Salmonella enterica as a Complementary Model to LPS for Immune Stress in Weaned Piglets: Systemic and Intestinal Alterations
Li Dong, Zhiyan Liu, Wenxi Li, Changwei Zhang, Haoyang Yuan, Jun Liu, Hongrong Wang, Lihuai Yu

TL;DR
This study compares Salmonella infection and LPS in piglets, showing Salmonella causes more severe gut damage and systemic inflammation, making it a better model for immune stress.
Contribution
The study introduces an optimized Salmonella enterica infection model that better mimics natural infection effects compared to LPS in weaned piglets.
Findings
Salmonella infection caused more severe and sustained systemic inflammation and gut structural damage than LPS.
Oral administration of 2 × 109 CFU Salmonella enterica for 24 h created a reproducible and physiologically relevant immune stress model.
Salmonella infection led to villus perforations and mitochondrial damage not observed with LPS.
Abstract
The post-weaning period is a critical transition phase in pig production, during which piglets exhibit increased susceptibility to enteric pathogens and associated intestinal disorders. To investigate the underlying mechanisms, lipopolysaccharide (LPS), a component of bacterial cell walls, is frequently used to induce immune stress in experimental models. However, LPS administration alone fails to recapitulate the full spectrum of pathological changes caused by live bacterial infections. In this study, we established an oral infection model using Salmonella enterica (SE), a major enteric pathogen, and systematically compared its effects with those of LPS in weaned piglets. Our results demonstrated that while both challenges in duced immune activation and intestinal injury, SE infection provoked more severe and sustained systemic inflammation, along with distinctive gut structural…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
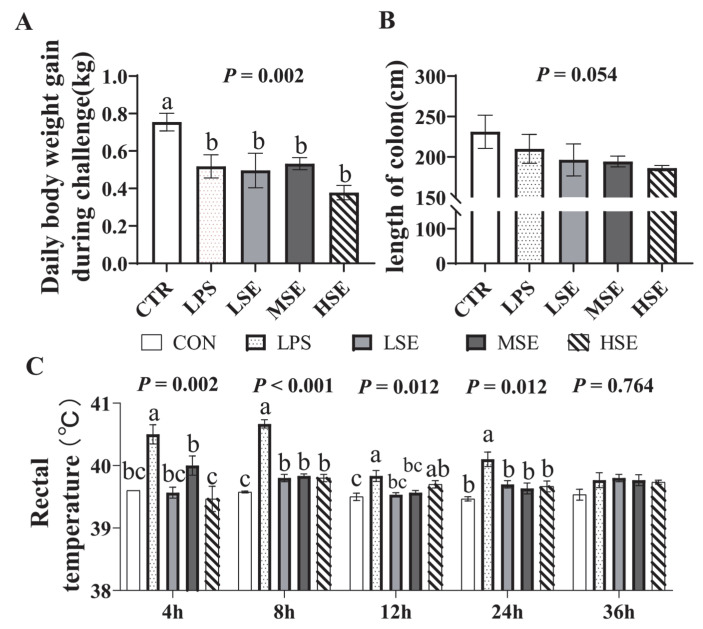 Figure 1
Figure 1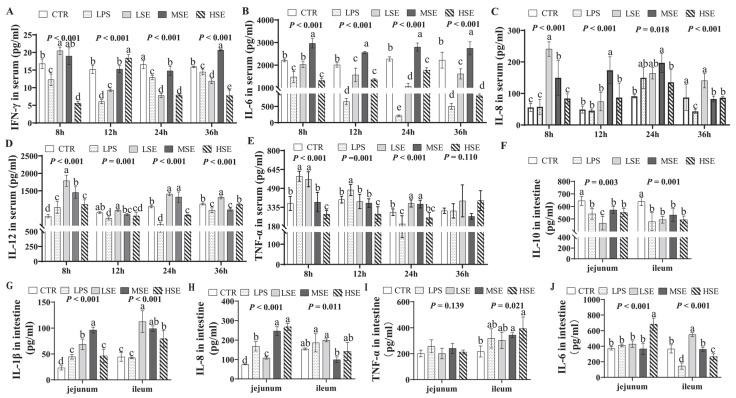 Figure 2
Figure 2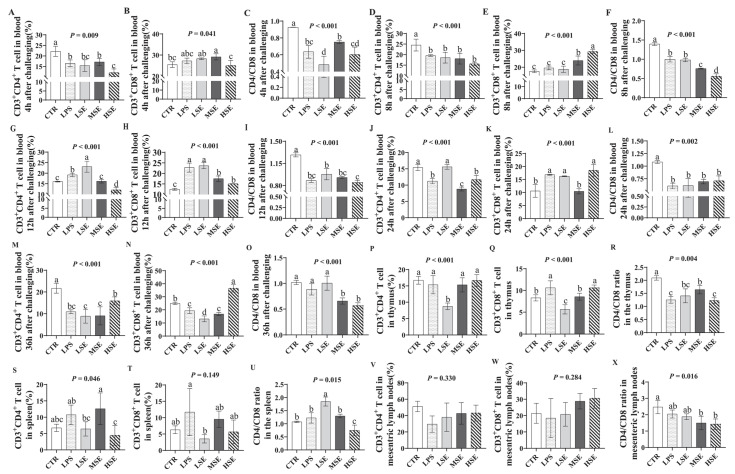 Figure 3
Figure 3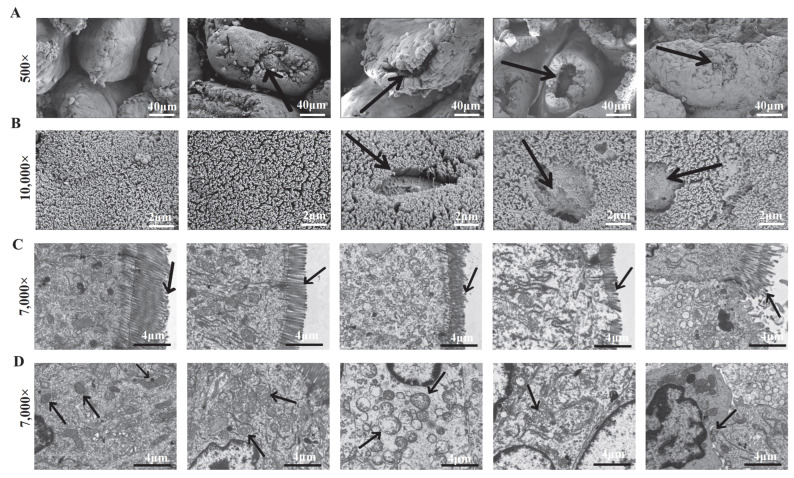 Figure 4
Figure 4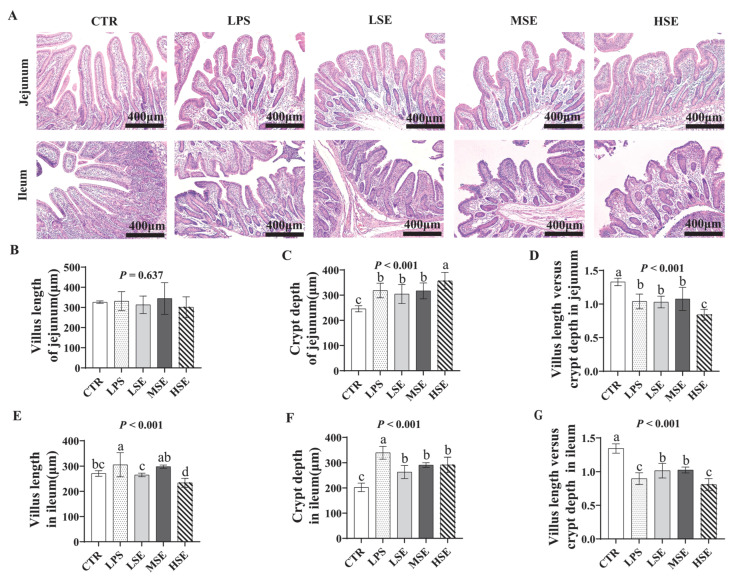 Figure 5
Figure 5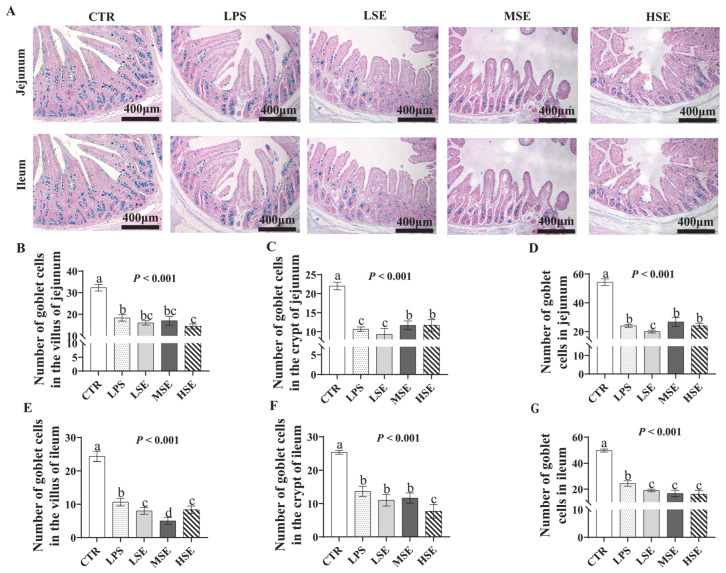 Figure 6
Figure 6- —National Natural Science Foundation of China
- —National Natural Science Foundation of China for Youth
- —Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · Animal Nutrition and Physiology · Immune Response and Inflammation
1. Introduction
Post-weaning stress represents a critical period in swine production, significantly increasing piglet susceptibility to enteric pathogens and associated immune dysregulation [1]. To study the ensuing pathophysiology, immune stress modeling in weaned piglets has primarily relied on lipopolysaccharide (LPS) challenge or live Escherichia coli infection. LPS, a key component of Gram-negative bacterial cell walls, is extensively utilized to induce acute systemic inflammation [2,3]. However, its model fidelity is limited, as it typically elicits transient systemic inflammation without reproducing the characteristic intestinal pathology of a live bacterial infection [4]. Conversely, while E. coli is also a common enteric pathogen, it often fails to provoke a robust and consistent immune activation in challenge models, limiting its utility for immune stress studies [5]. These constraints highlight a need for complementary models that better recapitulate the complex host responses to enteric pathogens, particularly in the intestinal compartment.
Salmonella enterica (SE) presents a promising complementary model, being a primary enteric pathogen capable of causing severe gastroenteritis in both animals and humans [6]. Its potential for modeling is underpinned by findings that SE challenge can induce significant pyrexia and marked alterations in serum immune markers [7,8]. Furthermore, the specific SE strain 50336 used in this study has demonstrated a pronounced ability to infect porcine jejunal epithelial cells (IPEC-J2) in vitro, indicating a high affinity for the porcine intestinal tract [9]. These findings suggest that oral SE challenge could effectively mimic key aspects of natural infection. More importantly, SE possesses virulence mechanisms, such as type III secretion systems (T3SS), that are absent in purified LPS and are known to drive more profound gut morphological damage, including villus perforations and disruption of tight junctions [10]. Consequently, an SE-based model may induce more severe and comprehensive intestinal pathophysiological alterations, providing a complementary approach to the classic LPS model, especially for studies focused on gut-specific immune responses and structural damage.
Therefore, this study aimed to directly address the limitations of existing models by systematically comparing the pathophysiological differences between the standard LPS challenge and an optimized oral SE infection in weaned piglets. We focused on a multidimensional assessment of systemic inflammation, intestinal immunity, and gut morphology to evaluate SE as a complementary model for immune stress in weaned piglets.
2. Materials and Methods
2.1. Bacterial Strain
The Salmonella enterica strain 50336 was provided by Professor Guoqiang Zhu (College of Veterinary Medicine, Yangzhou University).
2.2. Animals and Experimental Design
Forty piglets weaned at 21 days of age (28 days old at study initiation) were randomly allocated to five groups (n = 8/group): CTR (control), orally administered 10 mL of 0.9% saline; LPS, intraperitoneally injected with 100 μg/kg body weight LPS (E. coli O55:B5; Sigma L6529, Sigma-Aldrich, St. Louis, MO, USA); LSE, orally administered 1 × 10^8^ CFU/mL SE in 10 mL saline; MSE, orally administered 2 × 10^8^ CFU/mL SE in 10 mL saline; HSE, orally administered 3 × 10^8^ CFU/mL SE in 10 mL saline. The commercially available E. coli O55:B5 LPS was selected as it represents a standardized and widely referenced agent for inducing endotoxemia in swine, facilitating direct comparison with established models [4,11]. The LPS dose followed established protocols [4,11]. Piglets received a basal diet (Supplementary Materials Table S1; Yangzhou University Feed Factory) free of antibiotics, formulated according to the National Research Council (2012). The piglets were obtained from a commercial farm with a standard health management protocol but were not subjected to any vaccination program against Salmonella or other enteric pathogens prior to or during the trial. At 8, 12, 24, and 36 h post-challenge, rectal temperature was recorded and venous blood was collected. Six piglets per group were euthanized at 36 h for tissue sampling. The selected time points (8, 12, 24, and 36 h post-challenge) were based on the established kinetics of immune and pathophysiological responses in weaned piglet models [12,13,14].
2.3. Sample Collection
Blood samples were collected from the anterior vena cava using a sterile 21-gauge needle attached to a 5 mL syringe. The venipuncture site was disinfected with 70% ethanol prior to collection. Samples were collected at for cytokine analysis (IFN-γ, IL-6, IL-8, IL-12, TNF-α) and immune cell profiling. At 36 h, the abdominal cavity was opened, the intestine was separated, and the length of the duodenum, jejunum and ileum were measured. A 2-cm segment from the mid-section of the small intestine was rinsed with PBS, longitudinally incised, treated with 0.25% trypsin to remove mucus, and fixed in 2.5% glutaraldehyde for scanning and transmission electron microscopy. Segments (2 cm) from the mid-jejunum and mid-ileum were fixed in 4% formaldehyde for histological study. The intestinal contents of jejunum and ileum were flushed with pre-cooled PBS, the moisture was dried with filter paper, and the mucosa was gently scraped with a slide. The samples were then flash-frozen in liquid nitrogen for cytokine analysis. Fresh samples of the thymus, spleen, and mesenteric lymph nodes were collected immediately for T Lymphocyte subset analysis.
2.4. Cytokine Quantification
Serum and intestinal cytokine levels (IL-1β, IL-6, IL-8, IL-10, IL-12, TNF-α, IFN-γ) were measured using commercial ELISA kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China; Cat# H002, H008, H009, H010, H025, H052) according to the manufacturer’s protocols.
2.5. T Lymphocyte Subset Analysis
Heparinized whole blood (100 μL) was incubated with FITC-conjugated anti-CD4 (Solarbio K009445M; Solarbio Science & Technology Co., Ltd., Beijing, China) and PE-conjugated anti-CD8α (Solarbio K009446M; Solarbio Science & Technology Co., Ltd., Beijing, China; 10 μL each) for 30 min at 4 °C in dark. Erythrocytes were lysed with 3 mL of lysis buffer, samples were centrifuged (500× g, 5 min), and pellets were resuspended in 1% paraformaldehyde for flow cytometry [15]. Homogenates of thymus, spleen, and mesenteric lymph nodes were filtered through a 300-μm nylon mesh), centrifuged (100× g, 8 min, 4 °C), washed twice with PBS, and stained identically to blood samples.
2.6. Electron Microscopy
For scanning electron microscope (SEM): Fixed tissues were washed with 0.1 M PBS, post-fixed in 1% OsO_4_, dehydrated in a graded ethanol series (30–100%), critical-point dried, sputter-coated with gold, and imaged (ZEISS GeminiSEM 300; Carl Zeiss AG, Oberkochen, Germany) [16]. For transmission electron microscopy (TEM): Samples were fixed in 1% OsO_4_ (2 h, dark), dehydrated in ethanol/acetone, embedded in resin, polymerized (37 °C overnight; 60 °C, 48 h), ultrathin-sectioned, stained, and imaged (Hitachi HT7800; Hitachi High-Tech Corporation, Tokyo, Japan) [17].
2.7. Histological Analysis
Formaldehyde-fixed tissues were paraffin-embedded, sectioned (5 μm), and stained with hematoxylin and eosin (Solarbio G1120; Solarbio Science & Technology Co., Ltd., Beijing, China). Villus height, crypt depth, and villus height-to-crypt depth ratio (V/C) were quantified using Image-Pro Plus 6.0 (Media Cybernetics, Rockville, MD, USA) under an Olympus BX51 microscope (Olympus Corporation, Tokyo, Japan) [18].
2.8. Goblet Cell Staining
Goblet cells were stained with Alcian Blue (Solarbio G1560; Solarbio Science & Technology Co., Ltd., Beijing, China) according to the manufacturer’s instructions. Five intact villi per section were imaged (Nikon Eclipse E100, Nikon Instruments Inc., Tokyo, Japan), and goblet cells were quantified using Image J software 17.0.
2.9. Statistical Analysis
All data were analyzed using SPSS 25.0 (IBM Corp., Armonk, NY, USA). Group differences were assessed by one-way analysis of variance (ANOVA) followed by Duncan’s post hoc test for multiple comparisons. Figures were generated using GraphPad Prism 8.0.2 (GraphPad Software, San Diego, CA, USA). Statistical significance was established at p < 0.05.
3. Results
3.1. Body Weight, Colon Length and Rectal Temperature
Compared to the CTR group, the average daily weight gain (ADG) was significantly reduced in all challenged groups (LPS, LSE, MSE, HSE; p = 0.002; Figure 1A). Colon length tended to be shorter in SE-challenged groups compared to the CTR group (p = 0.054; Figure 1B). Rectal temperature increased significantly at 8 h post-challenge in the LPS and all SE groups compared to CTR (p < 0.001). By 12 h, significantly elevated temperatures persisted in the LPS and HSE groups (p = 0.012; Figure 1C).
3.2. Levels of Cytokines in Serum and Small Intestine
At 12 h post-stimulation, serum IFN-γ levels were significantly elevated in the HSE group (p < 0.05), while MSE-induced significant increases in serum IL-6 at 8, 12, 24, and 36 h (p < 0.05) (Figure 2A,B). Serum IL-8 levels were significantly higher in the LSE and MSE groups at 8 h, with sustained elevations in the MSE group at 12 and 24 h (p < 0.05) (Figure 2C). At 8 h, both LPS and SE groups showed significantly increased serum IL-12, and the LPS and LSE groups exhibited significantly higher serum TNF-α (p < 0.05) (Figure 2D,E). Conversely, the LPS group had significantly reduced serum IFN-γ and IL-6 at all time points, along with significantly lower IL-8 at 36 h (p < 0.05) (Figure 2A–C).
At 36 h post-challenge, jejunal and ileal IL-10 levels were significantly reduced in both LPS and SE groups relative to the CTR group (p < 0.05) (Figure 2F). In contrast, jejunal IL-1β was elevated in the LPS group, and both jejunal and ileal IL-1β were significantly increased in SE groups (p < 0.05) (Figure 2G). Jejunal IL-8 levels were significantly higher in the LPS, MSE, and HSE groups compared to CTR (p < 0.05) (Figure 2H). Additionally, the HSE group showed significantly increased ileal TNF-α and jejunal IL-6, while the LSE group had elevated ileal IL-6 (p < 0.05 vs. CTR) (Figure 2I,J).
3.3. T-Lymphocyte Subsets in Blood and Immune Organs
In blood, the CD4^+^/CD8^+^ T lymphocyte ratio was significantly decreased in the LPS and SE groups at 4, 8, 12, 24, and 36 h post-challenge (p < 0.05) (Figure 3A–O). At 36 h, this ratio was also significantly lower in the thymus, spleen, and mesenteric lymph nodes of LPS- and SE-challenged piglets compared to the control group (p < 0.05) (Figure 3P–X).
3.4. Intestinal Morphology
Scanning electron microscopy revealed villus tip damage in the small intestine of LPS and SE groups (Figure 4A), with surface perforations specific to SE-challenged piglets (Figure 4B). Microvilli were shorter in LPS and SE groups than in CTR, and SE groups exhibited disorganized microvilli in a dose-dependent manner (Figure 4C). Mitochondrial swelling was observed in both LPS and SE groups, with more severe damage in SE groups, including vacuolated cristae (Figure 4C,D). Hematoxylin–eosin (HE) staining showed significantly reduced villus height/crypt depth (V/C) ratios in the jejunum and ileum of LPS and SE groups compared to CTR (p < 0.05) (Figure 5).
3.5. Number of Goblet Cells
In the jejunum and ileum, the number of goblet cells in villi and crypts was significantly lower in LPS and SE groups than in CTR (p < 0.05) (Figure 6).
4. Discussion
Lipopolysaccharide (LPS) remains a cornerstone for modeling intestinal inflammatory injury, particularly in simulating Gram-negative bacterial infections such as Salmonella enterica (SE) [2,3]. However, our findings demonstrate that the immunological and pathophysiological distinctions between purified endotoxin exposure and live bacterial invasion are substantial, especially in the intestinal compartment. We systematically evaluated these responses in weaned piglets and demonstrated that oral SE infection (optimally at 2 × 10^9^ CFU) offers a complementary model. This model captures essential features of natural gut infection that LPS alone does not fully represent. The results of this study supported the utility of the SE model as a valuable adjunct to existing paradigms, particularly for studies focusing on intestinal pathology and sustained local immune activation.
The observed suppression of average daily gain (ADG) in both LPS- and SE-challenged piglets reflects the metabolic burden of systemic inflammation. This growth impairment likely stems from impaired nutrient absorption due to villus shortening and mitochondrial dysfunction [19], combined with pro-inflammatory cytokine-mediated anorexia [20]. Notably, SE infection induced more severe growth retardation and a unique reduction in colon length, aligning more closely with clinical reports of Salmonella-associated enteropathy characterized by persistent diarrhea and malabsorption [21]. The rectal temperature profiles further distinguished these stressors. LPS triggered a rapid, sharp hyperthermia (>40.5 °C at 8 h), characteristic of acute endotoxin shock mediated primarily via TLR4/MyD88-dependent signaling [22]. In contrast, SE elicited a more dose-dependent and sustained pyrexia, underscoring its engagement of multiple pattern recognition receptors (PRRs), including NLRs and cytosolic sensors, a hallmark of live bacterial invasion [23].
The cytokine responses unveiled a fundamental divergence in immune activation kinetics. The response to LPS was biphasic: an early surge in TNF-α and IL-12 was followed by the sustained suppression of IFN-γ, IL-6, and IL-8. This pattern is a classic manifestation of endotoxin tolerance, a compensatory mechanism that prevents uncontrolled inflammation [24]. This pattern starkly contrasts with the sustained, robust elevation of multiple pro-inflammatory cytokines, particularly IL-6 and IL-8, observed at 24 h post-SE infection, especially in the MSE group. This sustained response indicates persistent bacterial antigen presentation and inflammasome activation [25], more accurately mimicking the ongoing immune battle during a natural infection. Critically, the 2 × 10^9^ CFU SE dose generated optimal and reproducible cytokine kinetics for modeling. The spatial heterogeneity in intestinal cytokines, with ileal IL-6 dominating in low-dose SE and jejunal IL-8 prevailing at higher doses, may reflect differential microbial density or epithelial Toll-like receptor expression along the intestinal tract [26]. The concurrent suppression of the anti-inflammatory cytokine IL-10 in both jejunum and ileum across challenged groups underscores a critical disruption of the anti-inflammatory feedback loop, likely permitting uncontrolled neutrophil infiltration via IL-8-mediated chemotaxis [27].
A key finding was the significantly reduced CD4^+^/CD8^+^ T cell ratios across peripheral blood and all examined lymphoid organs (thymus, spleen, mesenteric lymph nodes). This signifies a profound, systemic immunosuppression induced by both challenges. Mechanistically, TNF-α overexpression can promote CD4^+^ T cell apoptosis [28], while chemokine gradients may sequester CD8^+^ T cells at inflamed intestinal sites [29]. The observed thymic involvement suggests a potential disruption in T cell ontogeny [30]. Moreover, dysfunction in the mesenteric lymph nodes, which are critical for intestinal immune surveillance, would severely compromise local immune responses [31]. This multi-organ immune impairment heightens susceptibility to secondary infections, a finding with critical implications for swine health management.
While both LPS and SE induced common morphological injuries like villus blunting, reduced V/C ratio, and microvillus atrophy, SE infection uniquely caused severe ultrastructural lesions. The dose-dependent villus perforations and mitochondrial cristae vacuolization observed specifically in SE-challenged piglets indicated direct epithelial cytotoxicity that extends beyond the effects of purified endotoxin. These distinctive pathological features are consistent with the action of Salmonella’s type III secretion systems (T3SS), which are known to disrupt tight junctions by redistributing claudin-1 and occludin [32]. The observed depletion of goblet cells across challenged groups further compromises the mucus barrier, but this effect was amplified in a dose-dependent manner by SE infection, facilitating potential pathogen translocation.
Collectively, our data robustly establish that oral challenge with 2 × 10^9^ CFU SE at 24 h post-infection constitutes a physiologically superior model for immune stress in weaned piglets. This model elicits severe and sustained systemic and intestinal immune activation, induces characteristic morphological damage in the gut, and leads to measurable growth suppression, thereby closely mimicking key features of natural infection. The resulting growth impairment and intestinal pathology translate directly into economic losses in swine production through reduced feed efficiency and increased morbidity. The optimized SE challenge model presented here offers a valuable platform for screening nutritional and therapeutic interventions aimed at mitigating such losses, ultimately supporting strategies to enhance gut health and improve productivity. It should be noted that while the sample size (n = 6 per group) followed established standards in porcine challenge models and was sufficient to detect significant differences in primary outcomes [33,34], a formal a priori power calculation was not conducted. Future studies applying this model, especially when evaluating interventions with subtler effects, would benefit from such calculations to strengthen experimental design. Finally, this study focused on acute responses within 36 h; longer-term observations would help clarify recovery dynamics and adaptive immunity. The model also remains to be tested with different Salmonella strains. Moving forward, this platform is well-suited for assessing feed additives, vaccines, and probiotics aimed at improving gut health. Deeper mechanistic insights could be gained by integrating transcriptomic or metabolomic approaches, and by evaluating specific interventions such as those targeting IL-10 pathway restoration [35] or mitochondrial integrity protection [36], with the goal of reducing post-weaning gastrointestinal morbidity in commercial swine production.
5. Conclusions
In summary, this study demonstrates that while both LPS challenge and Salmonella enterica (SE) infection induced immune stress in weaned piglets, oral SE infection provoked a more severe and comprehensive pathophysiological response. SE infection resulted in more pronounced growth impairment, sustained systemic inflammation, and profound intestinal damage, including distinctive lesions such as villus perforations and mitochondrial vacuolization not observed in the LPS model. Crucially, an oral dose of 2 × 10^9^ CFU SE for 24 h was optimized to establish a robust and physiologically relevant immune stress model. This validated protocol provides a potential tool for future research on intestinal immunology and for evaluating nutritional and therapeutic interventions in weaned piglets.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Fang L.H. Jin Y.H. Do S.H. Hong J.S. Kim B.O. Han T.H. Kim Y.Y. Effects of dietary energy and crude protein levels on growth performance, blood profiles, and nutrient digestibility in weaning pigs Asian-Australas. J. Anim. Sci.20183255610.5713/ajas.18.029430145868 PMC 6409451 · doi ↗ · pubmed ↗
- 2Bertani B. Ruiz N. Function and biogenesis of lipopolysaccharides Ecosal Plus 20188 e 10-112810.1128/ecosalplus.esp-0001-2018 PMC 609122330066669 · doi ↗ · pubmed ↗
- 3Kayagaki N. Wong M.T. Stowe I.B. Ramani S.R. Gonzalez L.C. Akashi-Takamura S. Miyake K. Zhang J. Lee W.P. Muszyński A. Noncanonical Inflammasome Activation by Intracellular LPS Independent of TLR 4Science 20133411246124910.1126/science.124024823887873 · doi ↗ · pubmed ↗
- 4Wyns H. Plessers E. De Backer P. Meyer E. Croubels S. In vivo porcine lipopolysaccharide inflammation models to study immunomodulation of drugs Vet. Immunol. Immunopathol.2015166586910.1016/j.vetimm.2015.06.00126099806 · doi ↗ · pubmed ↗
- 5Liu Y. Song M. Che T.M. Lee J.J. Bravo D. Maddox C.W. Pettigrew J.E. Dietary plant extracts modulate gene expression profiles in ileal mucosa of weaned pigs after an Escherichia coli infection J. Anim. Sci.2014922050206210.2527/jas.2013-642224663182 · doi ↗ · pubmed ↗
- 6Torrence M.E. Introduction to preharvest food safety Preharvest Food Safety John Wiley & Sons Washington, DC, USA 201811710.1128/9781555819644.ch 1 · doi ↗
- 7Singh M. Vohra H. Kumar L. Ganguly N.K. Induction of systemic and mucosal immune response in mice immunised with porins of Salmonella typhi J. Med. Microbiol.199948798810.1099/00222615-48-1-799920129 · doi ↗ · pubmed ↗
- 8Roy M.-F. Riendeau N. Bédard C. Hélie P. Min-Oo G. Turcotte K. Gros P. Canonne-Hergaux F. Malo D. Pyruvate kinase deficiency confers susceptibility to Salmonella typhimurium infection in mice J. Exp. Med.20072042949296110.1084/jem.2006260617998386 PMC 2118530 · doi ↗ · pubmed ↗