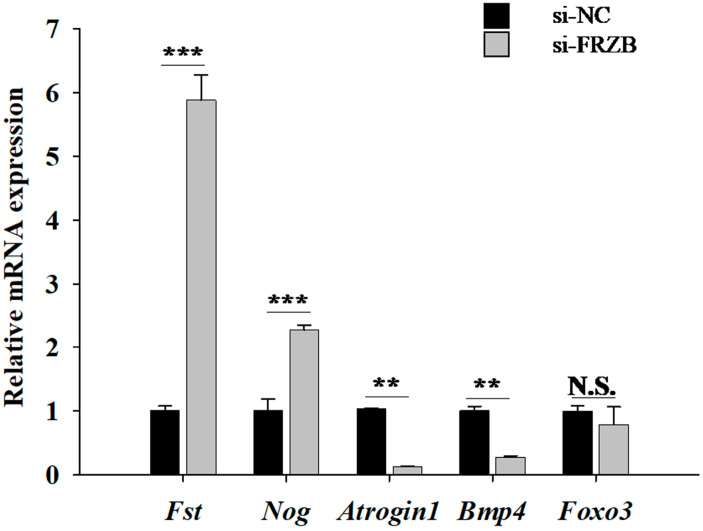 Figure 5
Figure 5- —Yunnan Major Science and Technology Project
- —Xizang Major Science and Technology Project
- —National Natural Science Foundation of China
- —Chinese Universities Scientific Fund
- —the 2115 Talent Development Program of China Agricultural University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsWnt/β-catenin signaling in development and cancer · Muscle Physiology and Disorders · Reproductive Biology and Fertility
1. Introduction
Pork is the most essential meat source in China, and modern swine breeding increasingly prioritizes the balance between growth performance and superior meat quality. Since skeletal muscle growth and fiber characteristics are primary factors determining meat production and quality, identifying key genetic regulators of myogenesis is essential for improving carcass traits across diverse pig breeds [1]. During vertebrate embryogenesis, skeletal muscles in the limbs arise from muscle progenitor cells derived from somites [2]. Postnatal muscle growth primarily depends on increases in muscle fiber volume. Muscle development involves a series of biological processes, including myoblast proliferation, differentiation, and fusion, which lead to the formation of multinucleated myotubes that eventually mature into functional muscle fibers [3]. The growth of these fibers is tightly regulated by specific muscle-related transcription factors [4] and protein kinases [5]. Among them, myogenic regulatory factors (MRFs) serve as central nodes within the transcriptional network and directly drive skeletal muscle development [6]. Cyclin-dependent kinases further regulate the myoblast cell cycle, balancing proliferation with differentiation [7,8]. The temporal and spatial expression of these regulators is strictly controlled to ensure the orderly progression of myogenesis [1,9].
Our previous study revealed that the expression level of the frizzled-related protein (FRZB) gene in the longissimus dorsi (LD) muscle of slow-growing Chinese indigenous pig breeds (Diannan small-eared pig and Tibetan pig, DSP-TP) was approximately 3.8-fold higher than that observed in fast-growing introduced breeds (Landrace and Yorkshire, LL-YY) [10]. FRZB encodes secreted frizzled-related protein 3 (sFRP3), which is a member of the sFRP family involved in the Wnt signaling pathway. sFRP3 contains a cysteine-rich domain (CRD) homologous to the Wnt-binding site of frizzled receptors, enabling it to competitively disrupt Wnt-frizzled interactions [11]. Members of the sFRP family are generally recognized as Wnt signaling inhibitors [12]. In China, the physiological dichotomy of the swine industry is best exemplified by the contrast between indigenous breeds and commercial breeds. The TP, native to high altitudes, is an evolutionary marvel adapted to extreme hypoxia, exhibiting slow growth but exceptional pork flavor. The Wujin pig (WJ), a classic “fat-type” breed, is celebrated for its superior intramuscular fat deposition and its role in premium ham production. In contrast, the Large White (LW) pig represents the global standard for rapid muscle hypertrophy and feed efficiency [13,14]. Investigating the molecular basis of these divergent growth phenotypes, particularly the role of FRZB, provides a roadmap for balancing production speed with meat quality.
Canonical Wnt/β-catenin signaling promotes the expansion of myogenic progenitors and drives the expression of MRFs during both fetal development and regeneration [15,16,17]. In contrast, increasing the levels of extracellular Wnt antagonists can impede myogenesis. For example, transplacental delivery or misexpression of sFRP3/FRZB in embryos reduces myofiber formation in a dose-dependent manner and blocks somitic myogenesis upstream of myogenic differentiation antigen (MyoD) [18,19]. These findings suggest that FRZB functions as a potential brake in myogenesis. Beyond the muscle, canonical Wnt signaling inhibits adipogenic differentiation, whereas Wnt antagonism promotes adipogenesis [20,21,22]. Therefore, tissue-specific FRZB expression levels may influence the balance between muscle growth and fat deposition in pigs.
Recent studies have further highlighted the importance of Wnt signaling in muscle biology. For example, Wnt/β-catenin signaling has been identified as a key mediator in muscle-bone crosstalk, influencing both tissue regeneration and homeostasis [23]. A comprehensive review also highlighted new insights into Wnt signaling pathways in development and disease, emphasizing their interplay with Notch, TGF-β, and Hippo pathways [24]. However, direct functional studies on FRZB in porcine muscle development remain limited. In the pig genome, FRZB is located on chromosome 15 (Sscrofa11.1: ~88.33–88.38 Mb) and is composed of multiple exons. Natural variations in its regulatory regions are associated with growth-related traits in pigs [25].
To bridge this knowledge gap, this study aimed to characterize the expression profile of FRZB in fetal tissues of diverse pig breeds and to elucidate its regulatory function in myoblast proliferation and differentiation using a C2C12 model. Given the established role of canonical Wnt signaling in promoting myogenesis, we hypothesized that the differential expression of FRZB might act as a critical molecular switch modulating myoblast behavior, thereby contributing to the phenotypic variations in muscle growth observed between pig breeds.
2. Materials and Methods
2.1. Experimental Materials
All animal procedures followed the National Research Council Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee (approval number: AW80203202-1-1). Tibetan (TP) and Wujin (WJ) pigs, two indigenous Chinese breeds with relatively slow growth rates, and Large White (LW) pigs, a fast-growing commercial breed, were reared at the Tibet Agriculture and Animal Husbandry College. The pregnant sows were raised in semi-closed cement floor pigsties under standardized environmental conditions. The sows were fed twice daily (09:00 and 16:00) with a quantitative allowance based on their gestational stage, while water was provided ad libitum. All animals were immunized and dewormed according to standard farm protocols. The dietary formula was designed based on the Chinese pig feeding standard (NY/T 65-2004) [26] for gestating sows. The detailed formula composition and nutritional levels are listed in Supplementary Table S1. The pregnant sows were slaughtered at 60 days post-insemination, and at least six fetuses (mixed sex) from each breed (TP, WJ, and LW) were collected. Fetuses were selected randomly based on normal morphology and comparable body size. Multiple fetal tissues, namely longissimus dorsi (LD), back fat (BF), kidney, heart, lung, hypothalamus, leg muscle, liver, brain, and intestine (jejunum), were collected for semi-quantitative reverse transcription PCR (sqRT-PCR). Fetal LD tissues (12th rib region) were collected for sqRT-PCR and quantitative real-time PCR (qRT-PCR).
2.2. C2C12 Cell Culture and RNA Interference
C2C12 myoblasts (Type Culture Collection of the Chinese Academy of Sciences, China) were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Watham, MA, USA) supplemented with 10% fetal bovine serum (FBS; Gibco) and 1% penicillin/streptomycin (Gibco). Cell differentiation was induced by culturing the cells in DMEM supplemented with 2% horse serum (Gibco) and 1% penicillin/streptomycin. To suppress FRZB expression, we designed small interfering RNAs (siRNAs) based on the FRZB gene sequence and selected the highest-scoring candidates (siRNA-1127, siRNA-1251, and siRNA-1391) from the design platform, with the numerical identifiers of each siRNA corresponding to its position relative to the FRZB transcriptional start site. The FRZB siRNA sequences were as follows: siRNA-1127, sense: CCGGAACAAUUACAACUAUTT, antisense: AUAGUUGUAAUUGUUCCGGTT; siRNA-1251, sense: GACACCGUCAAUCUUUAUATT, antisense: UAUAAAGAUUGACGGUGUCTT; and siRNA-1391, sense: GGAUCGGCUUGGUAAGAAATT, antisense: UUUCUUACCAAGCCGAUCCTT; si-NC, sense: UUCUCCGAACGUGUCACGUTT, antisense: ACGUGACACGUUCGGAGAATT. Cell transfection was performed using Lipofectamine 2000 (Invitrogen, Waltham, MA, USA) according to the manufacturer’s instructions. For differentiation assays, at 24 h post-transfection, the growth medium was replaced with differentiation medium (DMEM supplemented with 2% horse serum) to induce myogenic differentiation. Based on the knockdown efficiency results (verified by qRT-PCR), siRNA-1127 exhibited the strongest inhibitory effect and was thus selected for all subsequent functional assays.
2.3. RNA Extraction and mRNA Level Measurement
Total RNA was extracted from the cells and tissues using TRIzol reagent (Invitrogen). Briefly, TRIzol was added to the samples for ice-bath lysis or homogenization until tissue or cell debris was no longer visible. RNA was then separated from protein and DNA via phenol-chloroform extraction, precipitated with isopropyl alcohol, and collected via centrifugation (4 °C, 12,000 rpm, 15 min). The concentration and purity of RNA were determined using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), with an A260/280 ratio between 1.8 and 2.0. RNA integrity was verified by 1% agarose gel electrophoresis. The extracted RNA was reverse-transcribed using the FastQuant Reverse Transcriptase Kit (TIANGEN, Beijing, China). The mRNA expression levels of the target genes were analyzed using semi-quantitative PCR and qRT-PCR with specific primers (Supplementary Table S2). Primers were designed using the NCBI Primer-BLAST tool [27] based on the specific gene sequences. Expression levels were normalized using the 2^−ΔΔCt^ method [28], with β-actin serving as the housekeeping reference gene.
2.4. Cell Proliferation Measurement
5-Ethynyl-2-deoxyuridine (EdU; RiboBio, Guangzhou, China) and Cell Counting Kit-8 (CCK-8; Solarbio, Beijing, China) assays were conducted following the manufacturers’ protocols. For the EdU assay, cells transfected for 24 h were incubated with 50 µmol/L EdU for 2 h, fixed with 4% paraformaldehyde (PFA), permeabilized with 0.5% Triton X-100, incubated with Apollo reaction buffer, and counterstained with Hoechst to visualize nuclei. Images were acquired using a microscope (ZEISS, Jena, Germany). For the CCK-8 assay, transfected cells were incubated with 100 µL of 10% CCK-8 solution at 37 °C for 1 h, after which absorbance was measured at 450 nm using a microplate reader (Biotek, Winooski, VT, USA).
2.5. Transwell Migration Assay
Cell migration was measured using transwell inserts, as previously described [29]. At 24 h post-transfection, myoblasts were trypsinized and counted using a hemocytometer. The cells were then resuspended in serum-free DMEM and added to the upper chamber (200 µL/well), while 500 µL of DMEM supplemented with 10% FBS was added to the lower chamber as a chemoattractant. After incubation for 12 h at 37 °C in 5% CO_2_, cells remaining on the upper surface of the membrane were carefully removed. The migrated cells on the underside were stained with crystal violet (30–60 min), imaged in five randomly selected fields per well using a microscope, and quantified using ImageJ software (version 1.53t; National Institutes of Health, Bethesda, MD, USA). Three independent biological replicates were analyzed per group.
2.6. Immunofluorescence (IF)
Myoblasts and muscle tissues were fixed in 4% PFA (20 min) and permeabilized (0.3% Triton X-100 in PBS for 15 min). After blocking (2 h at RT; Beyotime, Haimen, China), the samples were incubated overnight at 4 °C with primary antibodies against the following: MyHC (M4276, 1:200; Sigma-Aldrich, St. Louis, MO, USA), eMyHC (F1.652, 1:50; DSHB, Iowa City, IA, USA), and laminin (L9393, 1:1000; Sigma-Aldrich). Fluorescent secondaries (SA00013-2/-3, 1:400; Proteintech, Rosemont, IL, USA) were added for 1 h at RT in the dark, after which nuclei were counterstained with DAPI (5 min). Images were acquired using a Leica Q500MC system (Leica, Cambridge, UK).
2.7. Statistical Analysis
All quantitative data are presented as the mean ± SD. Prior to statistical analysis, the normality of the data distribution was verified using the Shapiro-Wilk test, and the homogeneity of variance was assessed using Levene’s test. Data conforming to these assumptions were analyzed. Between-group comparisons were performed using a two-tailed Student’s t-test, while multigroup comparisons (e.g., pig breed expression data) were performed using one-way ANOVA followed by Tukey’s post-hoc test. A minimum of three independent biological replicates were included unless otherwise indicated. Statistical analyses were performed using SPSS (version 22.0; IBM, Armonk, NY, USA). The significance thresholds were set at * p < 0.05, ** p < 0.01, and *** p < 0.001.
3. Results
3.1. Tissue Distribution and Breed-Specific Expression of FRZB in Pigs
FRZB expression was broadly detected across multiple fetal tissues of TP, including the LD, BF, kidney, heart, lung, hypothalamus, leg muscle, liver, brain, and intestine (jejunum). Notably, while high expression levels were observed in the kidney and leg muscle, clearly detectable expression was also confirmed in the LD and BF (Figure 1A). Comparative analysis revealed that compared with the fast-growing LW, the slow-growing breeds TP and WJ had significantly higher FRZB expression in LD (Figure 1B). These findings are consistent with our previous transcriptomic analysis, validating the breed-specific upregulation of FRZB in the skeletal muscle of indigenous pigs [10].
3.2. FRZB Knockdown Enhances Proliferation of C2C12 Myoblasts
Skeletal muscle development relies on coordinated processes including myoblast proliferation, migration, differentiation, and myotube fusion. We investigated the role of FRZB via RNA interference in C2C12 myoblasts and confirmed that siRNA-1127 achieved the most efficient knockdown of FRZB expression (Figure 2A). Functional assays demonstrated that FRZB silencing markedly enhanced cell proliferation. Specifically, microscopic observation showed increased cell density (Figure 2B), and the CCK-8 assay revealed significantly higher absorbance values in the si-FRZB group than in the si-NC group at 48 h (Figure 2C). Consistently, the EdU assay showed a significant increase in the percentage of EdU-positive nuclei following FRZB knockdown (Figure 2D). Moreover, qPCR analysis revealed that FRZB depletion significantly upregulated proliferation-associated marker genes including Ki67, cyclin dependent kinase 4 (CDK4), and cyclin B (Figure 2E). Together, these results demonstrated that FRZB knockdown enhances the proliferative capacity of C2C12 myoblasts.
3.3. FRZB Knockdown Markedly Increases Transwell Migration of C2C12 Myoblasts
To assess whether FRZB influences myoblast motility, we performed Transwell migration assays after siRNA-mediated silencing of FRZB in C2C12 cells. Crystal violet staining revealed a visibly denser layer of migrated cells on the lower membrane surface in the si-FRZB group than that in the si-NC group (Figure 3A). Quantification of the average number of migrating cells in five randomly selected microscopic fields per well confirmed a robust and significant increase in the number of migrated cells following FRZB knockdown (*** p < 0.001; Figure 3B). These findings indicate that the loss of FRZB greatly enhances the migratory capacity of C2C12 myoblasts.
3.4. FRZB Knockdown Accelerates Myogenic Differentiation and Augments Myotube Formation in C2C12 Cells
We first monitored the expression profiles of MyHC and FRZB during C2C12 differentiation. Time-course qPCR analysis showed that the expression of MyHC gradually increased, confirming successful myogenic induction. Similarly, FRZB expression also rose throughout the process, reaching its peak on Day 4 (Figure 4A). Subsequent functional assays showed that FRZB knockdown markedly accelerated this myogenic progression. Consistently, immunofluorescence (IF) staining after 4 d in differentiation medium revealed an increased overall abundance and size of MyHC^+^ myotubes in si-FRZB cells (Figure 4B). Higher-magnification imaging further highlighted a significantly higher frequency of multinucleated myotube formation (white arrows, Figure 4C). qPCR on day 4 confirmed significant upregulation of differentiation markers (MyoG, MyoD, and MyHC) (Figure 4D) and fusion-related genes (β1-integrin and Myomaker) (Figure 4E). Together, these data demonstrate that loss of FRZB accelerates myogenic differentiation and enhances myotube growth and fusion.
3.5. FRZB Knockdown Skews C2C12 Transcription Toward a Pro-Hypertrophic Profile
To determine whether FRZB affects growth-promoting or catabolic programs, we profiled canonical hypertrophy- and atrophy-related genes via qPCR on day 4 of differentiation. FRZB knockdown markedly upregulated the hypertrophy-associated genes follistatin (Fst) and noggin (Nog) and significantly downregulated the atrophy-related E3 ligase Atrogin1 (Figure 5). In contrast, Foxo3 showed no significant change (N.S.), and Bmp4 was also downregulated. Taken together, these findings indicate that FRZB depletion shifts the transcriptional landscape toward a pro-hypertrophic and anti-atrophic state, which is consistent with the observed enhancement of myoblast proliferation, differentiation, and myotube formation upon FRZB knockdown.
4. Discussion
In this study, we showed that FRZB is widely expressed across multiple porcine fetal tissues and is relatively elevated in the LD muscle of slow-growing TP/WJ pig breeds compared with the fast-growing LW breed. Although FRZB expression was high in the leg muscle and kidney, our focus on the LD stems from its critical role as the primary indicator of meat quantity and quality in porcine production. In vitro, FRZB silencing in C2C12 myoblasts promoted their proliferation, migration, differentiation, and myotube fusion while shifting transcriptional programs toward hypertrophy and away from atrophy. Collectively, these results support the notion that FRZB acts as a negative regulator of myogenesis.
Mechanistically, FRZB (also known as sFRP3) belongs to the secreted frizzled-related protein family and contains a cysteine-rich domain (CRD) homologous to the Wnt-binding region of frizzled receptors, enabling it to bind to Wnt ligands and block their interaction with cell surface receptors [30]. Such binding can attenuate canonical Wnt/β-catenin signaling, which is critical for myogenic progression. Indeed, previous studies have shown that Wnt signaling promotes muscle regeneration and specification. Polesskaya et al. [31] found that Wnt signals can induce myogenic commitment in satellite cells, analogous to fetal myogenesis. In addition, during head muscle development, the Wnt antagonist Frzb collaborates with BMP inhibitors (e.g., noggin) to regulate myogenesis [32]. Thus, our data showing that FRZB knockdown enhances myogenic outcomes are consistent with the idea that reducing Wnt antagonism permits more robust activation of downstream myogenic gene programs. Moreover, the regulatory role of FRZB in myogenesis likely extends beyond simple Wnt antagonism through complex crosstalk with other signaling networks. For instance, sFRPs can interact with molecules unrelated to the Wnt cascade, such as acting as proteinase inhibitors to interfere with BMP signaling or serving as negative modulators of ADAM10, a key enzyme in Notch pathway activation. In skeletal muscle regeneration, Wnt and Notch signaling often act in opposition to control the balance between progenitor proliferation and terminal differentiation [33,34]. Thus, the observed myogenic effects of FRZB may represent the integrated output of a multi-layered signaling hub.
Beyond muscle regulation, canonical Wnt/β-catenin signaling is known to inhibit adipogenic differentiation by suppressing adipogenic transcription factors (e.g., C/EBPα and PPARγ). In contrast, extracellular Wnt antagonists may shift the cell fate toward adipogenesis under certain conditions [20,21,35]. While we did not directly test adipogenic markers in our experiment, the relatively higher FRZB expression in LD and BF of TP/WJ pigs suggests that elevated FRZB in these breeds partly suppresses myogenesis and favors adipose differentiation, contributing to their slower growth and distinct carcass traits. This hypothesis aligns with the established observation that Wnt antagonism generally promotes adipogenesis [22,35].
Placing our findings within a broader signaling landscape, recent reviews have highlighted the complex crosstalk between Wnt/β-catenin and other pathways (Notch, TGF-β, Hippo, and mechanotransduction) in tissue development and homeostasis [24]. Sharma et al. [36] demonstrated that chromatin-remodeling complexes (mSWI/SNF) regulate the accessibility of Wnt pathway genes during myogenic differentiation, indicating a deeper regulatory layer controlling Wnt responsiveness in muscle cells. Furthermore, recent systematic reviews have emphasized the stage-specific roles of Wnt signaling in muscle-bone crosstalk, suggesting that the precise timing of Wnt antagonism is crucial for balanced tissue growth [23]. These studies underscore that the effects of FRZB might be modulated by broader epigenetic and signaling networks, potentially explaining why a ubiquitously expressed protein can exert tissue-specific effects on muscle mass.
Interestingly, in the context of neuromuscular diseases, Kwan et al. [37] reported FRZB upregulation in limb muscles from patients with amyotrophic lateral sclerosis and mouse models, with expression localized to connective tissue surrounding atrophic fibers. This suggests that FRZB is induced by muscle damage or denervation and may inhibit regeneration via Wnt antagonism. This raises the intriguing possibility that in pathological states involving muscle atrophy or denervation, FRZB upregulation reduces muscle renewal by suppressing Wnt-mediated myogenic signaling. Our finding that FRZB knockdown significantly downregulated the atrophy-related gene Atrogin1 further supports this link between FRZB levels and muscle catabolism.
Despite the findings, our study had some limitations and opens future research directions. First, we acknowledge that the in vivo analysis involved a limited number of sows, which may introduce a nesting effect. However, the consistent molecular trends observed across multiple fetuses provided a rationale for the subsequent mechanistic validation. Second, all functional experiments in this study were conducted in C2C12 cells, which are a classic murine myoblast line. While this model was selected for its high experimental stability and reproducibility compared to primary cells, and despite the high conservation of the FRZB protein sequence and Wnt signaling pathway between pigs and mice, species-specific differences may still exist. Therefore, confirming these inhibitory effects in porcine primary myoblasts or through in vivo experiments remains a necessary step for future research. Third, we inferred a mechanism by which FRZB competitively binds to Wnt ligands and reduces downstream signaling, however, we did not directly measure β-catenin stabilization, TCF/LEF target activation, or receptor-ligand binding dynamics. Incorporation of reporter assays (e.g., TOP/FOP), immunoprecipitation of Wnt-receptor complexes, or rescue experiments (e.g., Wnt3a addition) would provide stronger support for this proposed mechanism. Furthermore, we must address potential alternative explanations and genetic compensation within the sFRP family. The sFRP family exhibits significant functional redundancy [38,39]. For instance, transcriptomic analysis of FRZB-knockout mice in articular cartilage demonstrated a marked compensatory upregulation of Sfrp1 and Sfrp2 to maintain the extracellular Wnt inhibitory tone. While our siRNA-mediated knockdown in the C2C12 model produced a robust myogenic phenotype, we cannot rule out the possibility that the system triggers transcriptional adaptation, a molecular response often mediated by mutant mRNA degradation that upregulates related paralogs to buffer against the loss of a single factor [33,40]. Therefore, the role of FRZB should be viewed as part of a highly resilient and redundant regulatory network, rather than an isolated controller. Finally, although we focused on canonical Wnt signaling, FRZB and related sFRPs may also modulate non-canonical Wnt pathways (such as Wnt/Ca^2+^ or planar cell polarity routes) or cross-talk with growth pathways such as PI3K/AKT, MAPK, or integrin signaling. Exploring these axes might reveal additional regulatory mechanisms.
In summary, we demonstrate that FRZB suppresses myogenesis via extracellular competition with Wnt ligands, thereby shifting muscle cell behavior toward reduced proliferation, fusion, and hypertrophy. Considering the differential expression of FRZB among pig breeds and its known genetic variations, this gene represents a candidate locus for further investigations aimed at improving livestock muscle growth and carcass traits.
5. Conclusions
In conclusion, our findings identify Porcine FRZB (sFRP3) as a negative regulator of myogenesis. In vitro, FRZB knockdown alleviated the suppression of Wnt signaling, thereby enhancing C2C12 myoblast proliferation, migration, differentiation, and myotube fusion, while simultaneously shifting transcriptional programs toward hypertrophy and away from atrophy. The significantly higher FRZB expression detected in the fetal longissimus dorsi muscle of slow-growing pig breeds (TP and WJ) compared with fast-growing LW pigs suggests that elevated levels of this Wnt antagonist may impose a molecular constraint on muscle development. Collectively, these results provide a mechanistic basis for the breed-specific differences in muscle growth and highlight FRZB as a biologically plausible candidate gene for future applications in porcine molecular breeding and carcass trait improvement.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mohammadabadi M. Bordbar F. Jensen J. Du M. Guo W. Key Genes Regulating Skeletal Muscle Development and Growth in Farm Animals Animals 20211183510.3390/ani 1103083533809500 PMC 7999090 · doi ↗ · pubmed ↗
- 2Buckingham M. Vincent S.D. Distinct and dynamic myogenic populations in the vertebrate embryo Curr. Opin. Genet. Dev.20091944445310.1016/j.gde.2009.08.00119762225 · doi ↗ · pubmed ↗
- 3Wigmore P.M. Stickland N.C. Muscle development in large and small pig fetuses J. Anat.19831372352456630038 PMC 1171817 · pubmed ↗
- 4Reis E.P. Paixao D.M. Brustolini O.J. Silva F.F. Silva W. Araujo F.M. Salim A.C. Oliveira G. Guimaraes S.E. Expression of myogenes in longissimus dorsi muscle during prenatal development in commercial and local Piau pigs Genet. Mol. Biol.20163958959910.1590/1678-4685-gmb-2015-029527801482 PMC 5127148 · doi ↗ · pubmed ↗
- 5Knight J.D. Kothary R. The myogenic kinome: Protein kinases critical to mammalian skeletal myogenesis Skelet. Muscle 201112910.1186/2044-5040-1-2921902831 PMC 3180440 · doi ↗ · pubmed ↗
- 6Wei D. Zhang J. Raza S.H.A. Song Y. Jiang C. Song X. Wu H. Alotaibi M.A. Albiheyri R. Al-Zahrani M. Interaction of Myo D and Myo G with Myoz 2 gene in bovine myoblast differentiation Res. Vet. Sci.202215256957810.1016/j.rvsc.2022.09.02336191510 · doi ↗ · pubmed ↗
- 7Skapek S.X. Rhee J. Kim P.S. Novitch B.G. Lassar A.B. Cyclin-mediated inhibition of muscle gene expression via a mechanism that is independent of p RB hyperphosphorylation Mol. Cell. Biol.1996167043705310.1128/MCB.16.12.70438943359 PMC 231707 · doi ↗ · pubmed ↗
- 8Skapek S.X. Rhee J. Spicer D.B. Lassar A.B. Inhibition of myogenic differentiation in proliferating myoblasts by cyclin D 1-dependent kinase Science 19952671022102410.1126/science.78633287863328 · doi ↗ · pubmed ↗