Retinoic Acid and Calcitriol Protect Mouse Primordial Follicles from Cyclophosphamide Treatment-Induced Apoptosis
Sihui He, Xiaodan Zhang, Wenjun Zhou, Ye Chen, Fengxin Liu, Weiyong Wang, Hongwei Wei, Yan Du, Meijia Zhang

TL;DR
Retinoic acid and calcitriol help protect mouse ovarian follicles from chemotherapy damage, preserving fertility without affecting cancer treatment.
Contribution
A new strategy using retinoic acid and calcitriol to protect ovarian follicles during chemotherapy is proposed.
Findings
RA and calcitriol reversed the decrease in primordial follicles caused by cyclophosphamide and doxorubicin.
Co-treatment preserved fertility in cyclophosphamide-treated mice without reducing antitumor efficacy.
RA and calcitriol reduced DNA damage and increased antioxidant proteins in oocytes.
Abstract
Chemotherapy causes primordial follicle apoptosis, resulting in premature ovarian insufficiency (POI) and infertility. In this study, we found that intraperitoneal injection of retinoic acid (RA) and calcitriol partially reversed the cyclophosphamide and doxorubicin treatment-induced decrease in primordial follicles in neonatal mouse ovaries. Furthermore, RA and calcitriol co-treatment reversed cyclophosphamide treatment-induced PI3K/Akt activity and FOXO3a nuclear export in the oocytes within primordial follicles, suggesting that the oocyte transcriptional activity was decreased, which in turn reduced the binding of chemotherapeutic drugs to DNA. Consistent with these findings, RA and calcitriol co-treatment reversed cyclophosphamide treatment-induced changes in reactive oxygen species (ROS), DNA damage response proteins (γH2AX, p-CHK2, p-p53, PUMA, BAX, Cleaved Caspase-3, and cPARP),…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
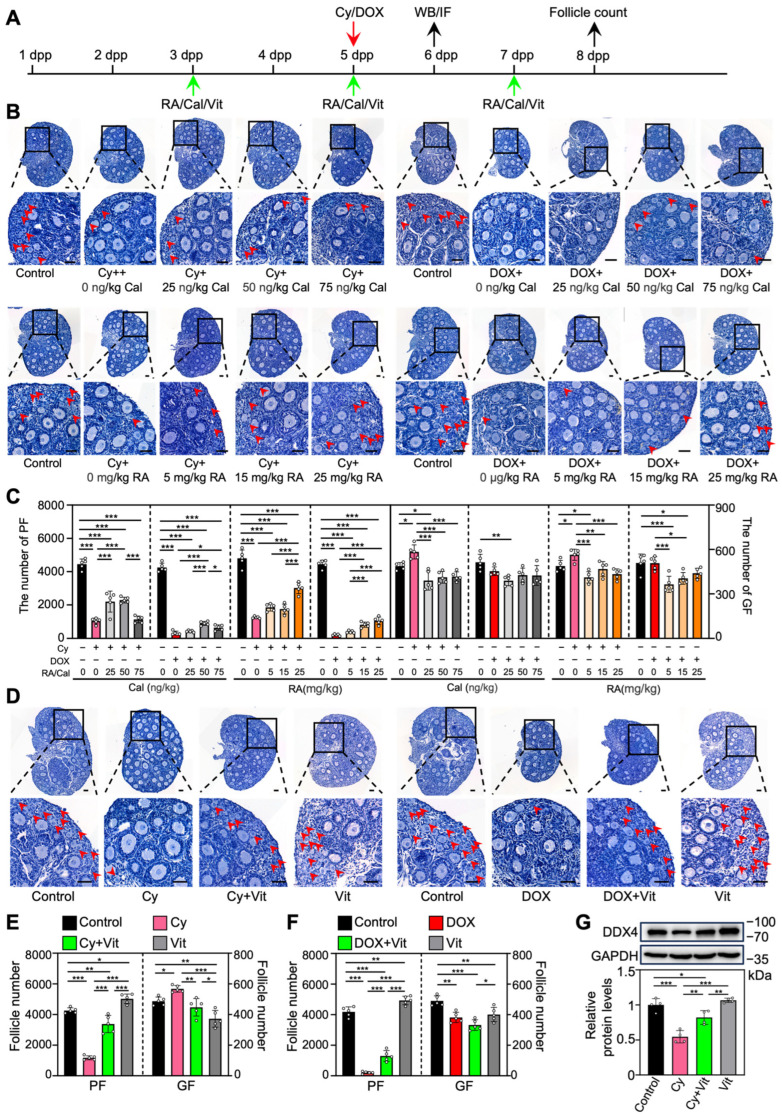 Figure 1
Figure 1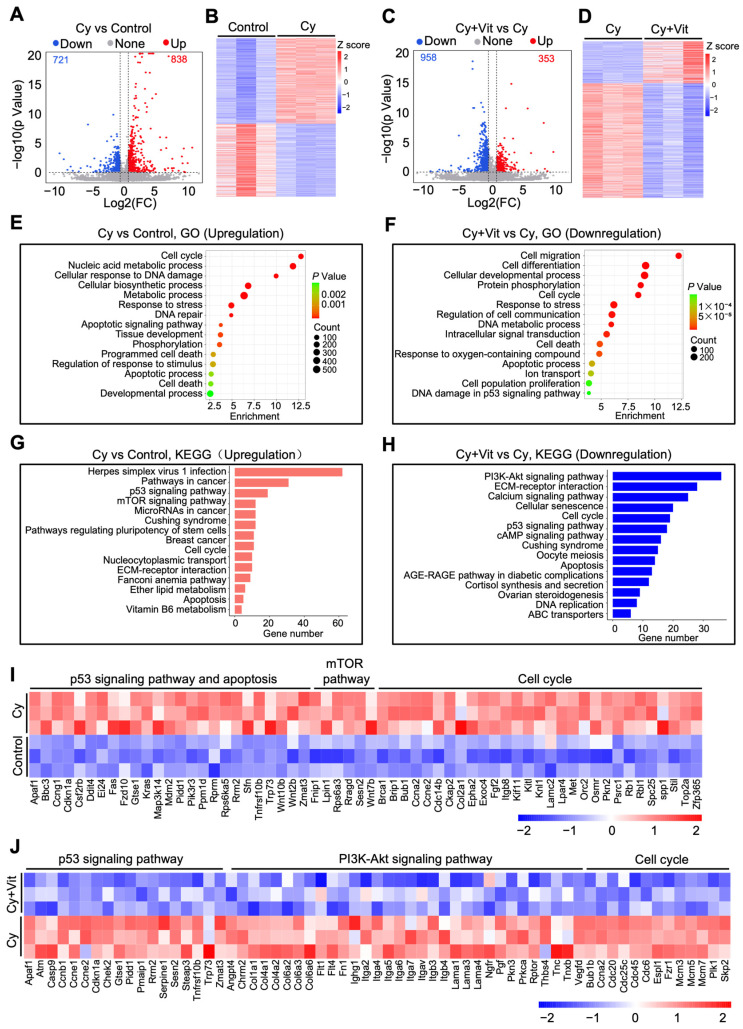 Figure 2
Figure 2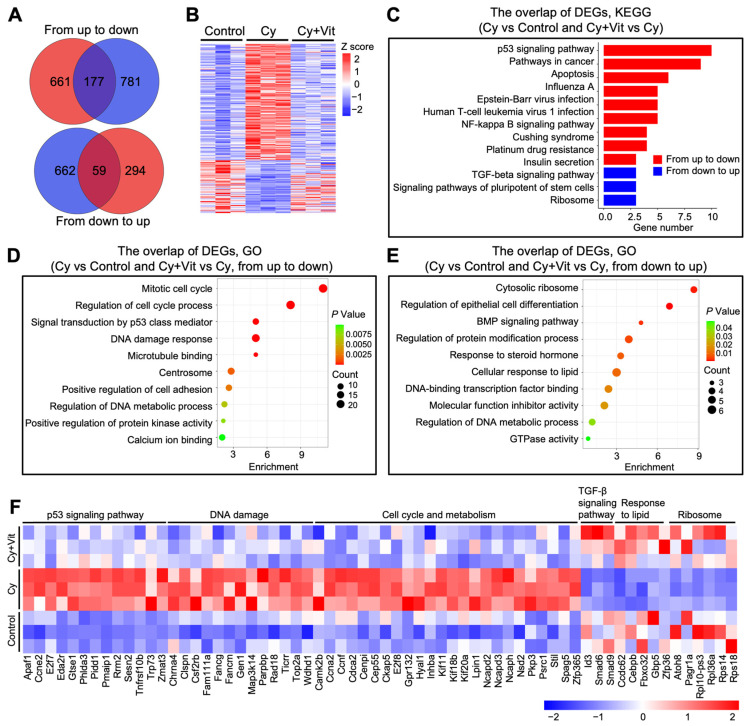 Figure 3
Figure 3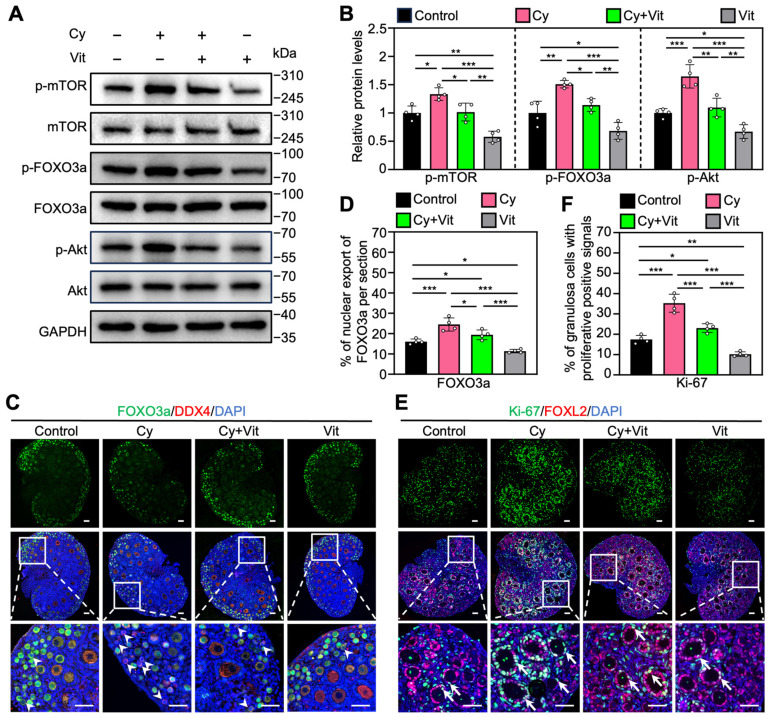 Figure 4
Figure 4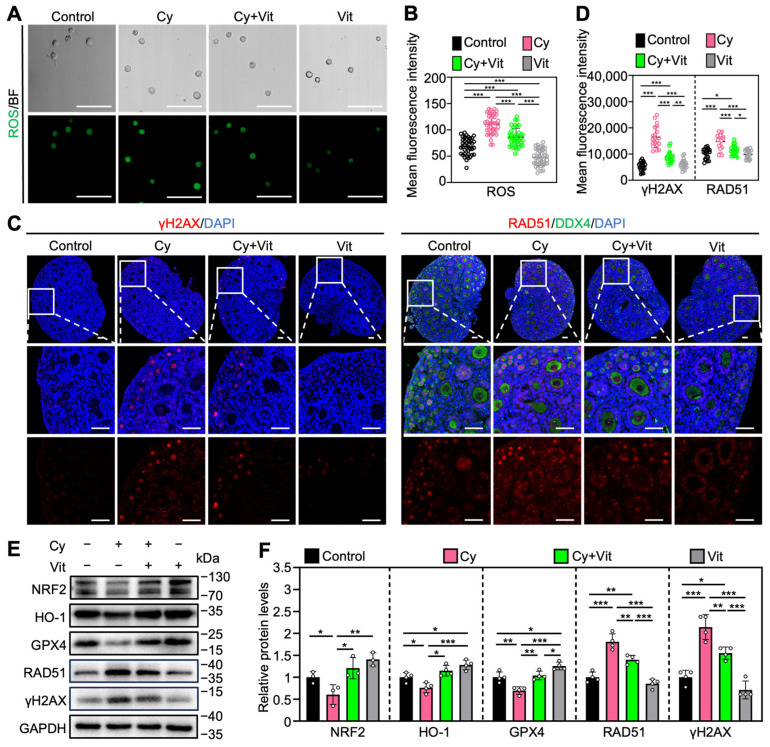 Figure 5
Figure 5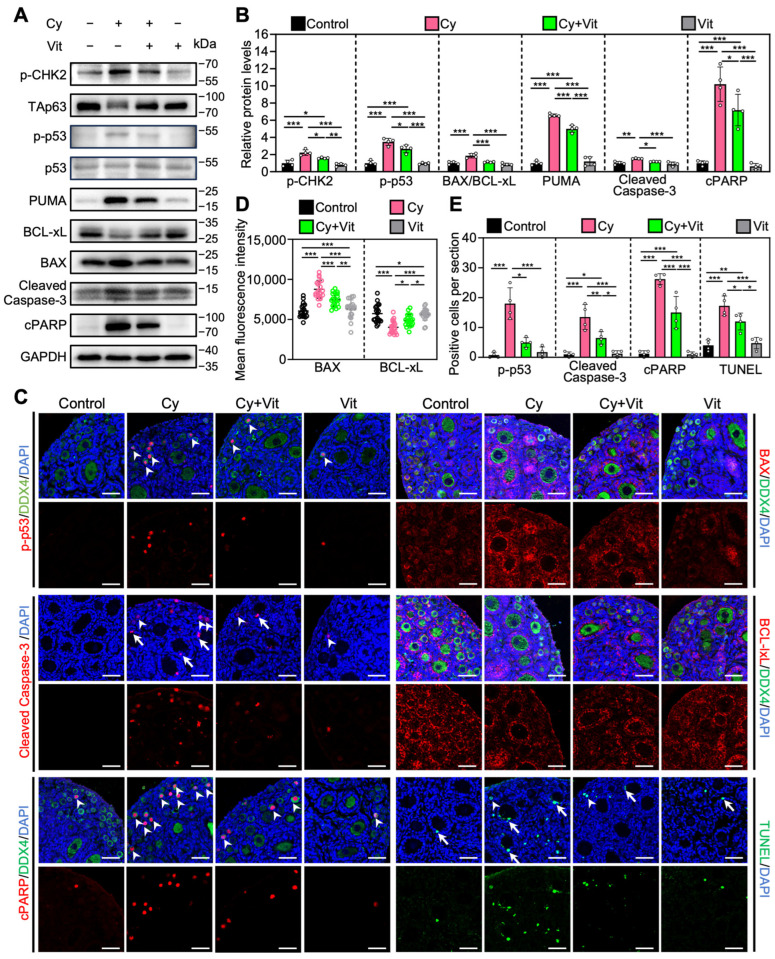 Figure 6
Figure 6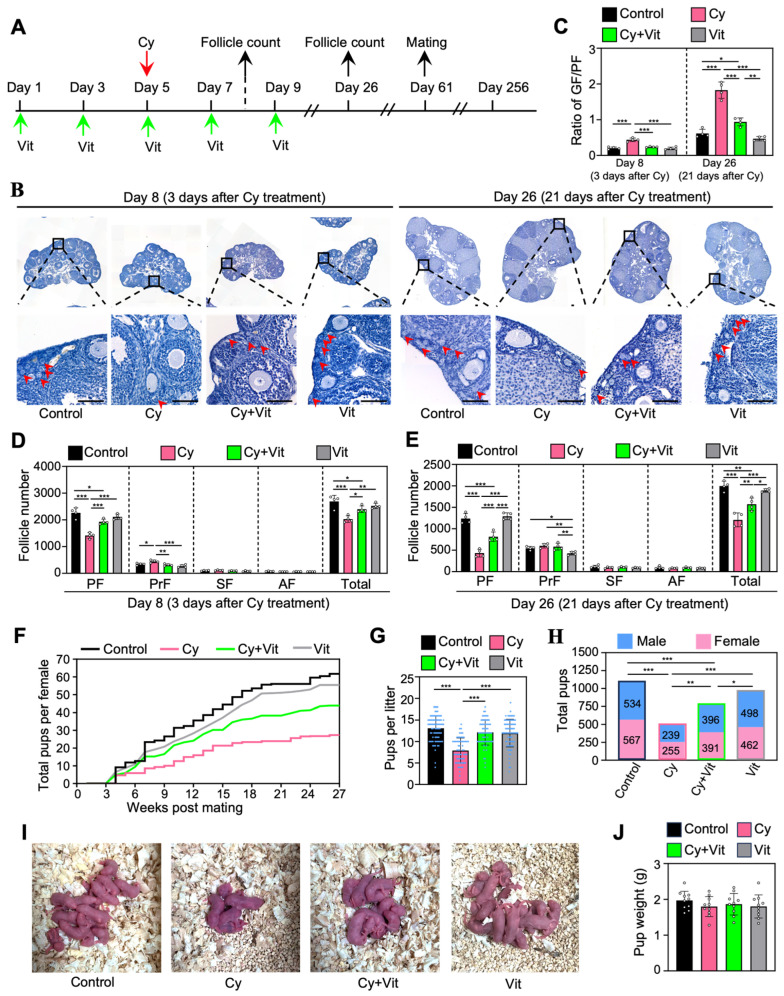 Figure 7
Figure 7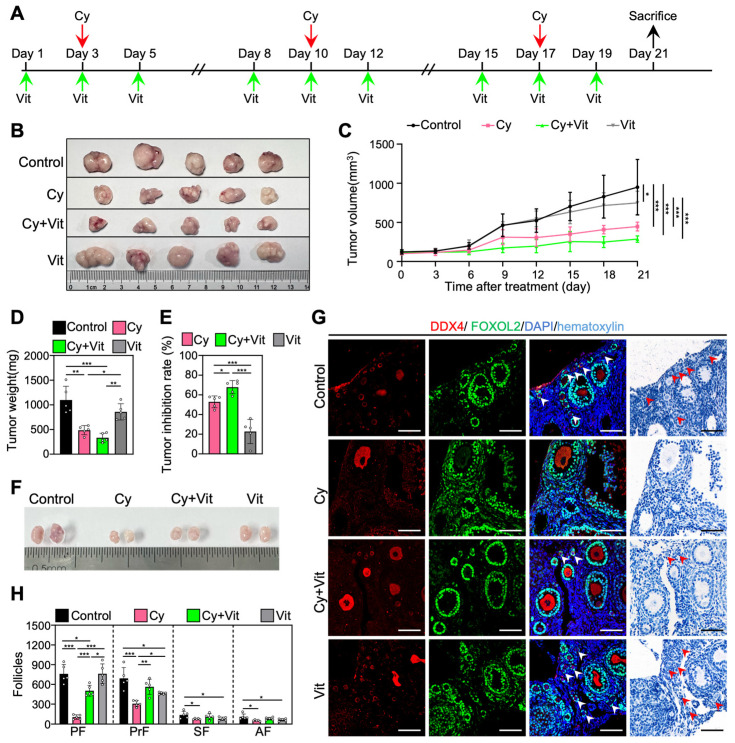 Figure 8
Figure 8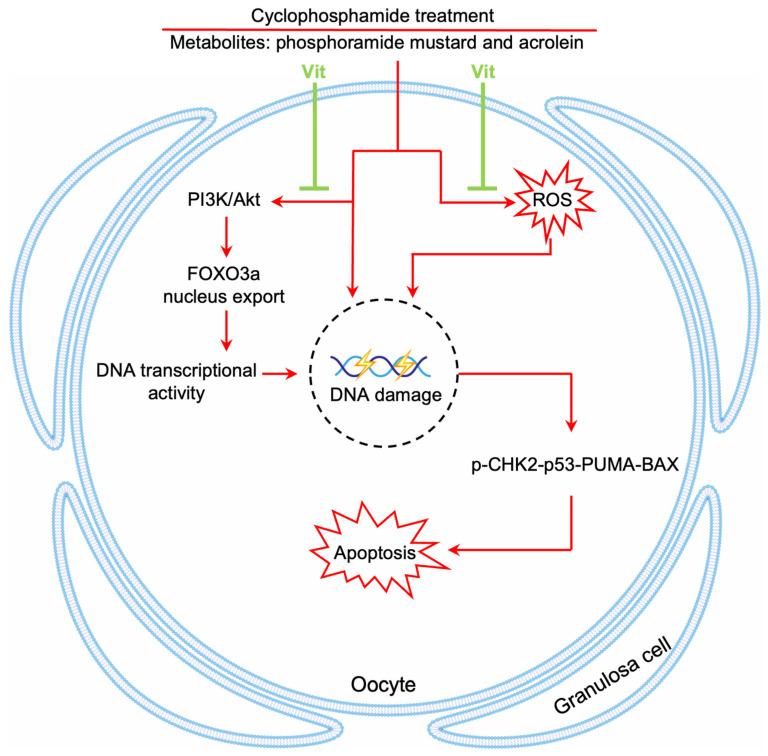 Figure 9
Figure 9- —National Key Research and Development Program of China
- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReproductive Biology and Fertility · Ovarian cancer diagnosis and treatment · Cancer Risks and Factors
1. Introduction
Cancer incidence is rising annually, with an observable trend toward younger ages [1]. This shift is driven by various factors [2], such as environmental pollution [3], drug residues [4], sleep deprivation [5], smoking prevalence [6], and unhealthy diets [7]. Chemotherapeutic drugs are widely used to improve the survival rate of patients with various cancers [8]. Both cyclophosphamide and doxorubicin are widely used to treat many malignant tumors such as lymphoma, leukemia, and breast cancer [9,10,11]. Cyclophosphamide is metabolized by hepatic microsomal cytochrome P450 to its active metabolites, phosphoramide mustard and acrolein [12]. Phosphoramide mustard alkylates DNA at the N-7 position of guanine, resulting in intra- and inter-strand DNA cross-links that prevent DNA replication and tumor cell division [13,14]. Acrolein causes a rapid reduction in intracellular glutathione (GSH) and overall antioxidant capacity, leading to excessive production of reactive oxygen species (ROS) and oxidative stress [9]. Acrolein also disrupts mitochondrial integrity and function, resulting in pro-apoptotic protein release and ATP depletion [15]. Doxorubicin (DOX) damages cancer cells by binding to topoisomerase II (TOP2) to form a covalent complex with DNA (DOX–TOP2–DNA) [16]. This complex inhibits DNA replication and transcription, resulting in DNA double-strand breaks (DSBs) and tumor cell apoptosis [17].
In the past few decades, the overall 5-year survival rates of all cancers combined have increased substantially, exceeding 90% for childhood acute lymphocytic leukemia and early-stage breast cancer [18]. However, most chemotherapeutic drugs, including cyclophosphamide and doxorubicin, cause massive apoptosis of primordial follicles, leading to premature ovarian insufficiency (POI) and female infertility [19,20,21]. Cancer survivors strongly seek to restore their ovarian function to maintain endocrine homeostasis and fertility [22]. Current fertility preservation strategies include oocyte and embryo cryopreservation for adult cancer patients [23] and ovarian tissue cryopreservation for prepubescent cancer patients before chemotherapy [24]. Subsequent assisted reproductive technologies are still needed to achieve fertility, including in vitro activation (IVA), in vitro fertilization (IVF), and/or embryo transfer [19,25]. All of these processes limit the final success rate. Moreover, the surgical removal of normal ovarian tissues from cancer patients results in a significant psychological burden, which will induce immunosenescence that decreases antitumor immunity [26,27]. Therefore, it is necessary to develop non-invasive strategies to protect ovarian function in situ during chemotherapy.
The follicles in the mammalian ovary are divided into two types: primordial and growing follicles. The primordial follicle pool, about 1–2 million oocytes in size, is established around the time of birth and is non-renewable [28]. In each wave, only a few primordial follicles develop into growing follicles, and they are ultimately depleted through ovulation or atresia [29]. The remaining primordial follicles are in a dormant state to maintain female reproductive lifespan [30]. Thus, the essence of protecting ovarian function is to protect the primordial follicles.
Retinoic acid (RA), also known as all-trans retinoic acid (ATRA), is the main active form of vitamin A [31]. RA can promote oocyte maturation [32,33], fertilization [34], and early embryonic development [35] in various mammalian species. RA works through the heterodimeric complexes of retinoic acid receptors (RARs) and retinoid X receptors (RXRs) [36]. RARs, including RARα, RARβ, and RARγ, are expressed in both human and mouse ovaries [37,38,39]. RXR can also form heterodimeric complexes with the vitamin D receptor (VDR) [40]. VDR is activated by calcitriol (1,25-dihydroxy vitamin D3), the active form of vitamin D [41]. Calcitriol/VDR is also crucial for follicular development [42]. Both RA and calcitriol have antioxidant, anti-inflammatory, and antitumor properties [42,43,44]. Therefore, we hypothesize that RA and calcitriol can maintain the quiescence of primordial follicle oocytes to reduce cyclophosphamide treatment-induced primordial follicle apoptosis, thereby preserving the primordial follicle reserve.
In the present study, RA and calcitriol co-treatment partially reversed cyclophosphamide treatment-induced primordial follicle apoptosis and fertility decrease in mice by inhibiting FOXO3a nuclear export and transcriptional activity in primordial follicle oocytes. Furthermore, RA and calcitriol preserved ovarian reserve without impairing cyclophosphamide’s antitumor efficacy in MCF-7 tumor-bearing mice. Therefore, our results suggest a promising non-invasive strategy for preserving ovarian reserve during chemotherapy in female cancer patients.
2. Methods
2.1. Animals and Chemicals
3- and 8-week-old ICR mice were purchased from the Guangdong Medical Laboratory Animal Center (Guangzhou, China), and 6–8-week-old female BALB/c-Nude mice were provided by GemPharmatech Co., Ltd. (Nanjing, China). All mice were housed in the animal facility of South China University of Technology under standardized conditions, with 20–24 °C, 50–70% relative humidity, and a 12-h/12 h light/dark cycle. The neonatal mice were obtained from the breeding of adult ICR mice, and the day of birth was designated as 0.5 days postpartum (dpp). Unless otherwise stated, all reagents were provided by Sigma-Aldrich (St. Louis, MO, USA).
2.2. Cell Culture
MCF-7 human breast cancer cells were purchased from Pricella Biotechnology Co., Ltd. (Wuhan, China), and cultured in RPMI-1640 medium (Gibco, Grand Island, NY, USA) containing 10% heat-inactivated fetal bovine serum (FBS, Thermo Fisher Scientific, Waltham, MA, USA) and 1% penicillin–streptomycin within an incubator at 37 °C with 5% CO_2_. Cells were passaged when they reached 90% confluence and were collected with 0.25% trypsin (Thermo Fisher Scientific) for experiments when they were in a logarithmic growth phase [45].
2.3. Animal Model and Treatments
Neonatal mice were intraperitoneally injected with cyclophosphamide (75 mg/kg), doxorubicin (10 mg/kg) or an equal volume of PBS on 5 dpp. At the same time, RA (0, 5, 15, 25 mg/kg), calcitriol (Cal; 0, 25, 50, 75 ng/kg), 25 mg/kg RA + 50 ng/kg calcitriol (Vit) or an equal volume of DMSO was intraperitoneally injected into the mice on 3, 5, and 7 dpp. Ovaries were collected from mice on 6 dpp for Western blotting and immunofluorescence staining, and were collected from mice on 8 dpp for follicle counting [46,47].
Adolescent mice (mice on 21 dpp as day 1) were intraperitoneally injected with cyclophosphamide (75 mg/kg) or an equal volume of PBS on day 5, and with Vit (25 mg/kg RA + 50 ng/kg calcitriol) or an equal volume of DMSO on days 1, 3, 5, 7, and 9. Six mice per group were sacrificed on days 8 and 26, and the ovaries were collected for follicle counting. The remaining females (n = 18/group) were mated with fertility-proven males from day 61 to day 256 [48,49].
The 6–8-week-old female BALB/c-Nude mice were subcutaneously injected with 1.4 × 10^7^ MCF-7 cells in 0.1 mL PBS into the left mammary fat pad. The drug treatment was started (designated as day 1) when the tumor volume of each mouse reached approximately 100 mm^3^. MCF-7 tumor-bearing mice were randomly divided into four groups: control, cyclophosphamide, cyclophosphamide + Vit, and Vit. MCF-7 tumor-bearing mice were intraperitoneally injected with cyclophosphamide (75 mg/kg) or an equal volume of PBS on day 3, 10, and 17, and with Vit (25 mg/kg RA + 50 ng/kg calcitriol) or an equal volume of DMSO on days 1, 3, 5, 8, 10, 12, 15, 17, and 19.
The tumor sizes were measured every 3 days, and the tumor volume was determined using the formula tumor volume (mm^3^) = LW^2^/2, where L represents the length, and W indicates the width. On day 21, BALB/c mice were euthanized by neck dislocation. Tumors and ovaries were immediately removed for tumor inhibition analysis and follicle counting, respectively [45]. The tumor growth inhibition rate was assessed according to the following formula: inhibition rate (%) = (1 − tumor volume of treatment group/tumor volume of control group) × 100%.
2.4. Histological Analysis and Follicle Counting
After paraformaldehyde fixing, graded ethanol dehydrating, and paraffin embedding, ovarian samples were sectioned serially at 5 µm as reported before [30], and then hematoxylin (Solarbio, Beijing, China) was used to stain these sections. To assess the total quantity of primordial follicles per ovary, primordial follicles were counted in every fifth section, with the total number calculated by multiplying the count by a correction factor of 5. Primary, secondary, and antral follicles (growing follicles) were counted in consecutive sections. Only non-overlapping follicles containing visible oocyte nuclei were counted to avoid double-counting. All sections were assessed by two independent individuals blinded to all groups.
2.5. Immunofluorescence
Immunofluorescence staining was performed following the protocol described in the previous study [29]. Briefly, after dewaxing and rehydrating, the ovarian sections underwent antigen retrieval with sodium citrate buffer, blockade with donkey serum, and then incubation with primary antibodies (Supplementary Table S1) at 4 °C overnight. The sections were then treated with secondary antibodies conjugated to Alexa Fluor 488 or 555 (Thermo Fisher Scientific) for 1 h at 37 °C. Finally, the sections were stained with 4′,6-diamidino-2-phenylindole (DAPI) for 5 min, and then treated with anti-fluorescence quenching mounting medium (Ruitaibio, Beijing, China). All sections were imaged using an LSM 800 confocal microscope (Carl Zeiss, Oberkochen, Germany) under the same imaging parameters. ZEN software (Carl Zeiss, Version 3.1) was used to analyze and quantify fluorescence intensity. The mean value of the five largest sections in each ovary was considered one independent replicate.
2.6. Isolation of Oocytes from Neonatal Mice
The collected ovaries were digested with 0.25% trypsin (Thermo Fisher Scientific) for 10 min at 37 °C and then were terminated via the addition of 10% FBS (Thermo Fisher Scientific) in pre-warmed M2 medium (Thermo Fisher Scientific). Oocytes were released from the follicles. Then, oocytes were collected from the cell suspension using mouth-operated glass pipettes under a stereomicroscope, washed thoroughly, and placed in pre-warmed M2 medium (Thermo Fisher Scientific) droplets for later use.
2.7. ROS Staining of Oocytes
The ROS levels in oocytes were detected following the ROS detection kit (Beyotime, Beijing, China) [50]. Briefly, oocytes were incubated in pre-warmed M2 medium (Thermo Fisher Scientific) with 10 μM 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA, Beyotime) for 30 min at 37 °C in the dark, and then washed three times in pre-warmed M2 medium. Subsequently, the oocytes were transferred to cell culture dishes (NEST, Beijing, China) and imaged under the same parameters using an LSM 800 confocal microscope (Carl Zeiss).
2.8. TUNEL Staining
Follicle apoptosis in the ovaries was detected by the Click-iT Plus TUNEL Assay (Thermo Fisher Scientific) [50]. Briefly, dewaxed and rehydrated ovarian sections were permeabilized with proteinase K for 30 min at room temperature. After two washes with PBS, the sections were incubated with the TUNEL reaction mixture for 1 h at 37 °C in the dark. Subsequently, nuclei were stained with DAPI (Beyotime) for visualization. All sections were imaged with an LSM 800 confocal microscope (Carl Zeiss) under the same parameters.
2.9. Western Blotting
6–8 ovaries per replicate were collected for protein extraction as reported before [29]. Briefly, the ovarian samples were lysed on ice, and proteins were collected to detect the concentration. Equal quantities of protein (20 μg) were combined with SDS loading buffer (Cwbio, Beijing, China) and denatured by heating at 95 °C for 10 min and loaded into a 5% stacking gel and a 10% separating gel for electrophoresis. After that, the proteins were transferred to polyvinylidene difluoride (PVDF) membranes. The membranes were then blocked with 5% skim milk, followed by an overnight incubation with primary antibodies (Supplementary Table S1) at 4 °C and a 1 h incubation with anti-mouse or anti-rabbit IgG (1:5000, ZSGB-BIO, Beijing, China) at room temperature. Protein bands were detected using a chemiluminescent substrate (NCM Biotech, Suzhou, China) and captured with a chemiluminescence imaging system (Tanon, Shanghai, China). All protein band densities were measured using ImageJ software (version 1.4.3.67; NIH Image, Bethesda, MD, USA), using GAPDH as the internal control.
2.10. RNA-Sequencing
Neonatal mice were intraperitoneally injected with cyclophosphamide (75 mg/kg), doxorubicin (10 mg/kg), or an equal volume of PBS on 5 dpp. At the same time, RA (5, 15, 25 mg/kg), calcitriol (Cal; 25, 50, 75 ng/kg), 25 mg/kg RA + 50 ng/kg calcitriol (Vit), or an equal volume of DMSO was intraperitoneally injected into the mice on 3 and 5 dpp. Ovaries were collected from mice on 6 dpp, and preserved in sample buffer before being submitted for RNA extraction and subsequent RNA sequencing at Kidio Biotechnology Co., Ltd. (Guangzhou, China). The analysis was performed using Metascape (https://metascape.org) and Omicstudio (https://www.omicstudio.cn).
2.11. Statistical Analysis
All experiments were performed with at least three independent replicates. Data are displayed as the mean ± standard deviation (SD) and analyzed by one-way ANOVA with Tukey’s multiple-comparison test or a two-tailed unpaired Student’s t-test. Statistical analyses were performed using the GraphPad Prism software (version 8.0.1, La Jolla, CA, USA).
3. Results
3.1. RA and Calcitriol Reduce Cyclophosphamide and Doxorubicin Treatment-Induced Primordial Follicle Loss in Mice
Analysis of scRNA-seq data (GSE263836) revealed that RARs were predominantly expressed in mouse oocytes of primordial follicles (Figure S1). The injection of RA or calcitriol in neonatal mice decreased the number of growing follicles in the ovaries (Figure S2), and the most effective doses were 25 mg/kg and 50 ng/kg for RA and calcitriol, respectively (Figure S2). This is consistent with our recent studies that both RA and calcitriol inhibit mouse primordial follicle activation. Consistent with previous studies [47,48], the injection of a single dose of 75 mg/kg cyclophosphamide or 10 mg/kg doxorubicin to neonatal mice resulted in the loss of a large number of primordial follicles, along with an increase in the number of growing follicles in the cyclophosphamide treatment group (Figure S3).
To investigate the effect of RA and calcitriol on cyclophosphamide or doxorubicin treatment-induced primordial follicle loss, we intraperitoneally injected neonatal mice with cyclophosphamide or doxorubicin, and with RA and calcitriol before and after chemotherapy drug treatment (Figure 1A). RA or calcitriol significantly reduced cyclophosphamide treatment-induced primordial follicle loss, with the most effective doses of 25 mg/kg RA and 50 ng/kg calcitriol (Figure 1B,C). The co-treatment of RA and calcitriol (definition as Vit) further reduced cyclophosphamide treatment-induced primordial follicle loss (Figure 1D,E). A relatively weak protective effect of Vit was observed in the doxorubicin-treated group (Figure 1B–F).
We also examined DDX4 (DEAD-box helicase 4), a germ cell marker protein. Compared with the control, cyclophosphamide treatment resulted in a significant decrease in DDX4 protein levels, likely due to the reduction in primordial follicle numbers (Figure 1G and Figure S4). This was partially reversed by Vit treatment (Figure 1G). Consistent with the results of RA or calcitriol (Figures S2 and S5), Vit treatment significantly decreased the number of growing follicles compared with control (Figure 1D–F). Thus, the subsequent experiments focused on the mechanism of Vit in preventing cyclophosphamide treatment-induced mouse primordial follicle loss.
3.2. Transcriptomic Analysis Reveals Regulatory Pathways Involved in Vit Effects on the Ovaries from Cyclophosphamide-Treated Neonatal Mice
We further studied the effects of Vit on transcriptomic changes in the ovaries of cyclophosphamide-treated neonatal mice. The volcano plot and heatmap analyses revealed that 1559 differentially expressed genes (DEGs, 838 upregulation and 721 downregulation) were present in the ovaries of the cyclophosphamide treatment group compared with the control (Figure 2A,B). The upregulated DEGs were mainly related to the p53 pathway, apoptosis, DNA damage and repair, the mTOR pathway, oxidative phosphorylation, cell proliferation, and tissue development (Figure 2E,G,I), and the downregulated DEGs were mainly related to ribosome metabolism, ATP production, and the mitochondrial electron transport chain (Figure S6A,B,E). Compared with the cyclophosphamide treatment group, the co-treatment of cyclophosphamide and Vit resulted in 1311 DEGs (353 upregulation and 958 downregulation, Figure 2C,D). The upregulated DEGs were associated with the TGF-β signaling pathway, the TNF signaling pathway, and immune response processes (Figure S6C,D), while the downregulated DEGs were mainly associated with the PI3K/Akt signaling pathway, the p53 signaling pathway, apoptosis, and cell cycle regulation (Figure 2F,H,J).
Further Venn diagram and heatmap analysis showed the overlap of these DEGs (Figure 3A,B). Cyclophosphamide treatment upregulated 838 DEGs, among which 177 DEGs were downregulated by Vit (from up to down). These 177 DEGs were mainly enriched in the p53 signaling pathway, DNA damage response, and apoptotic regulation pathways (Figure 3C,D,F). By contrast, cyclophosphamide treatment downregulated 721 DEGs, among which 59 DEGs were upregulated by Vit (from down to up). These 59 DEGs were mainly enriched in the TGF-β signaling pathway, response to lipid, and ribosome (Figure 3C,E,F). Based on the above observations, we hypothesize that Vit reduces cyclophosphamide treatment-induced primordial follicle loss by inhibiting DNA damage and apoptosis.
3.3. Vit Partially Reverses Cyclophosphamide Treatment-Induced Primordial Follicle Oocyte Transcriptional Activity
Consistent with previous studies [48,51,52], cyclophosphamide treatment increased the protein levels of p-mTOR, p-Akt, and p-FOXO3a (Figure 4A,B and Figure S7A), as well as the proportions of primordial follicle oocytes with FOXO3a nuclear export (Figure 4C,D) and of granulosa cells with Ki-67-positive signals (Figure 4E,F) compared with control. However, all of these cyclophosphamide treatment-induced increases were partially reversed by Vit (Figure 4). Vit treatment alone also reduced these phosphorylated protein levels, and the proportions of oocytes with FOXO3a nuclear export and granulosa cells with Ki-67 positive signaling compared with control (Figure 4). Therefore, Vit inhibited cyclophosphamide treatment-induced PI3K/Akt activation and FOXO3a nuclear export in primordial follicle oocytes, suggesting that Vit partially reverses cyclophosphamide treatment-induced primordial follicle oocyte transcriptional activity.
3.4. Vit Partially Reverses Cyclophosphamide Treatment-Induced Oxidative Stress and DNA Damage
Consistent with previous studies [20,53,54,55,56,57,58], cyclophosphamide treatment increased ROS levels in the primordial follicle oocytes (Figure 5A,B) and the protein levels of γ-H2AX and RAD51 in the ovaries (Figure 5C–F and Figure S7B), while decreasing the protein levels of NRF2, HO-1, and GPX4 in the ovaries compared with control (Figure 5E,F). However, all of these cyclophosphamide treatment-induced changes were partially reversed by Vit (Figure 5). Vit treatment alone also reduced ROS levels in primordial follicle oocytes (Figure 5A,B) and increased the protein levels of HO-1 and GPX4 in the ovaries compared with control (Figure 5E,F). These findings indicate that Vit treatment enhances the antioxidant capacity and restores redox homeostasis in the neonatal mouse ovary. Therefore, Vit partially reverses cyclophosphamide treatment-induced oxidative stress and DNA damage.
3.5. Vit Partially Reverses Cyclophosphamide Treatment-Induced Primordial Follicle Apoptosis
Consistent with previous studies [54,59,60], cyclophosphamide treatment increased the levels of pro-apoptotic proteins (p-CHK2, p-p53, PUMA, BAX, Cleaved Caspase-3, and cPARP) and the number of cells with TUNEL-positive signals, while decreasing the levels of the anti-apoptotic protein BCLxL in the ovaries compared with control (Figure 6 and Figure S8). In addition, cyclophosphamide treatment induced TAp63 activation, as indicated by a mobility shift that was absent in the control group (Figure 6A). Immunofluorescence analysis showed that cyclophosphamide treatment increased p-p53 levels (Figure 6C–E) and decreased TAp63 levels (Figure S9A,B) in the oocyte nuclei of primordial follicles compared with the control. However, all of these cyclophosphamide treatment-induced changes were partially reversed by Vit (Figure 6). Therefore, Vit partially reverses cyclophosphamide treatment-induced primordial follicle apoptosis.
3.6. Vit Preserves Fertility in Cyclophosphamide-Treated Mice
To investigate the effect of Vit on the fertility in cyclophosphamide-treated mice, we intraperitoneally injected adolescent mice with cyclophosphamide (day 5), and/or with Vit (days 1, 3, 5, 7, and 9) before and after cyclophosphamide treatment (Figure 7A). Ovaries were collected for follicle counting on day 8 (3 days post-cyclophosphamide treatment) and day 26 (Figure 7A). The remaining female mice were mated with fertility-proven males for the fertility test on day 61 after the start of the experiment (Figure 7A).
Consistent with previous studies [52,61], a single injection of cyclophosphamide significantly decreased primordial follicle numbers on day 8 and further decreased the number on day 26 (Figure 7B–E), but had no obvious effect on the ovarian morphology compared with control (Figure S10). Consistent with this, cyclophosphamide treatment increased the ratio of growing/primordial follicles compared with control (Figure 7C). However, all of these cyclophosphamide treatment-induced changes were partially reversed by Vit (Figure 7B–E). Cyclophosphamide significantly increased the number of primary follicles on day 8 compared with control, but had no effect on the number of growing follicles on day 26 compared with control (Figure 7B–E), possibly because cyclophosphamide treatment-induced growing follicles failed to develop normally.
Consistent with previous studies [48,49,59], cyclophosphamide also significantly decreased the cumulative number of pups per mouse, the number of pups per litter, and the total number of pups compared with control (Figure 7F–H). However, Vit treatment partially reversed these cyclophosphamide-induced decreases in fertility. All groups of neonatal mice had no obvious malformations and no weight differences (Figure 7I,J). Therefore, Vit preserves fertility in cyclophosphamide-treated mice by preserving the reserve of primordial follicles.
3.7. Vit Preserves the Primordial Follicle Reserve in Cyclophosphamide-Treated MCF-7 Tumor-Bearing Mice
We established an MCF-7 tumor-bearing mouse model to investigate whether Vit can protect the primordial follicle pool during cyclophosphamide treatment. The 8-week-old BALB/c-Nude mice were subcutaneously injected with MCF-7 cells into the left mammary fat pad. When the tumor volume was close to 100 mm^3^ (day 1), MCF-7 tumor-bearing mice were injected intraperitoneally with cyclophosphamide three times (days 3, 10, and 17), and/or with Vit before and after each cyclophosphamide treatment (Figure 8A). All mice were euthanized on day 21 after the start of the experiment for ovary and tumor collection (Figure 8A).
Consistent with previous studies [62,63], three doses of cyclophosphamide injection significantly decreased the volume and weight of tumors (Figure 8B–E and Figure S11), the size of ovaries (Figure 8F), and the number of primordial, primary, secondary, and antral follicles (Figure 8G,H) compared with control. However, Vit partially reversed the cyclophosphamide-induced decrease in the number of primordial and growing follicles (Figure 8G,H). Vit treatment alone also demonstrated modest antitumor activity and further increased the tumor inhibition rate of cyclophosphamide treatment in MCF-7 tumor-bearing mice (Figure 8B–E), consistent with previous studies on MCF-7 cells [64,65]. Therefore, Vit preserves the primordial follicle reserve in cyclophosphamide-treated MCF-7 tumor-bearing mice.
4. Discussion
Chemotherapy causes a dramatic decline in the ovarian reserve of cancer patients, ultimately leading to POI and even infertility [1]. In the present study, RA and calcitriol reduced cyclophosphamide and doxorubicin treatment-induced primordial follicle depletion in neonatal mouse ovaries. Furthermore, RA and calcitriol co-treatment preserved fertility in cyclophosphamide-treated mice without impairing cyclophosphamide’s antitumor efficacy in MCF-7 tumor-bearing mice. Therefore, the mechanism of RA and calcitriol involves enhancing antioxidant capacity and inhibiting cyclophosphamide treatment-induced DNA transcriptional activity, leading to reduced binding of cyclophosphamide’s active metabolites to primordial follicle oocyte DNA (Figure 9).
Cyclophosphamide treatment produces the active metabolite phosphoramide mustard in the liver [66]. Phosphoramide mustard forms cross-links with DNA and interferes with DNA replication and transcription, ultimately leading to cell cycle arrest and apoptosis in rapidly dividing cancer cells [9]. Postnatal mammalian oocytes are arrested at the diplotene stage of meiosis prophase I but nevertheless execute widespread transcriptional changes, and the oocyte nucleus is very large with a loose chromatin structure [67,68,69]. Thus, oocytes are sensitive to chemotherapy drugs [70]. Although the oocytes within primordial follicles are in a relatively quiescent state, they are more sensitive to chemotherapy drugs due to the lack of both DNA repair ability and antioxidant capacity in the oocytes within growing follicles [71,72]. In the present study, the injection of cyclophosphamide or doxorubicin into neonatal mice led to the depletion of primordial follicles by upregulating the DNA damage response and the p53-dependent apoptotic pathway in oocytes. These results are consistent with previous studies [20,59].
Cyclophosphamide treatment decreased ovarian reserve mainly by inducing primordial follicle apoptosis in neonatal mice [73,74]. Cyclophosphamide treatment increases the phosphorylation levels of PI3K/Akt and FOXO3a in the ovaries of neonatal mice, as shown in our study and previous studies [48,51]. FOXO3a, a nuclear transcriptional repressor, maintains primordial follicle oocytes’ dormancy by inhibiting their transcriptional activity [75]. Cyclophosphamide treatment induced FOXO3a phosphorylation and then promoted FOXO3a translocation from the nucleus to the cytoplasm, resulting in an increase in the transcriptional activity in primordial follicle oocytes. The increased transcriptional activity in primordial follicle oocytes could further promote the binding of cyclophosphamide’s active metabolites to DNA, thereby promoting primordial follicle apoptosis. RA and calcitriol could inhibit PI3K/Akt activity and reduce FOXO3a phosphorylation levels and nuclear export in primordial follicle oocytes, resulting in decreased transcriptional activity in primordial follicle oocytes. Further studies from our laboratory indicated that both RA and calcitriol could interact with p85α by binding to their respective receptors to inhibit the PI3K/Akt signaling pathway (unpublished data). Thus, RA and calcitriol inhibit cyclophosphamide treatment-induced DNA transcriptional activity, possibly leading to a reduction in oxidative stress and primordial follicle apoptosis by decreasing the binding of cyclophosphamide’s active metabolites to primordial follicle oocyte DNA [9,76].
The metabolic products of cyclophosphamide, particularly acrolein, generate excessive ROS and impair the antioxidant defense system (GSH depletion), disrupting the redox homeostasis [77]. In the present study, cyclophosphamide treatment increased oxidative stress by increasing ROS levels in the oocytes of primordial follicles, consistent with previous studies in ovaries [78,79]. Cyclophosphamide treatment also increased oxidative stress by decreasing the protein levels of NRF2, HO-1, and GPX4 in neonatal mouse ovaries, consistent with previous studies in various tissues [80,81,82,83]. RA combined with chemotherapeutic agents is usually applied for the prevention and treatment of different types of cancers, particularly acute promyelocytic leukemia (APL) [84]. One of the mechanisms is that RA reduces NRF2 activity through RARα to increase the susceptibility to electrophiles and oxidative stressors in cancer cells [85]. However, RA and calcitriol could reduce ROS production and enhance antioxidant enzyme activities in bovine oocytes [34,86] and goat granulosa cells [42,87]. RA could reduce doxorubicin-induced cardiomyocyte apoptosis by inhibiting ROS generation and activating the antioxidant defense system [44,88]. Similarly, calcitriol has been shown to reduce hyperosmotic stress-induced cytotoxicity in human corneal epithelial cells by reducing intracellular ROS generation and activating NRF2-antioxidant signaling [89]. Consistent with the above studies, RA and calcitriol reversed the increase in ROS levels and decrease in antioxidant capacity induced by cyclophosphamide treatment, resulting in a reduction in primordial follicle apoptosis in neonatal mouse ovaries.
RA and calcitriol are used to treat leukemia [84] and breast cancer [90], respectively. We found that RA and calcitriol protected mouse primordial follicles from cyclophosphamide treatment-induced apoptosis. The toxic dose of RA in mice is 40–80 mg/kg [91], while the intraperitoneal injection of 4 μg/kg calcitriol in mice has no toxicity [92]. In the present study, 25 mg/kg RA and 50 ng/kg calcitriol were used to protect mouse primordial follicles from cyclophosphamide treatment-induced apoptosis. RA and calcitriol had no negative impact on mouse reproduction after observation for half a year (Figure 7). However, the efficacy and safety of RA and calcitriol require further clinical validation. Additionally, whether RA and calcitriol also protect against ovarian damage from other chemotherapeutics (e.g., cisplatin) requires further investigation.
In conclusion, our study indicates that RA and calcitriol partially reverse cyclophosphamide treatment-induced primordial follicle apoptosis by inhibiting cyclophosphamide treatment-induced transcriptional activity and enhancing antioxidant capacity (Figure 9). These findings suggest a potential non-invasive therapeutic approach for ovarian reserve protection during chemotherapy in female cancer patients.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Guo Y. Xue L. Tang W. Xiong J. Chen D. Dai Y. Wu C. Wei S. Dai J. Wu M. Ovarian microenvironment: Challenges and opportunities in protecting against chemotherapy-associated ovarian damage Hum. Reprod. Update 20243061464710.1093/humupd/dmae 02038942605 PMC 11369228 · doi ↗ · pubmed ↗
- 2Ugai T. Sasamoto N. Lee H.Y. Ando M. Song M. Tamimi R.M. Kawachi I. Campbell P.T. Giovannucci E.L. Weiderpass E. Is early-onset cancer an emerging global epidemic? Current evidence and future implications Nat. Rev. Clin. Oncol.20221965667310.1038/s 41571-022-00672-836068272 PMC 9509459 · doi ↗ · pubmed ↗
- 3Navarrete-Meneses M.D.P. Salas-Labadía C. Gómez-Chávez F. Pérez-Vera P. Environmental pollution and risk of childhood cancer: A scoping review of evidence from the last decade Int. J. Mol. Sci.202425328410.3390/ijms 2506328438542255 PMC 10970446 · doi ↗ · pubmed ↗
- 4Khalifa H.O. Shikoray L. Mohamed M.I. Habib I. Matsumoto T. Veterinary drug residues in the food chain as an emerging public health threat: Sources, analytical methods, health impacts, and preventive measures Foods 202413162910.3390/foods 1311162938890858 PMC 11172309 · doi ↗ · pubmed ↗
- 5Peng Z. Song J. Zhu W. Bao H. Hu Y. Shi Y. Cheng X. Jiang M. Fang F. Chen J. Impact of sleep deprivation on colon cancer: Unraveling the Kyn A-P 4HA 2-HIF-1α axis in tumor lipid metabolism and metastasis Mol. Metab.20259310210910.1016/j.molmet.2025.10210939920992 PMC 11869867 · doi ↗ · pubmed ↗
- 6Islami F. Nargis N. Liu Q. Bandi P. Siegel R.L. Choudhury P.P. Freedman N.D. Warner K.E. Jemal A. Averted lung cancer deaths due to reductions in cigarette smoking in the United States, 1970–2022 CA Cancer J. Clin.20257521622510.3322/caac.7000540131130 PMC 12061630 · doi ↗ · pubmed ↗
- 7Chang K. Gunter M.J. Rauber F. Levy R.B. Huybrechts I. Kliemann N. Millett C. Vamos E.P. Ultra-processed food consumption, cancer risk and cancer mortality: A large-scale prospective analysis within the UK Biobank E Clinical Medicine 20235610184010.1016/j.eclinm.2023.10184036880051 PMC 9985039 · doi ↗ · pubmed ↗
- 8Bertoldo M.J. Smitz J. Wu L.E. Lee H.C. Woodruff T.K. Gilchrist R.B. Prospects of rescuing young eggs for oncofertility Trends Endocrinol. Metab.20203170871110.1016/j.tem.2020.07.00432807599 · doi ↗ · pubmed ↗