Activity of Natural Substances and n-Undecyl-α/β-l-Fucopyranoside Against the Formation of Pathogenic Biofilms by Pseudomonas aeruginosa
Christian Dietrich Vogel, Anne Christine Aust, Raffael Christoph Wende, Undraga Schagdarsurengin, Florian Wagenlehner

TL;DR
This study explores natural substances and a fungal metabolite to inhibit biofilm formation by Pseudomonas aeruginosa on urinary catheters.
Contribution
The study identifies fungal metabolite terrein and n-undecyl-α/β-l-fucopyranoside as potential agents to prevent P. aeruginosa biofilms.
Findings
Terrein at 256 µg/mL reduced P. aeruginosa biofilm formation by 55.6%.
n-undecyl-α/β-l-fucopyranoside at 512 µg/mL inhibited biofilm formation by 58.3%.
The same compound increased biofilm formation in another P. aeruginosa strain at 256 µg/mL.
Abstract
Background/Objectives: Emerging biofilms of uropathogenic bacteria, particularly P. aeruginosa, on medical devices such as urinary catheters, lead to complications in the treatment of urinary tract infections (UTI). Considering the spread of antibiotic resistance, the search for alternative efficient control options for biofilms is of great medical interest. Methods: Curcumin, 1-monolaurin, n-undecyl-α/β-l-fucopyranoside, and the fungal metabolite terrein were investigated for their influence on biofilm formation by P. aeruginosa on latex catheter pieces in artificial urine (AU), monitoring the number of colony-forming units per cm Latex-Catheter (CFU/cm Latex-Catheter). Results: Significant inhibition of P. aeruginosa biofilm formation [55.6% CFU reduction/cm2] was observed with the fungal metabolite terrein at 256 µg/mL AU. At a concentration of 512 µg/mL AU, terrein achieved almost…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
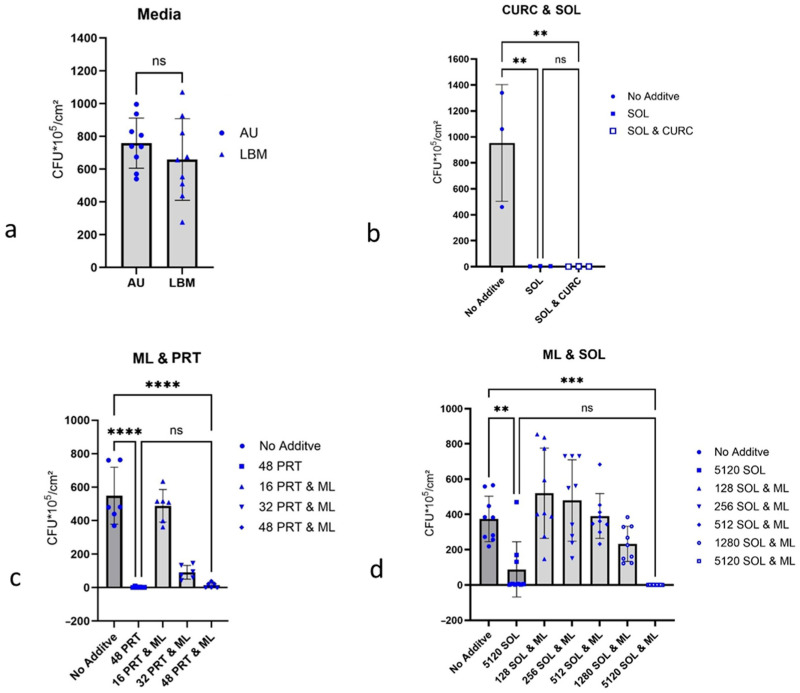 Figure 1
Figure 1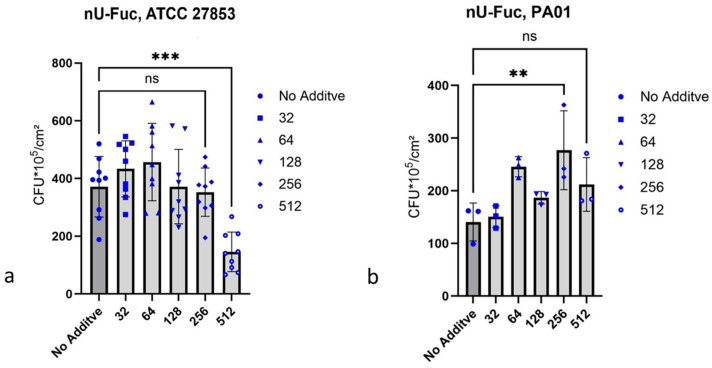 Figure 2
Figure 2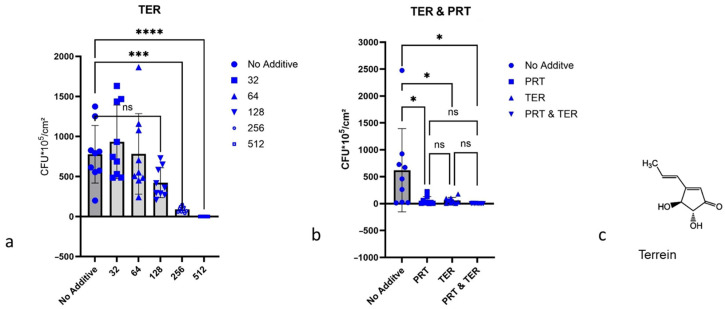 Figure 3
Figure 3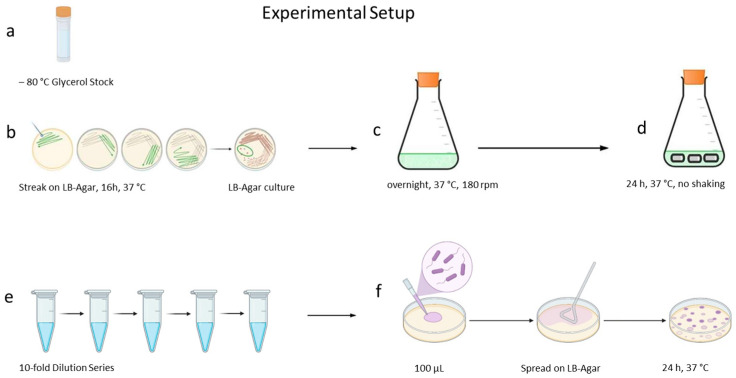 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTannin, Tannase and Anticancer Activities · Curcumin's Biomedical Applications · Biological Stains and Phytochemicals
1. Introduction
The biofilms of P. aeruginosa pose serious problems in everyday urological practice. 2 million people every year are affected worldwide, of which 90 thousand die from P. aeruginosa infection [1]. In this work, P. aeruginosa Strain ATCC 27853 serves as a test organism, which forms tenacious biofilms [2]. Biofilms are difficult to treat properly, including in the context of urinary tract infections. Urinary catheters cause approximately 75% nosocomial urinary tract infections [3,4,5]. Biofilm formation is a fast and complex process triggered by the nature of the surface, the interaction of individual bacterial cells with each other, Quorum Sensing [3,5,6], and receptor molecules on biological surfaces [7,8]. Within seconds, P. aeruginosa can increase the concentration of the second messenger c-di-GMP [9]. Once the biofilm is established, neither the body’s defences nor antibiotic treatment can eradicate bacteria, regardless of the susceptibility of biofilm-producing P. aeruginosa to a certain antibiotic [10,11]. The P. aeruginosa strain PA 01 reacts to the presence of aminoglycosides [12] in LB medium with increased formation of alginate, an exopolysaccharide or with increased biofilm formation. Micelles and liposomes stabilized with amphiphilic polymer components and loaded with antibiotics are already used in medicine. One stabilizing polymer with good biocompatibility used for stabilization of micelles or liposomes is Soluplus^®^, a polyvinyl caprolactam–polyvinyl acetate–polyethylene glycol graft copolymer [13]. The high dosing of classical antibiotics needed to treat biofilms can cause severe adverse events. For example, fluoroquinolone antibiotics can cause tendon ruptures [14], and the application of aminoglycoside antibiotics carries the risk of lifelong hearing loss [15]. Another issue is the emergence of antimicrobial resistance against antibiotics [16,17,18]. Due to this issue, plant-based agents, such as essential oils [19] or polyphenols, for example, curcumin [20], are investigated. Ding, T et al. observed the inhibiting and biofilm-mitigating effects of curcumin on Gram-negatives, including Pseudomonas spp. [21]. Moreover, the Food and Drug Administration (FDA) recognizes curcumin as safe [22]. Under light irradiation with a wavelength of 405 nm, curcumin showed an inhibition of biofilm formation by P. aeruginosa. A lower but still inhibiting effect without light irradiation was also observed [23]. 1-Monolaurin (glycerol-α-monolaurate) shows antibacterial effects, but only against Gram-positive bacteria [24,25]. We considered that detergents could shuttle 1-monolaurin through the outer membrane of the Gram-negative P. aeruginosa ATCC 27853. Due to the poor solubility of curcumin and 1-monolaurin, these were investigated together with Soluplus^®^ and, in the case of 1-monolaurin, additionally together with Prontosan^®^ (polyhexanide) for their inhibitory effect against biofilm formation by P. aeruginosa. Monovalent low molecular weight derivatives of l-fucose interact with LecB of P. aeruginosa and are possible inhibitors for biofilm formation by P. aeruginosa [26,27]. Therefore, we have synthesized n-undecyl-α/β-l-fucopyranoside according to the modified Fischer synthesis [28] and tested it as an inhibitor of biofilm formation by P. aeruginosa. The antibacterial agent terrein (4,5-dihydroxy-3-(1-propen-1-yl)-2-cyclopenten-1-one) from Aspergillus terreus acts effectively against the development of P. aeruginosa biofilms. It antagonizes quorum sensing of P. aeruginosa, acting as a dual inhibitor of QS and c-di-GMP signalling [29]. However, terrein does not affect P. aeruginosa cell growth [29]. The fungal metabolite terrein showed efficacy against biofilm formation by P. aeruginosa alone and in the presence of the detergent Prontosan^®^. The objective of this study is to evaluate effective alternatives to antibiotics for combating biofilm growth, especially on urological medical devices such as catheters.
2. Results
2.1. Chemical Synthesis of n-Undecyl-α/β-l-Fucopyranoside
The yield for the in-house synthetized n-undecyl-α/β-l-fucopyranoside was 2.297 g (7.213 mmol; 60%) related to the 1.96 g (11.95 mmol) of α/β-l-fucose used, and the α/β-ratio was ≈ 2:1, according to ^1^H NMR analysis (Supplementary Materials Figure S1).
2.2. MIC Values
The MIC values are shown in Table 1. For Piperacillin, the MIC value of 2–4 µg/mL in iron-depleted Mueller–Hinton broth, according to EUCAST standards, was confirmed with our result of 4 µg/mL AU as MIC for P. aeruginosa ATCC 27853. In combination with Soluplus^®^, the MIC for Piperacillin did not change. Surprisingly, the detergent Soluplus^®^ did not show any antibacterial effect. Prontosan^®^ showed an inhibitory effect against P. aeruginosa cells. The determined MIC value of 16/32 µg/mL AU corresponds to the value that is reported in the literature. For the MIC value of terrein, no comparable value in the literature was found. The combination of terrein and Prontosan^®^ resulted in a reduction in the MIC for terrein. 1-Monolaurin showed no antibacterial effect. The combination with Soluplus^®^ also did not cause any bacteriostatic effect of 1-monolaurin. We observed the same when combining curcumin with Soluplus^®^.
2.3. Dependence of Biofilm Formation on the Culture Medium
The primary culture of P. aeruginosa ATCC 27853, cultivated in AU and LBM, formed biofilms in both cases. The colony numbers from these biofilms grown in AU or LBM showed no significant difference (Figure 1a).
2.4. Growth Inhibition of P. aeruginosa Biofilms
2.4.1. Curcumin and 1-Monolaurin, Each with Detergent
Soluplus^®^ alone already showed a strong inhibitory effect, and in combination with curcumin, it masked the possible efficacy of curcumin (Figure 1b). Therefore, further tests with curcumin have not been carried out. 1-Monolaurin showed, in combination with Prontosan^®^, no additional effect (Figure 1c), with Soluplus^®^ additive inhibition against biofilm formation by P. aeruginosa ATCC 27853 proving to be insignificant (Figure 1d).
2.4.2. n-Undecyl-α/β-l-Fucopyranoside
n-Undecyl-α/β-l-fucopyranoside showed 58.3% inhibition of biofilm formation by P. aeruginosa ATCC 27853 at a concentration of 512 µg/mL (Figure 2a). In contrast, n-undecyl-α/β-l-fucopyranoside did not inhibit biofilm formation by P. aeruginosa PA 01, and at a concentration of 256 µg/mL, it caused an 87.0% increase in biofilm formation. Between n-undecyl-α/β-l-fucopyranoside concentrations of 64 µg/mL to 256 µg/mL, an increase in biofilm formation of 87% occurred (Figure 2b).
2.4.3. Terrein
Terrein showed 55.6% inhibition of biofilm formation by P. aeruginosa ATCC 27853 at a concentration of 256 µg/mL AU, and at 512 µg/mL AU, terrein caused nearly complete inhibition of biofilm formation (Figure 3a). It seems that terrein and the disinfectant Prontosan^®^, tested in combination against P. aeruginosa ATCC 27853 biofilm formation, have a synergistic effect against biofilm formation by P. aeruginosa ATCC 27853, but significance cannot be confirmed (Figure 3b).
3. Discussion
We investigated the impact of curcumin [21], n-undecyl-α/β-l-fucopyranoside [26,27,28], terrein [29,30], and 1-monolaurin [24,25] in terms of the inhibitory effect on the biofilm formation of P. aeruginosa ATCC 27853 and P. aeruginosa PA 01. The MIC values for the substances tested against P. aeruginosa ATCC 27853 showed that, for Soluplus^®^, the antibiofilm and bactericidal effects do not necessarily correlate. Soluplus^®^ neither reduces cell growth nor kills Pseudomonas cells; it prevents only the biofilm formation by P. aeruginosa ATCC 27853. The influence of the two media, LBM and AU, on the growth of P. aeruginosa ATCC 27853 was not significant. Considering the descriptions of Soluplus^®^ [13], we have chosen very high concentrations of Soluplus^®^ for pre-studies with curcumin, circumventing solubility issues with this substance in AU. Dissolving Soluplus^®^ or Prontosan^®^ in AU worked well in the high concentrations chosen. Concentrations of up to 512 µg curcumin in the presence of Soluplus^®^ resulted in stable suspensions. Due to their insolubility in the AU culture medium used, inhibition tests with curcumin or 1-monolaurin alone were not feasible in these preliminary studies. Curcumin, dissolved as a stem solution in a 50 mg/mL concentration in DMSO, precipitated in AU even in a 0.4 mg/mL concentration. However, both detergents overlap with the already proven antibacterial effects of curcumin or possibly of 1-monolaurin on P. aeruginosa. Both Soluplus^®^ and Prontosan^®^, at the concentrations applied, caused the near-complete inhibition of biofilm formed by P. aeruginosa. In the case of Soluplus^®^, the necessary concentrations used were too high for reasonable usage in medicine. Bioorganic excipients, such as milk powder, for example, may be an alternative to detergents for dissolving curcumin in AU and may facilitate the estimation of a possible inhibitory effect on P. aeruginosa biofilm formation on latex catheter surfaces. Unlike the detergents Soluplus^®^ or Prontosan^®^, they do not inhibit biofilm formation. Tested as a possible antilectin, n-undecyl-α/β-l-fucopyranoside led to a significant reduction in biofilm formation by P. aeruginosa ATCC 27853, whereas n-undecyl-α/β-l-fucopyranoside was ineffective against P. aeruginosa PA 01. The varying expressions of LecB in different P. aeruginosa strains could explain this. Due to the negative result obtained, further tests with n-undecyl-α/β-l-fucopyranoside against P. aeruginosa PA 01 were not performed. This result may be due to the high genomic diversity of P. aeruginosa among different isolates [31]. Terrein was considered nontoxic [30], and, therefore, it is a candidate for potential therapeutic use. Terrein showed a significant inhibitory effect against biofilm formation by P. aeruginosa ATCC 27853. Prontosan^®^ did not significantly increase the effect of terrein. The promising effects of terrein, observed by us as a novel inhibitor of biofilm formation, must be tested on further P. aeruginosa strains and on other uropathogenic bacteria. Furthermore, the cytotoxicity of terrein and n-undecyl-α/β-l-fucopyranoside on urogenital tract epithelial cells must be tested before medical use against biofilm-associated P. aeruginosa infections in urological practice. Due to the interference of terrein with breast cancer cells, toxicity studies with bladder epithelial cells are necessary to assess possible adverse effects on healthy eukaryotic cells [32]. To our knowledge, terrein and the easily accessible n-undecyl-α/β-l-fucopyranoside have not yet been tested for their potential to inhibit biofilm formation on latex catheter surfaces. Our focus of interest was to reveal the number of viable cells within a biofilm, so we did not apply any further methodology than CFU estimation.
4. Materials and Methods
4.1. Substances and Devices
4.1.1. Culture Media
Artificial urine (AU), as a medium for P. aeruginosa test cultures, was freshly prepared according to the recipe mentioned in the literature [19,33]. AU was sterilized by filtration through a 0.2 µm capsule filter (Thermo Fisher Scientific (Waltham, MA, USA) # Cat-No: 10526952) before the addition of autoclaved Tryptone Soy Broth (TSB, Oxoid Ltd., Basingstoke, UK) as a nutritional supplement. The other media used for the cultivation, storage, and testing of P. aeruginosa were glycerol ≥99% (Merck Millipore (Burlington, MA, USA) # CAS 56-81-5), Lysogeny Broth Medium (LBM) (Serva Electrophoresis GmbH (Heidelberg, Germany) # Cat-No. 48501.01), and Tryptone Soy Broth (TSB) (Oxoid # Code: CMO129B).
4.1.2. Chemicals and Supporting Material
Dulbecco’s phosphate-buffered saline (PBS) was purchased from Capricorn Scientific GmbH (Ebsdorfergrund, Germany, # Cat-No: PBS-1A). For chemical syntheses and their monitoring, α/β-l-fucose (Carl Roth # CAS No. 2438-80-4), pH indicator rod pH-Fix 0–14 (Carl Roth # Art. No. 0549.2), para-n-dodecyl benzene sulfonic acid (Sigma-Aldrich (St. Louis, MO, USA) # CAS No. 121-65-3), ethyl acetate (Sigma-Aldrich # CAS-No. 141-78-6), n-undecanol (Sigma-Aldrich # CAS-No. 112-42-5), ethanol ≥99,8% (Merck Millipore # CAS 64-17-5), phosphomolybdic acid monohydrate (Merck Millipore # CAS 51429-74-4), sodium hydroxide (Merck Millipore # CAS-No.: 1310-73-2), silica gel 60 (Merck Millipore # CAS No. 7631-86-9), and TLC silica gel 60 Aluminum Sheets 5 × 10 cm (Merck Millipore # Cat-No. 1.16835.0001) were used. Piperacillin (Sigma-Aldrich # CAS-No. 66258-76-2) served as a comparison to assess efficacy. As biofilm-inhibiting substances, curcumin (Merck Millipore # CAS 458-37-7; ≥80% purity), and 1-monolaurin (TCI (Tokyo, Japan) # CAS RN^®^: 142-18-7; >98% purity) were used. The other substances tested as inhibitors were Soluplus^®^ (BASF SE, Ludwigshafen am Rhein, Germany), Prontosan^®^ (B. Braun Melsungen AG, Melsungen, Germany), and terrein (AdipoGen # CAS-No. 582-46-7; ≥98% purity). For terrein and n-undecyl-α/β-l-fucopyranoside, synthesized by us, dimethyl sulfoxide (DMSO) (Sigma-Aldrich # CAS-No. 67-68-5) served as solvent. The substances tested as inhibitors of biofilm formation are shown in Table 2.
4.1.3. Devices
For incubating the bacterial cultures, an IncuLine IL 112 Prime.390-0910 (VWR International GmbH, Darmstadt, Germany) and a Shaking Incubator IKA^®^ KS 4000 i Control (IKA-Werke GmbH and Co. KG, Staufen, Germany) were used. To monitor the bacterial growth, a Laxco MicroSpek™ DSM-Series Cell Density Metre (LAXCO Bothell, Mill Creek, WA, USA) and semi-micro Cuvettes Polystyrene 45 × 12 mm (Sarstedt AG & Co.KG, Nümbrecht, Germany) were used. For catheter testing, Rüsch Gold Latex Catheters (Teleflex Medical GmbH, Fellbach, Germany) served as a substrate for biofilm growth. An ultrasonic device Bandelin SONOREX TK30 50 kHz (BANDELIN electronic GmbH & Co. KG, Berlin, Germany) and a Vortex Scientific Industries Vortex Genie 2 G560 S/N 2-417489 (Thermo Fisher Scientific GmbH, Langerwehe, Germany) were used to process the biofilms and subsequently determine the CFU count (see 4.7). The determination of the minimal inhibitory concentration (MIC) values was performed with 96-well plates checkerboard TC-Plate 96 Well flat-bottom Standard (Sarstedt AG & Co.KG, Nümbrecht, Germany) and a MultiScanGO (Thermo Fisher Scientific GmbH, Langerwehe, Germany).
4.2. Organisms
P. aeruginosa ATCC 27853 was obtained from ATCC (American Type Culture Collection; Manassas, VA, USA), and P. aeruginosa PA 01 from DSMZ (German Collection of Microorganisms and Cell Cultures; Braunschweig, Niedersachsen, Germany).
4.3. Culture Conditions
4.3.1. Primary Culture
The cultivation of P. aeruginosa and the biofilm inhibition studies on it were performed as follows: From the glycerol stock stored at −80 °C, an inoculum was streaked onto an LB agar plate using a sterile inoculating loop, according to the three-line method, and incubated for 16 h at 37 °C. For the culture in sterile liquid medium (LB medium or AU), 5 mL of medium was placed in a 100 mL Erlenmeyer flask. A single colony was then transferred from the fresh LB agar culture to the medium using a sterile inoculating loop. The culture mixture was then incubated overnight in a shaking incubator at +37 °C and 180 rpm (revolutions per minute).
4.3.2. Secondary Culture
Three 1 cm long catheter pieces were placed as three technical replicates in a sterilized 100 mL Erlenmeyer flask containing 8 mL of LB medium. The test substance was then added to the LB medium at the appropriate concentration. Finally, 100 µL of the P. aeruginosa ATCC 27853 primary culture, diluted with sterile PBS buffer to an optical density (OD_λ=600 nm_) of 0.02, was added. After incubation for 24 h at 37 °C, the catheter pieces were removed from the culture medium and were ready for processing (see Section 4.7).
4.4. Chemical Synthesis of n-Undecyl-α/β-l-Fucopyranoside
Firstly, 1.96 g α/β-l-fucose (11.95 mmol), 24.69 mL n-undecanol (20.49 g = 118.90 mmol), 1.832 mL (1.942 g = 5.98 mmol) para-n-dodecylbenzenesulfonic acid, and 190 μL fully desalinated H_2_O were stirred in a closed 25 mL Schott flask at 60 °C and 700 rpm for 24 h. The initial yellowish-white suspension was a slightly cloudy mixture after the reaction ended. To monitor the reaction process, thin layer chromatography was performed (eluent: ethyl acetate/ethanol 90:10). Unreacted n-undecanol was removed via column chromatography on silica gel eluting with ethyl acetate/ethanol (100:0 then 90:10) [28]. The product was a white crystalline solid. The yield was 60%, related to the α/β-l-fucose used; see 2.1. The identity of the product was confirmed by mass spectrometry (MW = 318.35 g) and nuclear magnetic resonance spectroscopy (NMR). HRMS (ESI): m/z calculated for C_17_H_34_O_5_Na [M + Na]^+^: 341.2299; found: 341.2297. ^1^H NMR (600 MHz, CD_3_OD): δ 4.73 (d, J = 2.7 Hz, 1 H, H-1α), 68% for n-undecyl-α-l-fucopyranoside, 4.17 (d, J = 7.5 Hz, 0.5 H, H-1β), 32% for n-undecyl-β-l-fucopyranoside, 3.97–3.91 (m, 1 H, H-5α), 3.83 (dt, J = 9.5, 6.8 Hz, 0.5 H, β–OCH_2_CH2), 3.74–3.70 (m, 2 H, H-2α/H-3α), 3.68–3.58 (m, 3 H, α–OCH2_CH_2/H-4α/H-6β), 3.51 (dt, J = 9.5, 6.7 Hz, 0.5 H, β–OCH2_CH_2), 3.47–3.41 (m, 2 H, α–OCH2_CH_2, H-4β, H-4β), 1.72–1.50 (m, 3 H, α–OCH_2_CH2/β–OCH_2_CH2), 1.43–1.23 (m, 25.5 H, α-CH2/β-CH2/H-6β), 1.21 (d, J = 6.6 Hz, 3 H, H-6α), 0.90 (t, J = 6.9 Hz, 4.5 H, α-CH3/β-CH3) ppm. ^13^C{^1^H} NMR (150 MHz, CD_3_OD): δ 104.8 (C-1β), 100.5 (C-1α), 75.2 (C-5β), 73.6 (C-4α), 73.1 (C-3β), 72.3 (C-2β), 71.8 (C-4β), 71.7 (C-3α), 70.8 (β–OCH_2_), 70.0 (C-2α), 69.3 (α–OCH_2_), 67.5 (C-5α), 33.1, 30.9, 30.8, 30.7, 30.6, 30.6, 30.6, 30.5, 27.3, 27.1, 23.7 (α-CH_2_/β-CH_2_), 16.8 (C-6β), 16.7 (C-6α), 14.4 (α-CH_3_/β-CH_3_) ppm. δ [ppm] indicates the chemical shift in an atomic nucleus relative to the solvent residual peak in a magnetic field, J [Hz] means the coupling constant between neighbouring atomic nuclei, and multiplet abbreviations mean the signal multiplicity. This indicates the number of adjacent atomic nuclei connected to neighbouring nuclei. More information can be found in the relevant textbooks on nuclear magnetic resonance spectroscopy [36].
4.5. Determination of Minimal Inhibitory Concentration
The MIC values were determined according to EUCAST standards [33,35] for antibacterial agents by broth dilution. From the agent to be tested, a two-fold dilution series with a volume of 160 µL per well was made in AU. Then, every well containing agent was inoculated with 10 µL of PBS buffer-diluted P. aeruginosa ATCC 27853 suspension (1:200 v/v). The agents tested, listed in alphabetical order with highest concentration in µg/mL [number shown in parentheses] were curcumin and Soluplus^®^ [512 (curcumin) and 5120 (Soluplus^®^)], 1-Monolaurin and Soluplus^®^ [512 (1-Monolaurin) and 5120 (Soluplus^®^)], Prontosan^®^ [512], Soluplus^®^ [5120], terrein [1024], and terrein and Prontosan^®^ [1024 (terrein) and 32 (Prontosan^®^)]. The concentration of the detergents Soluplus^®^ and Prontosan^®^ we held constant throughout the dilution series. Since we expected no inhibiting effect for n-undecyl-α/β-l-fucopyranoside, and curcumin, which is insoluble in aqueous media, no MICs were determined for these two substances. The medium without agent inoculation served as a positive control, and the medium without bacteria and agent served as a negative control. After overnight incubation at 37 °C, turbidity was measured at λ = 600 nm. MIC means the lowest concentration at which no additional turbidity (corresponding to bacterial growth) can be detected compared to the positive control. The MIC values shown are the median values calculated from three biological replicates. Each biological replicate consisted of three dilution series on 96-well plates as technical replicates, whose results were averaged. Piperacillin served as positive standard; we used it in every MIC test we performed (Table 1).
4.6. Catheter Biofilm Experiments
4.6.1. General Procedure Catheter Experiment
To 8 mL of medium, three 1 cm long catheter segments, as technical replicates, and the substance to be tested, 100 µL of a freshly cultivated (Figure 4a–c) PBS-buffer-diluted P. aeruginosa ATCC 27853 suspension, was added. Incubation for 24 h to 36 h at 37 °C followed, and, after processing, the grown biofilms, CFUs, were counted (see Section 4.7). Each test series was biologically replicated three times (Figure 4c–f).
4.6.2. Influence of the Culture Medium
Due to the results shown for Proteus mirabilis [37], differences in the growth behaviour of P. aeruginosa ATCC 27853 in AU and LB medium were estimated. To 8 mL of AU and LB medium without additives and to three 1 cm long latex catheter segments, respectively, P. aeruginosa ATCC 27853 suspension with an OD_λ=600nm_ = 0.02 in PBS buffer was added. After incubation for 24 h at 37 °C, the colonies were counted (Figure 4c–f).
4.6.3. Curcumin and 1-Monolaurin, Each in Combination with Soluplus®
To 8 mL of AU and three 1 cm long catheter segments, 512 µg/mL curcumin together with 5120 µg/mL Soluplus^®^ were added and incubated over 24 h at 37 °C in the presence of P. aeruginosa ATCC 27853 according to the general procedure. Three 1 cm long catheter segments and P. aeruginosa ATCC 27853 without an additive in AU and with 5120 µg/mL Soluplus^®^ without further additives served as control. In the same manner, 512 µg/mL AU 1-monolaurin together with 48 µg/mL AU Prontosan^®^ and 128 µg to 5120 µg/mL AU Soluplus^®^ were tested (Figure 4).
4.6.4. n-Undecyl-α/β-l-Fucopyranoside
n-Undecyl-α/β-l-fucopyranoside was tested in concentrations from 32 µg/mL AU to 512 µg/mL AU following the general procedure, against biofilm formation by P. aeruginosa ATCC 27853 and by P. aeruginosa PA 01 (Figure 4).
4.6.5. Terrein
Terrein, an antimicrobial metabolite from the mould A. terreus, was tested at concentrations from 32 µg/mL AU to 512 µg/mL AU following the general procedure (Figure 4). To estimate possible co-effects of terrein and Prontosan^®^, 256 µg/mL AU terrein together with 64 µg/mL AU Prontosan^®^ were tested for the mitigation of biofilm formation by P. aeruginosa ATCC 27853. Because keratinocytes tolerate Prontosan^®^ well in concentrations up to 175 µg/mL [34], we have chosen Prontosan^®^ as a co-agent instead of Soluplus^®^.
4.7. Estimation of Colony-Forming Units in the Biofilm
The biofilm processing procedure is as follows: After incubation, the 1 cm catheter pieces were transferred from the Erlenmeyer flask into an Eppendorf tube containing 1000 µL of PBS buffer using sterile tweezers and gently swirled. This was repeated two more times with 1000 µL of PBS buffer to ensure that all free-floating cells were washed out of the biofilm. To suspend the living cells within the biofilm, the catheter piece was sonicated at 50 kHz in 1000 µL of PBS buffer for 5 min, vortexed for 2 min, sonicated again for 5 min, and vortexed for 1 min. A ten-fold dilution series was prepared from the resulting biofilm suspension. Then, 100 µL of the suitable dilutions (10^−4^ to 10^−7^) were spread onto the LB agar plates. After incubating the LB agar plates at 37 °C for 16 h, the colonies were counted [34,38].
4.8. Statistical Analysis
For statistical analyses, we used GraphPad Prism 10; version: GraphPad Prism 10.4.2. Statistical significance was set as p ≤ 0.05, with α = 0.05. The degree of significance is shown using * for p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, and **** p ≤ 0.0001. For the statistical comparison of two data sets with GraphPad Prism 10.4.2 a two-tailed t-test was used. The one-way ANOVA test served for the analysis of more than three data sets. The ANOVA–Dunnett test served to compare multiple data sets with a single control. The Tukey test was used to compare every data set with every other.
5. Conclusions
The two active ingredients, n-undecyl-α/β-l-fucopyranoside and terrein, tested for the first time on latex catheters for urological practice, show an inhibition of biofilm formation by P. aeruginosa ATCC 27853. The chemically pure anomers of n-undecyl-α/β-l-fucopyranoside are necessary to specify their efficacy as inhibitors of biofilm formation by P. aeruginosa. Another goal for the future is to estimate the mitigating effects of terrein on established biofilms and test the potential of terrein to prevent the formation of biofilms by other bacterial strains.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Li X. Gu N. Huang T.Y. Zhong F. Peng G. Pseudomonas aeruginosa: A typical biofilm forming pathogen and an emerging but underestimated pathogen in food processing Front. Microbiol.202313111419910.3389/fmicb.2022.111419936762094 PMC 9905436 · doi ↗ · pubmed ↗
- 2Cao H. Lai Y. Bougouffa S. Xu Z. Yan A. Comparative genome and transcriptome analysis reveals distinctive surface characteristics and unique physiological potentials of Pseudomonas aeruginosa ATCC 27853 BMC Genom.20171845910.1186/s 12864-017-3842-z PMC 546726328606056 · doi ↗ · pubmed ↗
- 3Singha P. Locklin J. Handa H. A review of the recent advances in antimicrobial coatings for urinary catheters Acta Biomater.201750204010.1016/j.actbio.2016.11.07027916738 PMC 5316300 · doi ↗ · pubmed ↗
- 4Kranz J. Schmidt S. Wagenlehner F. Schneidewind L. Catheter-Associated Urinary Tract Infections in Adult Patients Dtsch. Arztebl. Int.2020117838810.3238/arztebl.2020.008332102727 PMC 7075456 · doi ↗ · pubmed ↗
- 5Zhu Z. Wang Z. Li S. Yuan X. Antimicrobial strategies for urinary catheters J. Biomed. Mater. Res. A 201910744546710.1002/jbm.a.3656130468560 · doi ↗ · pubmed ↗
- 6Rasamiravaka T. Labtani Q. Duez P. El Jaziri M. The formation of biofilms by Pseudomonas aeruginosa: A review of the natural and synthetic compounds interfering with control mechanisms Biomed. Res. Int.2015201575934810.1155/2015/75934825866808 PMC 4383298 · doi ↗ · pubmed ↗
- 7Öztürk F.Y. Darcan C. KariptaşE. The Determination, Monitoring, Molecular Mechanisms and Formation of Biofilm in E. coli Braz. J. Microbiol.20235425927710.1007/s 42770-022-00895-y 36577889 PMC 9943865 · doi ↗ · pubmed ↗
- 8Sauer K. Camper A.K. Ehrlich G.D. Costerton J.W. Davies D.G. Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm J. Bacteriol.20021841140115410.1128/jb.184.4.1140-1154.200211807075 PMC 134825 · doi ↗ · pubmed ↗