TRPV1 from the TRP family: Structure, function, implication in autoimmune diseases and potential therapies
Typhaine Bejoma, Yanna Pan, Qingjie Zhao

TL;DR
This paper reviews the TRPV1 ion channel's structure, function, and role in autoimmune diseases, suggesting it as a potential target for new therapies.
Contribution
The paper provides a comprehensive overview of TRPV1's involvement in autoimmune diseases and its therapeutic potential.
Findings
TRPV1 regulates lipid synthesis and inflammatory responses in immune cells.
TRPV1 is implicated in autoimmune diseases like lupus, multiple sclerosis, and rheumatoid arthritis.
Selective agonists and antagonists for TRPV1 are being explored for therapeutic development.
Abstract
The transient receptor potential vanilloid type 1 (TRPV1) channel, a member of the TRP ion channel family, plays a crucial role in both physiological and pathological processes. This review provides an overview of the structure, biological functions, and implications of TRPV1 in autoimmune diseases. The structural characteristics of TRPV1, including its transmembrane and intracellular domains, are examined to understand its activation and modulation. In addition to its well-known role as a thermosensor in nociceptive neurons, TRPV1 has been found to have functions in immune cells where it regulates lipid synthesis and inflammatory response. The investigation of TRPV1’s involvement in autoimmune conditions such as systemic lupus erythematosus, multiple sclerosis, and rheumatoid arthritis highlights its potential as a therapeutic target. The search for selective agonists and antagonists…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
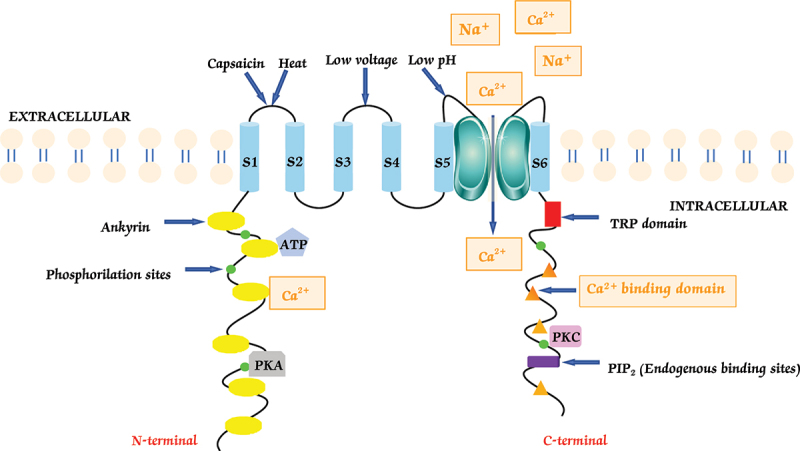 Figure 1
Figure 1 Figure 2
Figure 2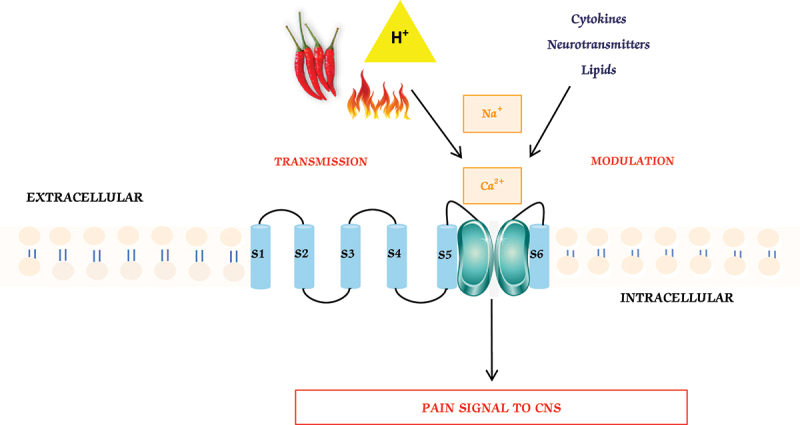 Figure 3
Figure 3 Figure 4
Figure 4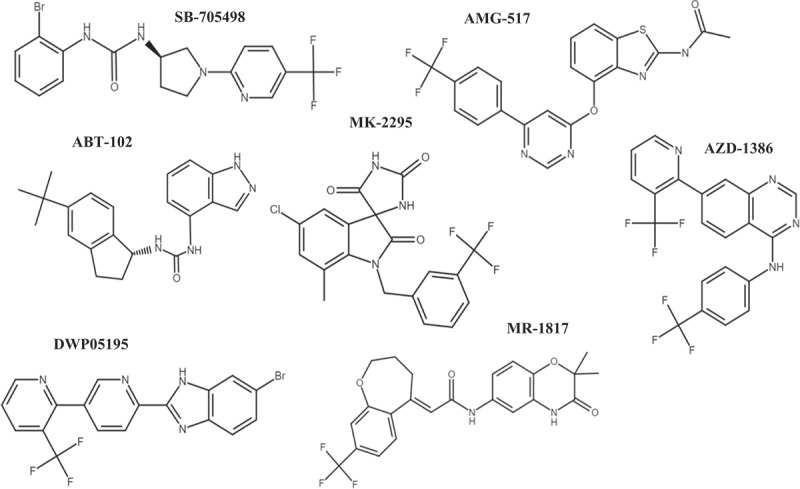 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsIon Channels and Receptors · Pain Mechanisms and Treatments · Cardiovascular, Neuropeptides, and Oxidative Stress Research
Introduction
TRPV1 which stands for Transient Receptor Potential Vanilloid 1, is a member of the Transient Receptor Potential (TRP) ion channel group [1] that plays a role in the process of phototransduction [2] and is denominated following the study made by Cosens and Manning in 1969 on the Drosophila mutant fly that initially brought attention to the TRP’s distinctive transient functionality [3]. The TRP channels are of greatest significance in various cellular processes that are intricately linked to adaptation, homeostasis [4], and pathological conditions by their ability to detect and respond to various stimuli such as temperature, pain, pressure, and chemical signals [1,5]. The TRPV1 protein assumes a pivotal function in the regulation of pain perception, exhibiting a notable presence within the sensory neurons where it is prominently expressed [6]. In a research work led by David Julius, Michael Caterina, and colleagues, it was determined that TRPV1 is the specific nociceptor responding to the stimulation induced by a compound found in red chili pepper known as capsaicin, that causes a spicy sensation [7]. They employed a calcium-based expression cloning strategy to investigate the structure of the capsaicin receptor. Initially, a complementary DNA (cDNA) library was generated from dorsal root ganglion cells, which are sensory neurons. The cDNA library was then transfected into the HEK 293 cell line. Through this process, a clone containing a 3-kilobase cDNA insert was identified. Remarkably, this clone conferred capsaicin sensitivity, leading to the discovery of the capsaicin receptor, known as TRPV1, in 1997 [7]. TRPV1 is a nonselective cation channel abundantly expressed in a wide array of cell types distributed throughout the body [8], and it has been reported that TRPV1 plays a role in the modulation of the immune system [9–11], consequently, investigations are examining the potential role of TRPV1 in the development and progression of autoimmune disorders [12].
Autoimmune diseases comprise a diverse spectrum of disorders such as multiple sclerosis, rheumatoid arthritis and type 1 diabetes, wherein the immune system erroneously directs its assault toward the body’s own healthy cells [13]. Recent studies have indicated that autoimmune diseases exert a clear impact on approximately 10% of the population [14] drawing therefore our attention to these widely destructive disorders and consequently, growing our interest in investigating the potential influence of the TRPV1 channel on autoimmune diseases. In this review, we will concisely explore the structure and function of TRPV1. Additionally, we will discuss its potential roles in autoimmune diseases, with a specific focus on its interaction with the immune cells. Furthermore, we will explore the relevance of TRPV1 in drug development for the treatment of autoimmune disorders.
Structure of TRPV1
The TRPV1 channel has a tetrameric architecture, with four identical subunits forming a cohesive and functional channel complex. These subunits interact with each other to assemble a central pore (Figure 1) [15,16]. The TRPV1 channel has six transmembrane (TM) regions that traverse the cell membrane. These regions, designated as S1, S2, S3, S4, S5, and S6, are composed of alpha-helices [15]. Figure 1.Structural architecture of the TRPV1 ion channel. Schematic diagram depicting the tetrameric structure of the TRPV1 channel. Each subunit consists of six transmembrane domains (S1-S6) that form the central ion-conducting pore. The pore loop region between S5 and S6 is critical for the selective permeability of ions such as Ca^2+^ and Na^+^. The N-terminal domain contains multiple phosphorylation sites and binding domains, while the C-terminal region includes a TRP domain and additional regulatory elements.
Furthermore, the helices (S1-S6) span the cell membrane and connect through a pore loop domain, positioned between the fifth and sixth transmembrane segments. The S5 and S6 segments contribute to the formation of the ion-conducting pore for ions like Ca^2+^ and Na^+^. This pore loop region is crucial for selectively allowing ions to pass through the channel (Figure 1) [17].
The arrangement and interactions of the transmembrane helices, along with the pore loop domain, determine the structure and function of the TRPV1 channel (Figure 1).
TRPV1 is uniquely sensitive to changes in membrane voltage [18], which take part in regulating its responses. They are very similar to the voltage-gated ion channels, including voltage-gated sodium, calcium and potassium channels [19]. Unlike typical voltage-gated ion channels, TRPV1’s voltage sensitivity appears critical for its ability to detect and transmit pain signals. This regulation does not come from the usual voltage-sensing region, but instead from negative charges in TRPV1’s outer pore [17].
The N-terminal region of the TRPV1 channel has multiple phosphorylation sites such as the protein kinase A (PKA) sites: S502, T370, T144, S116 [20], adenosine triphosphate ATP and calmodulin binding sites located on the ankyrin repeat domain that are formed by 6 ankyrin repeats of approximately 33 amino acids, which facilitate protein-protein interactions [21].
The C-terminal region of the TRPV1 channel contains a TRP domain, binding sites for substances like phosphatidylinositol 4,5-bisphosphate (PIP2) [19], and also multiple calmodulin binding domains [22]. The amino and carboxy terminals can interact with the calmodulin molecule, and the serine (S) and threonine (T) residues can be phosphorylated by calcium calmodulin kinase II (CaMKII), PKA, and protein kinase C (PKC) [23] (Figure 1) [20].
Biological function of TRPV1
TRPV1 is known as a heat receptor due to its high sensitivity to heat [7]. When exposed to temperatures above a specific threshold ( >43°C), TRPV1 becomes activated, resulting in the perception of heat and pain. This heat sensitivity enables TRPV1 to function as a temperature sensor, playing a role in temperature regulation and the detection of harmful heat (Figure 2) [7]. Figure 2.Biological function of TRPV1. Highlights of the diverse biological functions of the TRPV1 ion channel. As a thermosensor, TRPV1 responds to noxious heat stimuli, triggering the influx of calcium and sodium ions, leading to depolarization and the transmission of pain signals to the central nervous system. TRPV1 also plays a crucial role in the modulation of pain sensitivity, with phosphorylation-mediated sensitization and desensitization mechanisms. TRPV1 integrates various painful stimuli, including chemical signals, pH changes, and endogenous substances.
Transmission
3.1.
TRPV1’s pore opens between TM5 and TM6, allowing calcium and sodium ions to enter the cell [7,24,25]. This triggers depolarization and generates action potentials, transmitting pain signals to the CNS. TRPV1’s activation by noxious stimuli informs about tissue damage or harmful conditions [26]. TRPV1-expressing nociceptive neurons relay sensory information to the brain for pain perception and processing [27]. Thus, TRPV1 plays a significant role in pain transmission.
Modulation
3.2.
TRPV1 regulates pain perception and sensitivity through sensitization and desensitization [28]. Phosphorylation of TRPV1 channels, particularly at N- and C-terminals (S116, T144, T370, T704, and S800), enhances sensitivity via protein PKA and PKC [20,27,29]. PKC-mediated phosphorylation increases TRPV1’s openness and pain signal transmission, while PKA phosphorylation counters desensitization, restoring responsiveness [8,20] This leads to the release of neuropeptides like substance P and CGRP [30]. TRPV1 activation is linked to hyperalgesia (increased pain reactivity) and allodynia (painful response to non-painful stimuli) [30].
Integration
3.3.
TRPV1 is crucial in integrating diverse painful sensations, enabling perception and reaction to harmful inputs. It responds to endogenous substances like low pH, anandamide, N-oleyl-dopamine, and N-arachidonoyl-dopamine [31]. Capsaicin, an exogenous compound, selectively activates TRPV1 [8,32]. TRPV1 integrates these stimuli, contributing to pain perception, detection of noxious heat, temperature regulation, and response to chemical compounds and inflammatory mediators during injury or inflammation. TRPV1’s integration refers to its ability to detect and respond to various types of painful or noxious inputs, contributing to overall pain perception and processing.
TRPV1 and immune cells
TRPV1 is now known to be present and functional across the entire immune family (Table 1). Including macrophages, microglia, dendritic cells, mast cells, neutrophils, eosinophils, CD4^+^ and CD8^+^ T cells, regulatory T cells, B cells and innate lymphoid cells. Channel opening gives a rapid Ca^2 +^/Na^+^ pulse that tunes survival, migration, cytokine balance and effector activity, letting TRPV1 act as a context-dependent rheostat that can either amplify or resolve inflammation.Table 1.TRPV1 and the immune cells.IMMUNE CELLTRPV1 EXPRESSION IN IMMUNE CELLTRPV1 EFFECTCAPSAICIN(TRPV1 AGONIST)REFERENCEMACROPHAGETNF-αCytokineM1 macrophageIncreased secretion of TNF-αAnti-inflammatory effectAlleviate inflammation[33,34]T CELLCD4+ helper T cell fractionRequired for TCR signaling transduction in CD4+ T cellsN/A[38,39]B CELLAdaptive immune response to bacterial infectionModulate B cell immune responseN/A[43]DENTRIC CELLDermal DCBut other studies were unable to identify TRPV1 in DCEssential for the synthesis of lL-23 by dermal DCInduce DC maturation and migration to lymph nodes[41–43]NEUTROPHILNot clearly mentionedModulate neutrophil function and inflammationInduce Calcium flux in Neutrophils[47,49]
Macrophages
4.1.
Macrophages play a crucial role in immune processes like phagocytosis and cytokine production [33]. TRPV1 is expressed in macrophages and affects their function. In atherosclerosis, TRPV1 protects against lipid accumulation and TNF-α in macrophages [34]. TRPV1 activation in macrophages triggers pain hypersensitivity [35] and influences the tumor microenvironment [36]. In Parkinson’s disease, TRPV1 activation shifts macrophages/microglia from M1 to M2, increasing arginase-1 and CD206 while reducing iNOS and IL-6 [37]. In osteoarthritis, TRPV1 promotes a proinflammatory M1 macrophage phenotype [36].
T cells
4.2.
T cells, including CD4+ helper cells and CD8+ cytotoxic T lymphocytes, are crucial for immune defense against infections. TRPV1 is found in CD4+ T cells, exhibiting high mRNA expression and protein localization at the plasma membrane [38]. TRPV1 is necessary for effective TCR signaling in CD4+ T cells, as demonstrated by studies on mice and humans [38,39]. TRPV1 inhibition blocks TCR-mediated signaling pathways and cytokine expression in CD4+ T cells [39]. Furthermore, TRPV1 activation in CD4+ T cells inhibits direct HIV-1 infection through calcitonin gene-related peptide (CGRP) [40].
B cells
4.3.
B cells are essential for humoral immunity, producing antibodies to defend against infections [41]. The role of TRPV1 in B cells is not well understood. Studies using TRPV1 knockout mice showed no significant effect on B cell counts [42]. However, vagal TRPV1 neurons were found to regulate B cell populations during Streptococcus pneumoniae lung infection, as their removal reduced B cell infiltration [43]. Limited data exists on the TRPV1-B cell relationship. Piperine, a component of black pepper, affected B cells independently of TRPV1, suppressing activation markers and cytokine synthesis [43].
Dendritic cells
4.4.
TRPV1 expression and functional significance in dendritic cells (DC) are uncertain and inconsistent [44]. In the context of inflammation, TRPV1+ nociceptive neurons interact with dermal DCs, influencing IL-23 synthesis [41]. Some studies suggest TRPV1 expression on DCs, with capsaicin activating DC maturation and migration [41]. However, other studies failed to detect TRPV1 expression or observe calcium influx changes in DCs treated with capsaicin [42,44]. Discrepancies may be due to dosage, duration, or cell type variations. Capsaicin may affect DCs through non-TRPV1-mediated mechanisms [43]. Recent research supports functional TRPV1 expression in DCs, macrophages, and lymphocytes, showing calcium elevation and CGRP production in response to capsaicin, dependent on TRPV1 [43].
Neutrophils
4.5.
Neutrophils, comprising 70% of white blood cells, play a crucial role in the immune system [45]. Study reported that TRPV1 knockout mice showed increased inflammation and organ damage, with elevated neutrophil infiltration and proinflammatory cytokines [46]. Phytochemicals from Ferula akitschkensis [47] modify neutrophil responses via TRPV1 [48]. Neutrophils exhibit calcium fluxes upon exposure to capsaicin, and this response can be reduced by N-acetyl cysteine, a TRPV1 antagonist. In individuals with polycystic ovarian syndrome (PCOS), TRPV1 in neutrophils is involved in calcium influx, potentially contributing to the release of proinflammatory cytokines [49].
TRPV1 in autoimmune diseases
Autoimmune hepatitis
5.1.
TRPV1 plays a role in immune system modulation, particularly in autoimmune hepatitis. Studies have shown that TRPV1 mediates the anti-inflammatory effects of cannabidiol (CBD) [50]. CBD administration decreased pro-inflammatory cytokine levels and increased the presence of suppressor cells in a hepatitis model, but these effects were abolished in TRPV1-knockout mice [50]. TRPV1 activation by CBD may also contribute to its actions in immune function [51].
Multiple sclerosis
5.2.
In Multiple Sclerosis (MS), TRPV1 is involved in controlling neuroinflammation [52]. Genetic variations in the TRPV1 gene, specifically single-nucleotide polymorphisms (SNPs), may modulate its activity and impact neuroinflammation progression in MS [53]. The TRPV1 SNP rs222747 GG/GC genotype, associated with cortical excitability and pain modulation in MS patients, influences cerebrospinal fluid cytokine composition [53–55] However, in individuals with relapsing-remitting MS, the mRNA expression of TRPV1 and other TRP channels in peripheral blood mononuclear cells is decreased [56]. TRPV1 can be phosphorylated by calcium calmodulin-dependent kinase alpha (CaMKIIα), which plays a role in pain sensitivity and perception in MS, supporting TRPV1’s involvement in MS pain signaling [57].
Type 1 diabetes
5.3.
TRPV1 expression is elevated in sensory nerves of the pancreas [58] and is expressed on nerves innervating islet β cells, modulating T cell function. Capsaicin administration to neonatal mice destroyed TRPV1 neurons and protected them from autoimmune diabetes. This suggests that these neurons may induce type 1 diabetes by modulating T cell proliferation and activity through substance P release, TRPV1 as a potential therapeutic target for Type 1 diabetes [59].
Rheumatoid arthritis
5.4.
Study showed that TRPV1 knockout mice showed reduced arthritis progression after adjuvant treatment [60]. Increased TRPV1 expression in synovial fibroblasts of arthritis patients correlates with higher cytokine production [61]. TRPV1 agonists like capsaicin can alleviate arthritic pain by activating TRPV1 and inducing pain desensitization [62].
Systemic lupus erythematosus
5.5.
TRPV1 is linked to chronic inflammatory pain and neuropeptide dysregulation in systemic lupus erythematosus (SLE). Capsaicin release of substance P increases sensitivity in SLE patients [63]. TRPV1 is expressed in endothelial colony-forming cells (ECFCs) and its activation promotes angiogenesis through anandamide uptake [64]. Optical stimulation of TRPV1 in ECFCs boosts proliferation and tube formation, potentially aiding vascular healing in SLE [65].
Therapeutic approaches
TRPV1 has been extensively studied for pain management [28,66,67]. Capsaicin, derived from chili peppers [68], is the most recognized TRPV1 activator. It is FDA-approved for shingles-associated nerve pain and has shown promise in relieving pruritus in psoriasis [69]. Other TRPV1 agonists, like resiniferatoxin (RTX) found in the Euphorbia plant, have demonstrated analgesic effects in rheumatoid arthritis [70].
Capsazepine is a competitive antagonist of capsaicin, blocking capsaicin-induced responses in dorsal root ganglion (DRG) neurons [71]. It interacts with and binds to TRPV1, specifically in the transmembrane region and the pore between transmembrane 5 and 6 (TM5, TM6) [72]. By blocking the intracellular passage of ions like Ca^2+^, which is crucial for signal transduction and immune system activation [73]. Although Capsazepine has shown therapeutic potential in conditions like cancer, hepatitis, and malaria [72], it has not undergone clinical trials and has not been specifically studied for autoimmune disorders. Several compounds, including SB-705498, AMG-517, ABT-102, MK-2295, AZD-1386, DWP05195 and MR-1817 (Figure 3), have been synthesized and tested in clinical trials as TRPV1 antagonists [72,74]. Figure 3.Chemical structure of TRPV1 antagonists in clinical trials. Overview of several TRPV1 antagonists that have been investigated in clinical trials for various therapeutic applications. The compounds SB-705498, AMG-517, ABT-102, MK-2295, AZD-1386, DWP05195, and MR-1817. Each of these TRPV1 antagonists has been evaluated for their potential to modulate the activity of the TRPV1 channel and their potential therapeutic benefits, particularly in the context of pain management and other clinical conditions.
The TRPV1 antagonist SB-705498, developed by GlaxoSmithKline, was the first to undergo human testing [74]. It was evaluated as an intranasal treatment for rhinitis in Phase II trials (NCT01424397, NCT01439308) [75,76]. Additionally, it was investigated for potential therapeutic effects on migraine, pain after dental surgery, and rectal hypersensitivity in 2005. However, these trials were terminated the following year without reported data [77].
AMG-517 is a potent TRPV1 antagonist with an IC_50_ of less than 10 nM. It is available in oral form and selectively inhibits the TRPV1 channel, showing lower potency against other TRP channels (with an IC_50_ value of >5–20 µM) [78,79] However, the AMG-517 trial reported hyperthermia as an adverse effect, leading to the discontinuation of the Phase I trial [80,81].
ABT-102, developed by Abbott, is a TRPV1 antagonist that inhibits all modes of
TRPV1 activation [82]. It has been found to effectively increase body temperature in both animal models and human subjects [83,84] In a Phase I clinical trial (NCT00854659), ABT-102 administration led to an average increase of 0.6°C in core body temperature, with the highest recorded temperature reaching 38.7°C. Repeated administration of ABT-102 showed favorable tolerability, as there were no significant temperature increases observed by day 7. However, ABT-102 was also found to have a negative impact on the perception of painful heat, and this deficiency did not improve with a 7-day administration schedule [84].
The TRPV1 antagonist MK-2295, developed by Merck and Neurogen, increases thresholds for painful heat sensitivity, clinically impairing the perception of potentially harmful heat [85]. In a simulated bathing experience, participants taking MK-2295 took longer to perceive heat discomfort and remove their hands from a 49°C water bath. Furthermore, a lower percentage of MK-2295 recipients perceived 70°C water as too hot compared to the placebo group [85]. These effects were observed at doses similar to those providing pain relief.
AZD-1386 from AstraZeneca did not cause significant hyperthermia [86]. It provided acute but short-duration analgesia effects for postoperative dental removal [87]. However, in knee osteoarthritis (OA) patients, four weeks of twice-daily administration did not reduce OA discomfort [88]. The study was terminated due to elevated liver enzymes with 90 mg doses. AZD-1386 did not provide analgesia for esophageal thermal pain but increased cutaneous thermal tolerance in non-erosive gastroesophageal reflux disease (NERD) patients compared to the placebo group [88].
DWP05195, a TRPV1 antagonist synthesized by Daewoong Pharmaceutical, underwent three clinical investigations. The earliest trial (NCT00969787) assessed the safety, tolerability, pharmacokinetics, and pharmacodynamics of orally administered DWP05195 in healthy adult males [89]. The second study (NCT01094834) evaluated multiple concentrations of DWP05195 for safety, tolerability, pharmacokinetics, and pharmacodynamics in healthy adults [90]. The most recent trial (NCT01557010) investigated the efficacy and tolerability of DWP05195 in post-herpetic neuralgia patients [91]. However, no results from these trials have been reported.
MR-1817 is indicated for pain, and its initial human investigation aimed to assess its safety and tolerability [92]. However, no study data have been reported.
Conclusion
TRPV1 plays a putative role in autoimmune diseases. Its structure and biological functions, including transmission, modulation, and integration, contribute to its implication in various autoimmune disorders. TRPV1 is closely associated with different immune cells, such as macrophages, T cells, B cells, DCs, and neutrophils, further highlighting its involvement in immune responses. Autoimmune hepatitis, multiple sclerosis, type 1 diabetes, rheumatoid arthritis, and systemic lupus erythematosus are among the autoimmune diseases where TRPV1 has been implicated.
Several therapeutic approaches targeting TRPV1 have been explored, including capsaicin, capsaicin analogs like capsaizapine, and other selective TRPV1 modulators such as SB-705498, AMG-517, ABT-102, MK-2295, AZD-1386, DWP05195, and MR-1817.
Understanding the structure, function, and implications of TRPV1 in autoimmune diseases provides a foundation for future research and the development of innovative therapeutic strategies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhang M, Ma Y, Ye X, et al. Trp (transient receptor potential) ion channel family: structures, biological functions and therapeutic interventions for diseases. Sig Transduct Target Ther. 2023;8(1):261. doi: 10.1038/s 41392-023-01464-x PMC 1031990037402746 · doi ↗ · pubmed ↗
- 2Hardie RC, Juusola M. Phototransduction in Drosophila. Curr Opin Neurobiol. 2015;34:37–12. doi: 10.1016/j.conb.2015.01.00825638280 · doi ↗ · pubmed ↗
- 3Cosens D, Manning A. Abnormal electroretinogram from a Drosophila mutant. Nature. 1969;224(5216):285–287. doi: 10.1038/224285 a 05344615 · doi ↗ · pubmed ↗
- 4Vandewauw I, De Clercq K, Mulier M, et al. A TRP channel trio mediates acute noxious heat sensing. Nature. 2018;555(7698):662–666. doi: 10.1038/nature 2613729539642 · doi ↗ · pubmed ↗
- 5Smani T, Shapovalov G, Skryma R, et al. Functional and physiopathological implications of TRP channels. Biochim Biophys Acta (BBA)-Mol Cell Res. 2015;1853(8):1772–1782. doi: 10.1016/j.bbamcr.2015.04.01625937071 · doi ↗ · pubmed ↗
- 6Julius D. Trp channels and pain. Annual review of cell and developmental biology. Annu Rev Cell Dev Biol. 2013;29(1):355–384. doi: 10.1146/annurev-cellbio-101011-15583324099085 · doi ↗ · pubmed ↗
- 7Caterina MJ, Schumacher MA, Tominaga M, et al. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 1997;389(6653):816–824. doi: 10.1038/398079349813 · doi ↗ · pubmed ↗
- 8Shuba YM. Beyond neuronal heat sensing: diversity of TRPV 1 heat-capsaicin receptor-channel functions. Front Cell Neurosci. 2021;14:612480. doi: 10.3389/fncel.2020.61248033613196 PMC 7892457 · doi ↗ · pubmed ↗