Comprehensive Identification and Male-Biased Expression Analysis of Odorant-Binding Protein Genes in the Hawaiian Flower Thrips, Thrips hawaiiensis (Thysanoptera: Thripidae)
Qingqing Fan, Yanjun Li, Xiaodi Hu

TL;DR
This study identifies 12 odorant-binding protein genes in the Hawaiian flower thrips, finding they are more active in males, suggesting a role in male-specific behaviors like mating and foraging.
Contribution
The first comprehensive identification and expression analysis of odorant-binding protein genes in Thrips hawaiiensis, revealing male-biased expression patterns.
Findings
Thrips hawaiiensis has 12 OBP genes, a number lower than most insects but comparable to other thrips.
OBPs and CSPs show male-biased expression, suggesting roles in male-specific olfactory behaviors.
The study provides a foundation for future functional studies of olfactory proteins in insects.
Abstract
The Hawaiian flower thrips, Thrips hawaiiensis, is a widespread pest that infests the flowers of numerous horticultural crops. We characterized the previously unknown olfactory system of Thrips hawaiiensis by analyzing its odorant-binding proteins (OBPs). Our genome survey revealed 12 OBP genes, a count comparable to other thrips but low relative to most insects. Subsequent transcriptomic and RT-qPCR analyses identified consistent male-biased expression, implicating these OBPs in male-specific olfactory behaviors, such as mate location and foraging. We further identified 11 CSPs, the majority of which showed a male-biased expression pattern similar to that of the OBPs. Olfaction is essential for key insect behaviors, such as host-seeking and mating, and is initiated by odorant-binding proteins (OBPs), which bind and transport hydrophobic odors. Thrips hawaiiensis is a major pest that…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
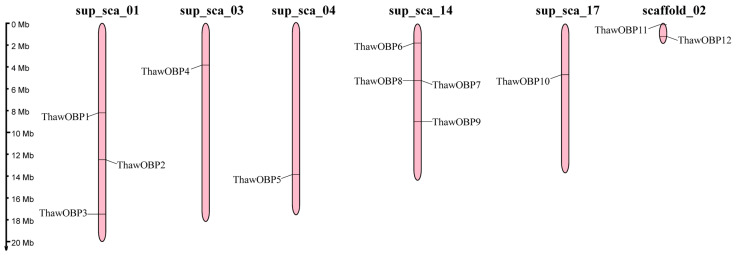 Figure 1
Figure 1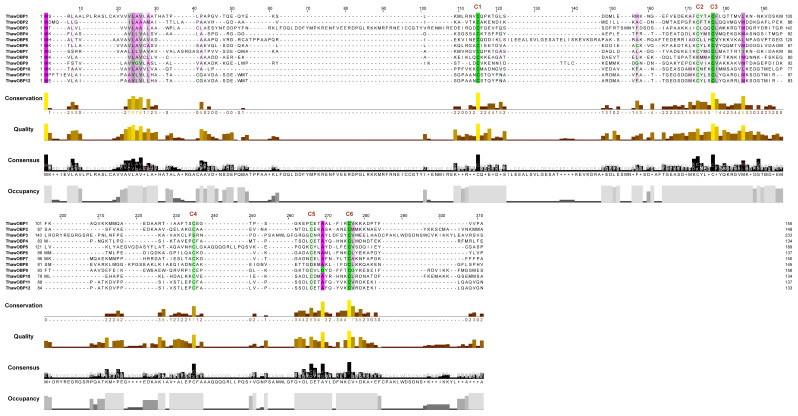 Figure 2
Figure 2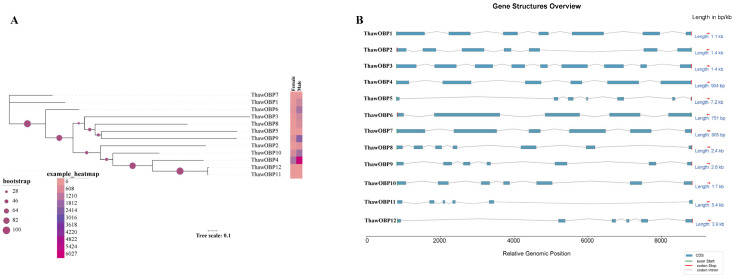 Figure 3
Figure 3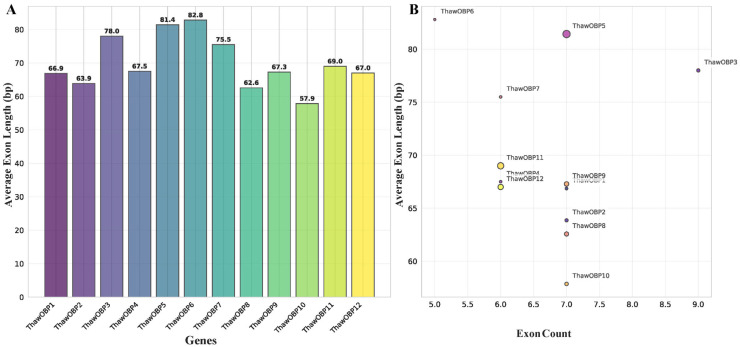 Figure 4
Figure 4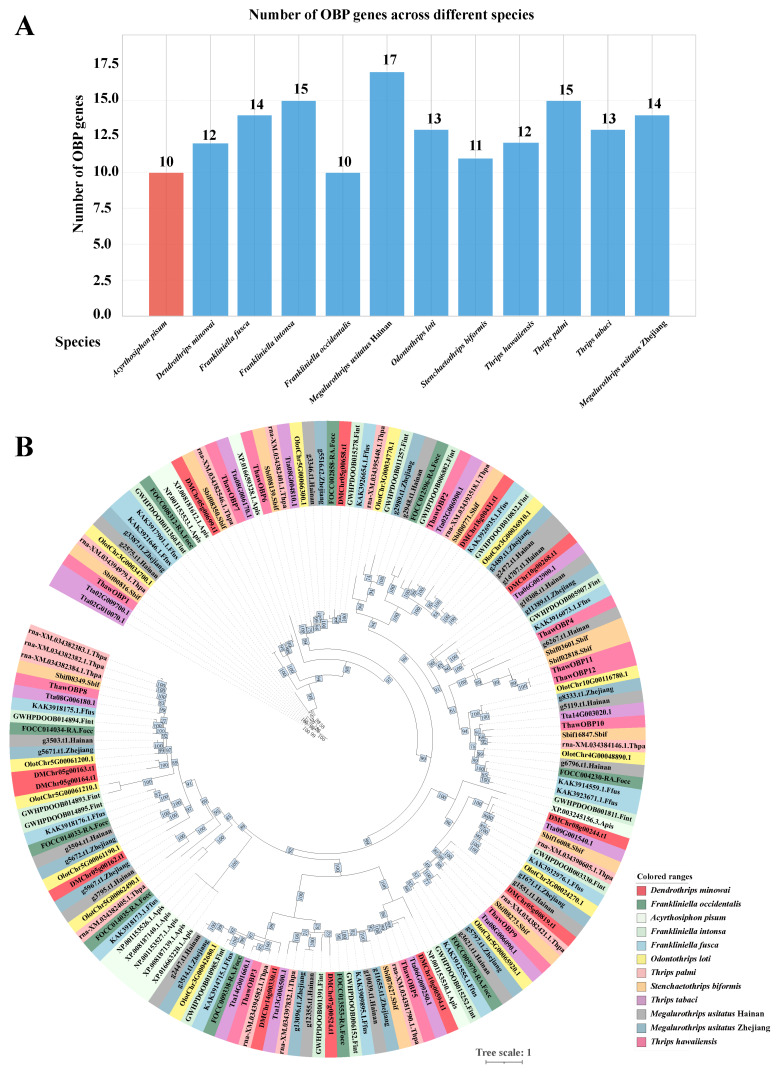 Figure 5
Figure 5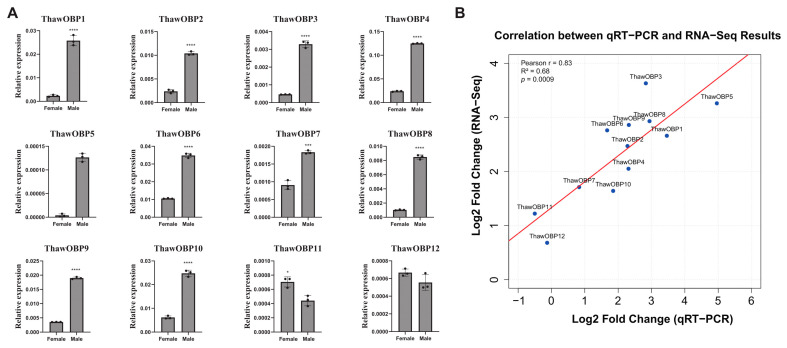 Figure 6
Figure 6- —Ningbo Yongjiang grant
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurobiology and Insect Physiology Research · Insect-Plant Interactions and Control · Olfactory and Sensory Function Studies
1. Introduction
Thrips hawaiiensis (Morgan) (Thysanoptera: Thripidae), a common flower-dwelling thrips species native to the Oriental and Pacific regions, has expanded its geographical range through international trade to Africa, Australia, Europe, and the Americas [1,2,3,4,5]. T. hawaiiensis primarily feeds and resides within floral tissues [1,5,6]. The elevated risk of global introduction and spread of T. hawaiiensis, driven by its strong thigmokinetic behavior, high fecundity, and short life cycle [1,7], is fueling growing concern about its potential to become a major pest of many crops. As a strictly flower-dwelling species, T. hawaiiensis inflicts damage on a wide range of crops, including tobacco [8], rose [9], gladiolus [10], Brassica oleracea [11], coffee [12], mango [13], citrus [14], apples and pears [6], and bananas [15,16]. Direct feeding on floral and fruit tissues induces symptoms such as scarring, necrosis, and malformation, the severity of which depends on infestation intensity. Furthermore, its pollen-feeding behavior can adversely affect plant fertility [17].
Olfaction is essential for detecting and interpreting environmental cues, enabling organisms to locate food, hosts, mates, and oviposition sites, as well as to evade predators and pathogens [18]. Both invertebrates and vertebrates use the olfactory system to mediate odorant detection [19]. The accurate operation of the olfactory system depends on a coordinated series of olfactory proteins. Key among these are odorant-binding proteins (OBPs), chemosensory proteins (CSPs), odorant receptors (ORs), gustatory receptors (GRs), ionotropic receptors (IRs), sensory neuron membrane proteins (SNMPs), odorant-degrading enzymes (ODEs), and Niemann–Pick protein C2 (NPC2) [20,21]. Notably, OBPs participate in the initial biochemical reaction of the olfactory transduction cascade. They play a critical role by discriminating, binding, and transporting odorant molecules to their corresponding Ors [22,23,24]. OBPs are characterized as small, water-soluble, and extracellular, residing in the sensillar lymph that bathes the sensory dendrites [19,25]. The classification of OBPs is based on their conserved cysteine patterns, yielding distinct subfamilies: Classic (six cysteines), Minus-C (four or five), Plus-C (at least eight), and Dimer/Atypical OBPs, the latter two comprising tandem or fused classic domains that may form dimer-like structures [23,26,27,28]. The first OBP was identified in Antheraea polyphemus [29]. Subsequently, transcriptomic and genomic data have facilitated the identification of a rapidly increasing number of OBPs. Although OBPs have been characterized in several thrips species, including Frankliniella occidentalis [30], Frankliniella intonsa [30], and Megalurothrips usitatus [26], this gap persists for Thrips hawaiiensis. The recent availability of a genome assembly for T. hawaiiensis [31] provides a crucial resource to address this gap, allowing for a comprehensive exploration and comparative analysis of its OBP repertoire.
We conducted a comprehensive analysis of odorant-binding protein (OBP) genes in Thrips hawaiiensis, beginning with genome-wide identification. This was followed by characterization of their chromosomal distribution, phylogenetic evolution, gene structures, and protein domains. Tissue-specific expression profiling across both sexes was performed via RT-qPCR. Collectively, our results lay the necessary groundwork for elucidating the functional roles of OBPs and for devising targeted, eco-friendly control methods for this pest.
2. Materials and Methods
2.1. Identification of OBP Genes in Thrips
The genome assembly, along with its GFF annotation, coding sequence (CDS), and protein sequence files for T. hawaiiensis, were obtained from figshare (https://doi.org/10.6084/m9.figshare.26125162) [31]. To investigate the prevalence of OBPs in thrips, we also retrieved publicly available whole-genome data of thrips (as of that available from June 2025), including two other populations of Megalurothrips usitatus (samples from Hainan, China [32], and Zhejiang, China [33]), Dendrothrips minowai [34], Frankliniella fusca [35], Frankliniella occidentalis [36], Odontothrips loti [37], Thrips palmi [38], Thrips tabaci [39], Frankliniella intonsa [40], and Stenchaetothrips biformis [41]. Known OBP amino acid sequences from Frankliniella intonsa and Frankliniella occidentalis [30] were retrieved from GenBank (National Center for Biotechnology Information, NCBI) and used as query sequences for a BLASTP (v2.5.0) search against the thrips amino acid database, applying an identity cutoff of 30%. Then, gene annotation was performed using hmmscan (v3.1b1) with the Pfam-A.hmm database, applying an E-value threshold of 1 × 10^−5^ to identify putative OBP genes (PF01395.18). To improve accuracy, the hmmscan results were combined with BLAST-based annotations by retaining only overlapping gene predictions, yielding a high-confidence gene set for downstream analyses. The putative N-terminal signal peptides of T. hawaiiensis OBPs were identified using the SignalP 6.0 online server (https://services.healthtech.dtu.dk/services/SignalP-6.0/, accessed on 28 October 2025) [42]. The OBP sequences were aligned using MUSCLE (v5.1) [43] software. The resultant alignment file was then imported into Jalview software (v2.11.5.0) [44] for visualization.
2.2. Characteristics Analysis of OBPs
The OBP sequences of thrips were aligned using MUSCLE (v5.1) [43]. Following alignment, a maximum likelihood (ML) phylogenetic tree was constructed on the aligned dataset with IQ-TREE (v2.2.5) [45], employing 1000 ultrafast bootstrap replicates (-B 1000). The chromosomal locations of OBP genes were retrieved from the T. hawaiiensis GFF annotation file using a custom Python (v3.10.13) script. They were subsequently visualized on the chromosomes via the online tool MapGene2Chrom (http://mg2c.iask.in/mg2c_v2.1/, accessed on 28 October 2025) [46,47]. The exon–intron structures of the ThawOBP genes were determined through an in-house Python script.
2.3. Insect Sample Collection
A laboratory population of Thrips hawaiiensis was established from individuals collected on kidney beans (Phaseolus vulgaris) at the Institute of Plant Protection, Fujian Academy of Agricultural Sciences, China (119°34′ E, 26°13′ N), and has been continuously reared on this host since 2016. The kidney bean diet was prepared by soaking in water, coating with a 10% honey solution, and air-drying. Thrips were maintained in MGC-350HP artificial climate incubators (Yiheng Scientific Instruments, Shanghai, China) under the following conditions: 27 ± 1 °C, 60 ± 5% RH, and a 16:8 (L:D) photoperiod.
2.4. RNA Extraction and Real-Time Quantitative PCR Analysis of ThawOBPs
A total of 300 adult male and female T. hawaiiensis samples were selected for analysis of relative mRNA expression levels. Total RNA was extracted from the samples using the Trizol method. Subsequently, the RNA was reverse transcribed into cDNA using a reverse transcription kit (Accurate Biology, Changsha, Hunan, China).
A primer design tool (https://www.ncbi.nlm.nih.gov/tools/primer-blast/, accessed on 28 October 2025) was used to design primers for the ThawOBP gene of Thrips hawaiiensis, with β-actin as the internal reference gene [48]. RT-qPCR was performed on an ABI QuantStudio 5 system (Thermo Fisher Scientific, Waltham, MA, USA), with the entire process conducted on ice. Each experiment included at least two technical replicates and three biological replicates. The expression levels in male and female Thrips hawaiiensis were calculated using the 2^−ΔCT^ method. Data analysis and visualization were conducted using GraphPad Prism (v9.5.0) and R (v4.2.0) for correlation calculations. All the primers used in this study are listed in Table 1.
2.5. Transcriptome Sequencing
To obtain a comprehensive overview of gene expression, total RNA was isolated from adult male and female subjects. Criteria for cDNA library qualification are as follows: the AD260/280 absorbance ratio should be between 1.8 and 2.0, the A260/230 ratio should be between 1.9 and 2.4, and the concentration measured by Qubit should be between 0.95 and 3.0. Each biological replicate comprised a pool of approximately 800 individuals. RNA extraction was performed using TRIzol Reagent (Thermo Fisher Scientific, USA). Subsequently, Illumina paired-end libraries were constructed with the TruSeq RNA Library Preparation Kit (Illumina, San Diego, CA, USA) according to the manufacturer’s instructions and sequenced on an Illumina NovaSeq 6000 platform. This generated approximately 14.3 GB of high-quality 150 bp paired-end sequence data.
2.6. Phylogenetic Analysis of OBP Genes
A maximum likelihood phylogeny was reconstructed using IQ-TREE (v2.3.3) [45]. To determine the most appropriate substitution model, we used the built-in ModelFinder tool, which selected the optimal model from a candidate set based on the Bayesian Information Criterion. The analysis was subsequently conducted under the best-fit model, WAG + R4. Branch support was assessed using ultrafast bootstrap approximation, with 1000 replicates.
3. Results
3.1. Identification and Sequence Analysis of OBPs in T. hawaiiensis
The genome of T. hawaiiensis was assembled using a hybrid approach that integrated the technologies of Oxford Nanopore long-read sequencing, Illumina short-read sequencing, and Hi-C chromatin conformation capture. This strategy produced a final assembly of 287.59 Mb, with a scaffold N50 of 13.84 Mb. According to BUSCO analysis, the assembled genome exhibits a high completeness of 98.7% [31]. As shown in Table 2, a total of 12 ThawOBP genes were identified through our BLASTP analysis. The genes, named ThawOBP1-ThawOBP12 based on their chromosomal locations (Figure 1), each possessed a complete open reading frame (ORF). The putative proteins range from 133 (ThawOBP12) to 233 (ThawOBP3) amino acids, while most are approximately 150 amino acids long. The analysis classified these genes into two subfamilies: the Classic subfamily (10 genes) and the Minus-C subfamily (two genes, namely ThawOBP3 and ThawOBP8) (Figure 2). All encoded proteins possess a putative N-terminal signal peptide, with the cleavage site predicted between amino acids 17 and 28. These putative ThawOBPs share from 31.4% (ThawOBP2) to 86.8% (ThawOBP5) sequence identity with their closest matches in the database, supported by highly significant E-values ranging from 8.28 × 10^−128^ to 3.67 × 10^−8^ (Table 2). Chromosomal mapping of the 12 identified ThawOBP genes across major genomic scaffolds was conducted. The genes are unevenly distributed, with sup_sca_14 harboring a significant number of ThawOBP loci. Several additional scaffolds also contain ThawOBP genes (Figure 1), suggesting a dispersed genomic organization of the odorant-binding protein family in this species.
3.2. Analysis of Phylogenetic Relationship and Gene Structure of OBPs in T. hawaiiensis
Investigation of the phylogenetic relationships and exon–intron structures of OBP genes in T. hawaiiensis unveiled considerable diversity within this gene family. A phylogenetic tree was constructed from full-length OBP sequences. The tree reveals the evolutionary relationships among the 12 ThawOBP genes, which are categorized into three distinct clades (Figure 3A). ThawOBP11 and ThawOBP12 form a closely related pair with high bootstrap support, suggesting a strong functional constraint. Similarly, ThawOBP4 and ThawOBP10 cluster together, indicating a close evolutionary relationship. ThawOBP1 and ThawOBP7 are positioned more distantly from the other groups, with ThawOBP1 and ThawOBP7 appearing as two of the more divergent sequences in the dataset (Figure 3A). The number of exons in these OBP genes varied between five and nine, with the majority containing six or seven (Figure 3B). Our analysis of the ThawOBP family showed that the average exon length was 69.97 bp. Furthermore, we found that the exon lengths across all 12 genes were relatively constrained, ranging from 57.9 to 82.8 bp (Figure 4A). Among the 12 ThawOBP genes, ThawOBP10 possessed the shortest average exon length (57.9 bp) and contained 7 exons. In contrast, ThawOBP3 had the highest exon count (9) and an average exon length of 78.0 bp. Furthermore, ThawOBP4, ThawOBP11, and ThawOBP12 shared an identical exon number of 6 and exhibited similar average exon lengths (Figure 4B). These structural features are entirely consistent with the clustering pattern observed in Figure 2.
3.3. Phylogenetic Relationship Analysis of All OBPs
Using the same identification methodology, we conducted analyses in species including Dendrothrips minowai [34], Frankliniella occidentalis [36], Frankliniella intonsa [40], Frankliniella fusca [35], Odontothrips loti [37], Thrips palmi [38], Stenchaetothrips biformis [41], Thrips tabaci [39], Megalurothrips usitatus (Hainan) [32], Megalurothrips usitatus (Zhejiang) [33], and Acyrthosiphon pisum [49]. The results revealed that the number of OBPs identified ranges from 10 to 17 across these species (Figure 5A). To assess the phylogenetic relevance between Thrips hawaiiensis OBPs and other OBPs, all OBPs were aligned to generate unrooted trees. As shown in Figure 5B, the OBPs from the same species (Zhejiang and Hainan populations) were tightly clustered, whereas those from other thrips species were interspersed, a pattern likely attributable to their close phylogenetic relationship within the same family, Thripidae.
3.4. Expression Patterns of T. hawaiiensis OBP Genes
The transcriptomic analysis was conducted using a comprehensive reference-based pipeline. Raw sequencing reads were quality-trimmed and adapter-removed using Trimmomatic (v0.39) [50] to ensure data quality for downstream analyses. Subsequently, the processed reads were aligned to the reference genome using HISAT2 (v2.2.1) [51], a splice-aware aligner optimized for RNA-seq data. Finally, gene-level read counts were quantified from the aligned reads using HTSeq-count (v2.0.2) [52] with default union-counting mode, generating a count matrix for subsequent differential expression analysis. To validate the gene expression profiles obtained from transcriptome sequencing, we selected 12 OBP genes in Thrips hawaiiensis for confirmation by RT-qPCR. The results demonstrated that 10 of these genes exhibited male-biased expression (Figure 6A), consistent with transcriptomic predictions. A strong correlation (Pearson correlation coefficient r = 0.83) was observed between the RNA-Seq and RT-qPCR results, validating the reliability of the transcriptomic data (Figure 6B). The male-biased expression patterns were consistent across both methods for most genes, except for ThawOBP11 and ThawOBP12. For these two genes, a discrepancy was noted: they showed minimal expression bias by RT-qPCR (Log2FC ≈ 0 or negative) but were indicated as low-level male-biased by RNA-Seq, a divergence potentially due to their low expression levels or technical limitations.
To investigate whether other olfaction-related gene families follow similar patterns, we also identified genes encoding chemosensory proteins (CSPs) (PF03392.9) using the same pipeline. A total of 11 CSP genes were identified in the T. hawaiiensis genome. The expression patterns of the 11 identified CSP genes were analyzed based on transcriptome sequencing and validated by RT-PCR. Transcriptomic data revealed that eight genes were significantly upregulated in males compared to females of T. hawaiiensis (Figure S1A). The RT-qPCR validation confirmed the accuracy of the transcriptomic expression profiles (Figure S1B). All primers used in this study are listed in Table S1. The results demonstrate that over half of the genes show significantly higher expression in males, consistent with the expression pattern observed for OBP genes.
4. Discussion
The identification of olfactory genes is fundamental to elucidating the molecular mechanisms of olfaction. In many insect species, odorant-binding proteins (OBPs) have been identified through transcriptomic and genomic analyses. Notably, studies have reported 51 OBP genes in Drosophila melanogaster [53], 65 in Anopheles gambiae [54], 64 in Aedes aegypti [54], 53 in Culex quinquefasciatus [54], 44 in Bombyx mori [55], and 50 in Tribolium castaneum [56]. In the present study, by analyzing our previously published genome data, we expanded the repertoire of OBPs in the insect species Thrips hawaiiensis to 12. While the 12 OBPs in T. hawaiiensis are substantially fewer than those in model insects such as D. melanogaster, this number is comparable to other thrips species, such as M. usitatus (14,17), F. occidentalis (10), T. palmi (15), F. intonsa (15), and O. loti (13) (Figure 5A). The number of OBP genes identified in other insects is significantly larger than that in thrips. This comparative reduction in thrips could be due to a simplified olfactory system or a unique evolutionary trajectory. Furthermore, phylogenetic analysis revealed that ThawOBP11 and ThawOBP12 cluster into a clade and exhibit the highest degree of sequence similarity (Figure 3A), a finding supported by sequence alignment (Figure 2).
For adult thrips, the most critical behaviors are host plant seeking, mating, and reproduction. These behaviors involve the extensive detection of both plant volatiles and thrips pheromones [57,58]. Male adults of both thrips species synthesize an aggregation pheromone detectable by both sexes that mediates attraction [59,60]. Consequently, these genes likely play a role in mediating the detection of both plant volatiles and the male-derived aggregation pheromone in Thrips hawaiiensis. Extensive research has demonstrated that odorant exposure can alter the expression levels of OBPs [61,62,63]. Through in vitro and in vivo functional assays in Bactrocera dorsalis, OBP83g-2 was identified as a key OBP involved in γ-octalactone perception, which was further confirmed to play a significant role in γ-octalactone-mediated oviposition behavior [64]. In Hyphantria cunea, OBP2 plays a crucial role in guiding larvae toward food sources that contain adult sex pheromones [65]. While the knockdown of SfruOBP18 did not impair larval survival or development, our combined RNAi and bioassay approach uncovered its critical function in conferring tolerance to multiple insecticides. This suggests a novel and non-canonical role for SfruOBP18 in insecticide susceptibility in Spodoptera frugiperda [66]. In our study, gene expression pattern analysis shows strong male-specific expression of OBP and CSP genes in T. hawaiiensis (Figure 6A and Figure S1). This pattern, also reported for OBPs and CSPs in M. usitatus, Frankliniella occidentalis, and Frankliniella intonsa [26,30], further supports their functional role in male-driven behaviors such as mate location and foraging. In experiments with Rhynchophorus ferrugineus, females injected with OBP-dsRNA showed a significant decrease in the expression of both RferOBP3 and RferOBP1768, which led to impaired perception of the odorants trans-2,4-nonadienal and trans-2-nonenal [67]. Docking results suggested a role for FoccOBP4/FintOBP4, FoccOBP6/FintOBP6, and FoccCSP2/FintCSP2 in transporting the major pheromone neryl (S)-2-methylbutanoate, with FoccOBP6/FintOBP6, FoccCSP2/FintCSP2, and FoccCSP3/FintCSP3 also implicated in binding the minor component (R)-lavandulyl acetate [30]. Based on these findings, we hypothesize that the majority of OBP and CSP genes in T. hawaiiensis are expressed at higher levels in males than in females, suggesting that males may possess a heightened sensitivity to environmental odors.
5. Conclusions
In summary, we report the first genome-wide identification of odorant-binding protein (OBP) genes and chemosensory protein (CSP) genes in Thrips hawaiiensis and characterize their expression profiles in both female and male T. hawaiiensis. This work provides crucial data for elucidating the functions of olfactory proteins in T. hawaiiensis, thereby paving the way for novel, targeted management strategies against thrips pests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Murai T. Development and reproductive capacity of Thrips hawaiiensis (Thysanoptera: Thripidae) and its potential as a major pest Bull. Entomol. Res.20019119319810.1079/BER 20018611415473 · doi ↗ · pubmed ↗
- 2Reynaud P. Balmès V. Pizzol J. Thrips hawaiiensis (Morgan, 1913) (Thysanoptera: Thripidae), an Asian pest thrips now established in Europe EPPO Bull.20083815516010.1111/j.1365-2338.2008.01201.x · doi ↗
- 3Aliakbarpour H. Che Salmah M.R. Seasonal abundance and spatial distribution of larval and adult thrips (Thysanoptera) on weed host plants in mango orchards in Penang, Malaysia Appl. Entomol. Zool.20114618519410.1007/s 13355-011-0030-5 · doi ↗
- 4Goldarazena A. First record of Thrips hawaiiensis (Morgan, 1913) (Thysanoptera: Thripidae), an Asian pest thrips in Spain EPPO Bull.20114117017310.1111/j.1365-2338.2011.02450.x · doi ↗
- 5Mound L.A. Thysanoptera: Diversity and interactions Annu. Rev. Entomol.20055024726910.1146/annurev.ento.49.061802.12331815355240 · doi ↗ · pubmed ↗
- 6Fu B. Qiu H. Li Q. Tang L. Zeng D. Liu K. Gao Y. Flower injection of imidacloprid and spirotetramat: A novel tool for the management of banana thrips Thrips hawaiiensis J. Pest Sci.2020931073108410.1007/s 10340-020-01209-7 · doi ↗
- 7Zhang F. Fu B. Liu K. Qiu H. Wu Y. The effect of temperature on the development and survival of Thrips hawaiiensis (Morgan)Acta Ecol. Sin.20143438953899
- 8Kurosawa M. Takaoka I. Naito T. Thrips infesting the tobacco plants in Japan (Thysanoptera)Jpn. J. Entomol.196432402