Therapeutic potential of MSCs and their exosomes in hepatic Ischaemia-Reperfusion injury: a systematic review and meta-analysis of rodent studies
Yanxi Mu, Weixiong Zhu, Wentao Ma, Yu Cheng, Bo Ren, Yusheng Cheng, Wence Zhou

TL;DR
This study reviews and analyzes how mesenchymal stem cells and their exosomes can help reduce liver damage in rodents, offering potential for future clinical use.
Contribution
The paper provides a comprehensive meta-analysis of MSCs and exosomes in rodent HIRI models, highlighting their therapeutic mechanisms and outcomes.
Findings
MSCs and exosomes significantly reduced liver injury markers like ALT, AST, and inflammation in rodent models.
Therapeutic effects were linked to antioxidative, anti-inflammatory, and anti-apoptotic pathways.
Heterogeneity in results was attributed to variations in dosage, administration routes, and reperfusion durations.
Abstract
This meta-analysis comprehensively evaluates the therapeutic efficacy and mechanisms of mesenchymal stem cells (MSCs) and their exosomes in rodent models of hepatic ischemia-reperfusion injury (HIRI), providing preclinical support for future clinical translation. In accordance with the PRISMA guidelines, we systematically searched PubMed, Web of Science, Embase, Cochrane Library, and ClinicalTrials.gov for studies published from inception to January 13, 2025, and identified 64 eligible studies. Risk of bias was evaluated using the SYRCLE tool, and Review Manager 5.4.1 was employed for meta-analysis, calculating SMD and 95%CI. Primary outcomes included liver function (ALT/AST), histopathological scores (Suzuki’s score, necrotic area ratio), inflammatory cytokines (TNF-α), and apoptosis markers (c-caspase 3). MSCs and their exosomes significantly ameliorated HIRI. In the 60-minute…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
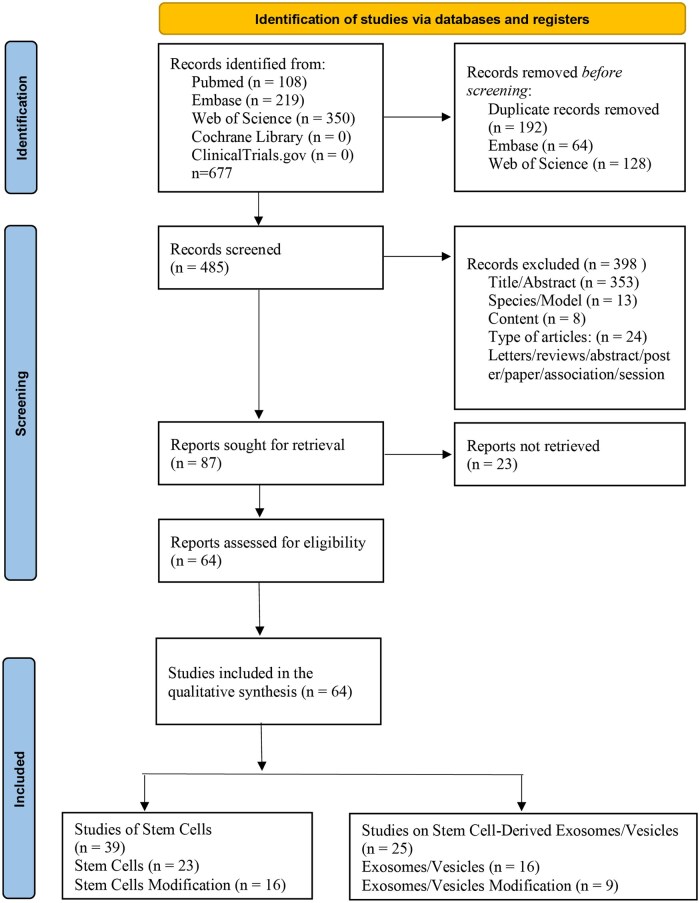 Figure 1
Figure 1 Figure 2
Figure 2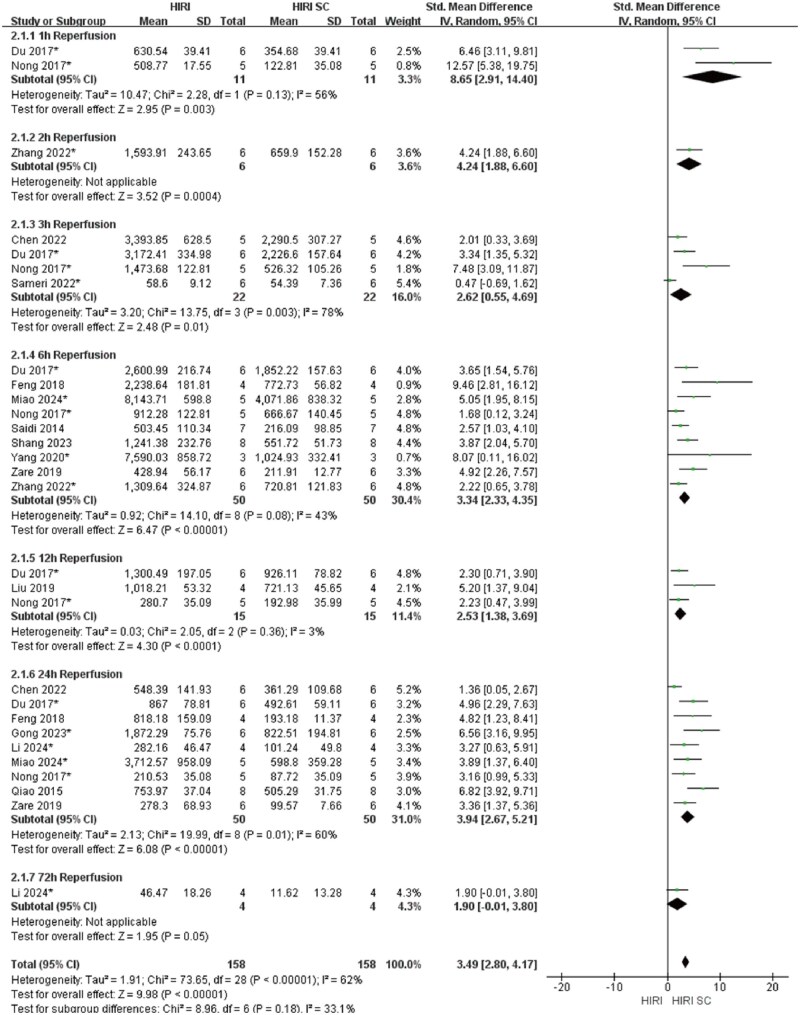 Figure 3
Figure 3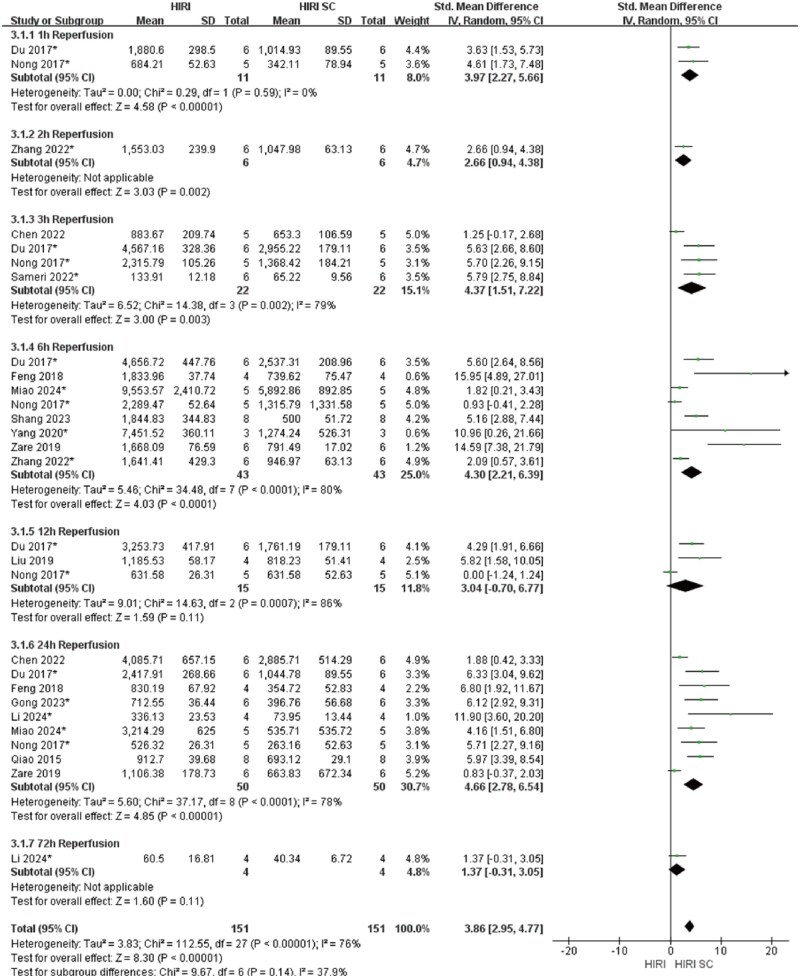 Figure 4
Figure 4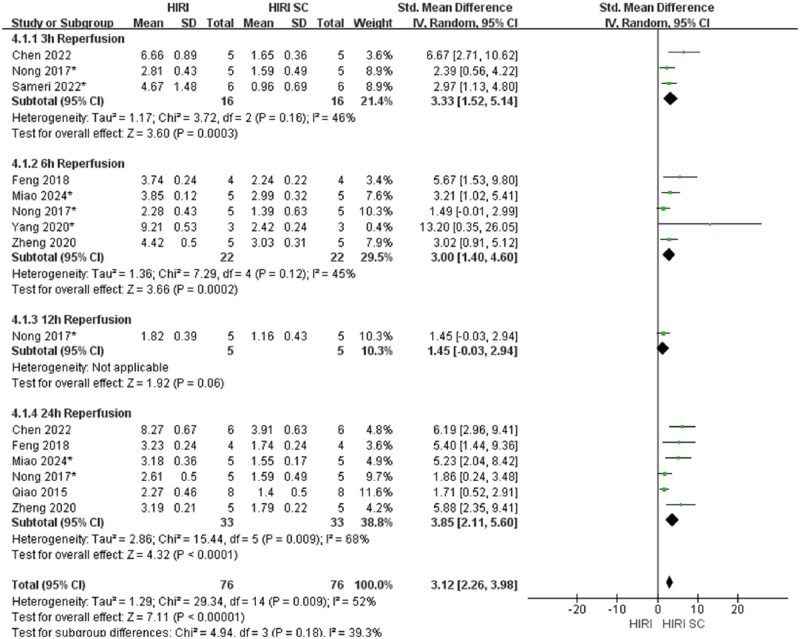 Figure 5
Figure 5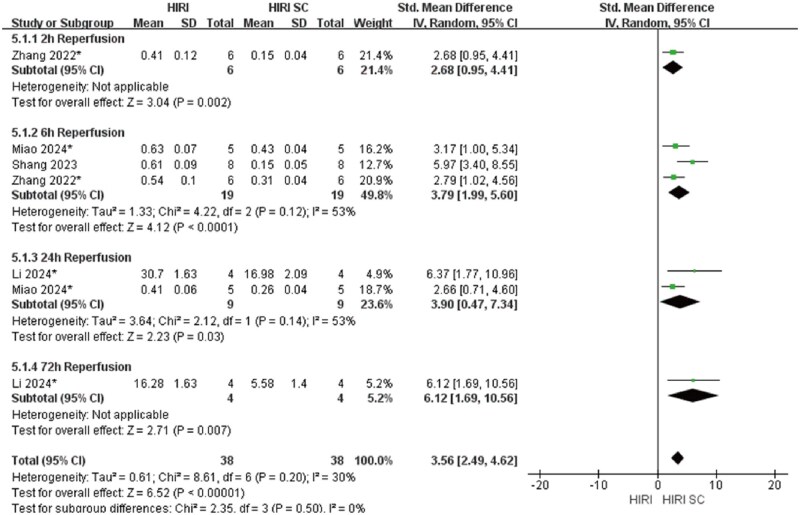 Figure 6
Figure 6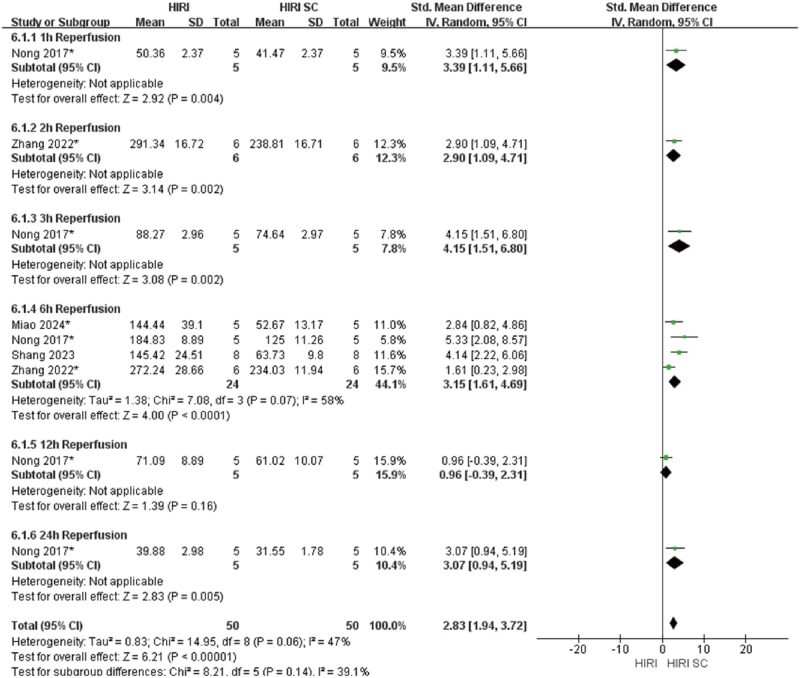 Figure 7
Figure 7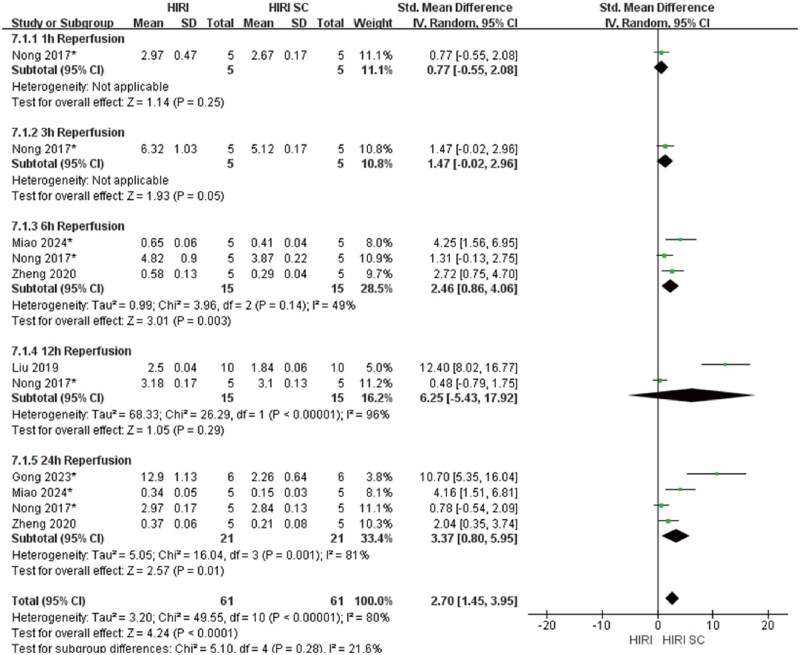 Figure 8
Figure 8| Author | Country | Strain (number) | block liver’s blood supply | HIRI Duration | SC Types (modification) | SC Administration Route | SC Dosage | SC therapy time | Positive surface markers |
|---|---|---|---|---|---|---|---|---|---|
|
| |||||||||
|
| China | C57BL/6 mice(10) | 70% | I90minR6h | hMSCs | the portal vein | 1.0 × 105cells/mice | after 90 min of ischemia and 6 h of reperfusion | CD90, CD73 and CD105 |
|
| China | C57BL/6 mice(104) | 70% | I60minR6h | MSCs | the caudal vein | 1.0 × 106 | 30 min prior to hepatic warm I/R | NE |
|
| Turkey | Wistar albino rat(30) | 70% | I40minRx | ADSCs | liver parenchyma | 1.0 × 106cells/kg | following 20 min of ischemia | NE |
|
| Japan | Wistar rats(37) | 70% | I60minR3/24h | ASCL | intrasplenic | 1.0 × 106 | 24 h before HIRI | CD44 and CD90 |
|
| Iran | BALB/c | the portal vein, hepatic artery, and bile duct of the liver | I60minR1/5h, 7d | AMSCs/BMSCs | the tail vein | 1.0x106 | Immediately after reperfusion onset | CD29, CD73, CD105, and CD90 |
|
| China | SD rats(30) | 70% | I30minR24h | ADSCs | the tail vein | 2.0 × 106 | 2 h before surgery | NE |
|
| China | C57BL/6 mice(10) | 70% | I60minR6/24h | UC-MSCs | peripheral vein | 1.0x106/100 μl | after reperfusion | NE |
|
| China | Gene editing mice | NE | I90minR6h | MSCs | the tail vein | 1 × 106cells/mouse | 24 hours before ischemia | NE |
|
| Iran | BALB/c(24) | 70% | I60minR6/24h | BM-MSCs | the portal vein | 1.0x106 | Immediately after reperfusion | Sca-1, CD44 |
|
| China | Wistar rats(30) | the portal triad for the left and median liver lobes | 6h (I60minR6h) | BM-MSCs | penis dorsal vein | 1.0x106 | 30 min prior to hepatic warm I/R | CD29, CD44 and CD90 |
|
| China | C57/B6 and inbred Buffalo rats | the right hepatic pedicle | I60minR6h, 1/3/5d | hiPSC-MSCs | the portal vein | 5 × 104 cells/100µL PBS | after reperfusion | NE |
|
| China | SD rats | the hepatic artery and portal vein to the left lateral and median lobes | I60minR12/24/72h, 14d | MSCs | the portal vein | 3.0 × 105 | Reperfusion was initiated | CD29, CD44, and CD90 |
|
| Japan | F344 wild-type rats(60) | 70% | I60minR1/3/7d | BM-MSCs | the tail vein | 1.5x106 | 15 min after initiating reperfusion | NE |
|
| Netherlands | C57BL/6(49) | 70% | I60minR6/48h, 5d | MSC | the tail vein | 2.0 × 105 | 2 hours before and 1 hour after ischemia | NE |
|
| China | SD rat(60) | NE | I40minR1/2/3W | hAD-MSCs | the tail vein | 0.5 mL | 2 weeks after HIRI model | NE |
|
| Poland | Wistar rats(20) | the portal triad under the left lateral lobe and left median lobe | I60minR3M | BM-MSCs | intraportally | 1.0x106 | After IRI | NE |
|
| Korea | BALB/c mice(100) | the portal vein, hepatic artery, and bile duct above the branching to right lateral lobe. | I45minR3/6/12/24h | ASCs | the tail vein | 1.0x106 | within 1 hour of HIRI | CD90 |
|
| Netherlands | C57BL/6 mice(32 + 16) | 70% | I90minR6/24h, I60minR48h | MSC-CM | intraperitoneally | 200 μL | at the end of surgical procedure. | NE |
|
| USA | C57BL/6(28) | 70% | I60minR6/12/18/24h | HADMSCs | the tail vein | 1–2 million | 30 minutes before ischemia | CD105, CD29, CD44, CD90 |
|
| Japan | BALB/c nu-nu mice | the hepatoduodenal ligament | I15minR6/24h, I20minR24h | h-ADSCs | the tail vein | 1.0x105/0.1 ml/mouse | after the hepatectomy | CD29, CD44, CD73, CD90, CD105, and CD166 |
|
| Taiwan, China | Fisher rats(30) | the left lobe liver | I60minR72h | ADMSCs | venous | 1.2x106 | immediately, 6 hrs, and 24 hrs after reperfusion | CD-90 and CD-29 |
|
| Japan | Wistar rats | the hepatoduodenal ligament | I15minR1/2d | ADSCs | the penile vein | 2x106cells/rat | After releasing the clamp | NE |
|
| China | SD rats | the hepatic artery, portal vein, and bile duct to left lateral and median lobes of liver. | I60minR6h, 1/2/3/5d | MSCs | the tail vein | 3.0x106 | 2 h before the surgical operation | CD54 and CD90 |
|
| |||||||||
|
| China | C57BL/6 mice | the artery/portal vein blood supply to left and middle liver lobes. | I90minR2/6/12/24/48h | UC-MSCs(MMCLs) | the tail vein | NE | after the initiation of reperfusion | NE |
|
| China | C57 mice(24) | 70% | I90minR6h | MSCs(CD47-overexpressed) | the tail vein | 1.0x106 | 24 h before surgery | NE |
|
| Taiwan, China | SD rats (50) | NE | I60minR18/72h | tacrolimus-ADMSCs | intravenous | 1.2 × 106 | 3h after IR | NE |
|
| China | C57BL/6J(27) | 70% | I60minR6h/7d | MenSCs(Interferon-γ) | intravenous | 1.0x106/100 μL | 1h before the surgery | CD29, CD73, CD90, and CD105 |
|
| China | SPF SD rats(36) | 70% | I80minR24h | BMMSCs(HO-1) | caudal and portal vein | 2.0 × 106 | 1 day before surgery and during reperfusion | CD90, CD29, and RT1A |
|
| UK | C57BL/6 mice | NE | I60minR24h | PDGFRα/Sca-1 (PaS) sorted MSC | intraperitoneal | 1.0 × 106 | One hour prior to the induction of ischemia | NE |
|
| China | Wistar albino rats | NE | I45minR24h | BMSCs(Baicalin Combined) | the portal vein | 1.0x106 | after reperfusion | CD29, CD44 and CD105 |
|
| Korea | wild-type ICR mice(25) | 70% | I90minR24h | MSC(Nanozyme Impregnated) | hepatic vein | 1 × 105cells/mice | 90 min post hepatic ischemia | CD90 and CD29 |
|
| China | Wistar rats(80) | 70% | I60minRx | SOD2-BMMSCs | the tail vein | 1 × 106PKH26-labeled | NE | Bax, Bcl-2 and caspase-3 |
|
| China | SPF SD rats | the left outer and left middle hepatic artery trunks | I30minRx | BMSCs(TNF-α) | the portal vein | 0.5ml | opened the blood vessels | CD79, CD45, CD90, and CD29 |
|
| China | C57BL/6 mice (36-42) | 70% | I90minR24h | UC‐MSCs(rapamycin) | the peripheral vein | 1 × 106cells/100 µL per mouse | After model was established | CD105, CD44, CD29, CD90, CD73 and CD166 |
|
| China | SD rats(90) | 70% | I60minR12h | ADMSCs(Mild hypothermia combined) | the femoral vein | 1.0x109 | 30min before ischemia | NE |
|
| China | SD rats(24) | 70% | I90minR6/24/48h | UC-MSCs(Spheroid-cultured) | intraperitoneal | 3 × 106 per rat | after reperfusion | NE |
|
| China | SD rats(44) | 70% | I60minR6/24h | hBM-MSCs(Intravenous Anesthetics) | the portal vein | 5.0 × 104 | NE | NE |
|
| China | SD rats(32) | 70% | I60minR24h | MSC/HSP-MSCs(HSP) | the portal veins | 1.0x106 | The clamp was removed | NE |
|
| China | SD rats(30) | the portal triad | I30minR6/24h | MSCs(N-acetyltransferase 8) | the tail vein | 3.0x106 | reperfusion was initiated | CD29, CD44, CD73, CD90, and CD105 |
|
| |||||||||
|
| China | Kunming mice(30) | the mid and left lobes’ portal vein and hepatic artery. | I60minR6h | BMSC-Evs | the tail vein | 50 μg in 100 μL PBS | 24 hours before surgery | CD73, CD90, CD63, TSG101 and CD9, |
|
| China | KM mice(30) | the portal vein and hepatic artery of the middle and left lobes. | I60minR3/6/9h | BMSC-exosomes miR-25-3p | the tail vein | 50 μg in 100 μL PBS | 24 h before surgery | CD73, CD90, CD63, TSG101 and CD9 |
|
| China | SD rats(30) | 70% | I60minR24h | ADSCs-exo | the tail vein | 30mg/kg | NE | CD90, CD105, CD29, and CD73 |
|
| China | SD rats (n=41) | 70% | I60minR2/6h | ADSCs-exo | the portal vein | 50 µl (30 µg) | NE | CD73, CD90 and CD105 |
|
| China | SD rats(24) | the median and left lateral liver lobes | IxR24h | ADSCs-Exo | the tail vein | 2 × 106 ADSCs or 100 gADSCs-Exo | immediately after the operation | NE |
|
| China | SD rats(24) | 70% | I30minR24h | ADSCs-exo | the tail vein | 100 μg | After surgery | CD29, CD44 and CD90 |
|
| Italy | C57BL/6 mice(38) | 70% | I90minR6h | HLSC-EV | the tail vein | 3 × 109 or 7.5 × 109 | Immediately after reperfusion | CD9, CD63, CD81, CD29, CD44, CD105 and CD49e, CD142, CD146, SSEA-4, and MCSP |
|
| China | C57BL/6, B6 mice | 70% | I90minR6h | UC-MSC-EVs | peripheral intravenous | 1.0x106 | after reperfusion | CD63, CD9 TSG101, and ALIX |
|
| China | C57BL/6 mice(36) | the artery/portal vein blood supply to the left and middle liver lobes. | I90minR6/12/24h | hUCB-MSCs-exo | intravenously | 10 μg | following 90 min of ischemia | NE |
|
| China | C57BL/6 mice(42) | the artery/portal vein blood supply to the left and middle liver lobes. | I90minR6/12/24h | hUCB-MSCs-exo | the portal vein | 2.5 × 1012 particles for each moue | after the initiation of reperfusion | CD29, CD105, CD9 and CD63 |
|
| China | SPF SD rats(43) | the medial and left hepatic hila | I45minR12h | MSC-exo | the tail vein | 500 μg of exosomes dissolved in 1 mL of PBS solution | 24 hours before the surgery. | CD29 and CD90 |
|
| China | SD rats | 70% | I90minR6/24h | hucMSC-EVs | the tail vein | 10 mg/kg | immediately after reperfu sion | CD29, CD90, CD105, CD73, CD63 and CD9 |
|
| Germany | C57BL/6 mice(25) | 70% | I90minR1/2/3d | hMSCs-EV | V. cava inferior | NE | After a midline laparotomy | CD105, CD90, CD73, CD9 and CD63 |
|
| China | C57Bl/6 mice(18) | 70% | I60minR1/3/6/12/24h | hiPSC-MSCs-Exo | inferior vena cava | 2.5 × 1012 particles | immediately after the initiation of reperfusion | CD29, CD73, CD90 and CD105 |
|
| China | SD rats | 70% | I60minR1/3/6/12/24h | hiPSCMSCs-Exo | inferior vena cava | 600 μg | immediately after the initiation of reperfusion | CD29, CD73 and CD90 |
|
| USA | C57BL/6(25) | 70% | I90minR1/3/6h | MSC-EV | the tail vein | 2.0x1010 | 30 minutes prior to ischemia | NE |
|
| |||||||||
|
| China | C57BL/6 mice | 70% | I90minR1/3/6h | Baicalin-pretreated BMSCs-Exos(Ba-Exo) | the tail vein | NE | after 90 min of ischemia and 6 h of reperfusion | CD9, CD63, CD81, TSG101 |
|
| China | SD rats(30) | the Glisson’s blood supply to the middle and left lobes of the liver. | I30minRx | Metformin and ADSCs-Exo complex (Met-Exo) | the tail vein | 100ug | immediately after the operation | ALIX, CD9, CD81 and TSG101 |
|
| China | C57BL/6 albino mice | 70% | I60minR1/3/7/14d | Phosphatidylserine-mediated MSC-Evs | the tail vein | 100 µg/100 µL per mouse | after 12 h of reperfusion | CD9, TSG101, Alix, CD73, CD90 and CD105 |
|
| China | C57BL/6(25) | 70% | I60minR6/24h | MSC-EVs containing GAS6 | the tail vein | 2 × 1010 particles/body | immediately after the restoration of blood supply | Alix, TSG101, CD9 |
|
| Iran | BALB/C mice(24) | 70% | I60minR3h | MSC-Exo/H2S-Exo(NaHS (1 mmol)) | the tail vein | 100 mg | ending the time of ischemia | CD9 and CD63 |
|
| China | SD rats(30) | 70% | I80minR24h | BMMSCs-exo(Heme Oxygenase-1) | tail and portal vein | 2.0 × 106 | 1 day before surgery and immediately after reperfusion | CD29, CD90, and RT1A |
|
| China | SD rats(48) | 75% | I60minR24h | UC-MSCs(MiR-20a-containing exosomes) | NE | 20 mg | NE | CD73, CD90, CD105, CD34 and CD45 |
|
| China | C57BL/6 mice | 70% | I60minR6h | BMMSC-Derived Hepatocyte-Like Cell Exosomes(MSC-Heps-Exo) | the tail vein | 100 μg | before and after the operation | CD29, CD44, CD105, Sca-1, CD9, CD63, and TSG101 |
|
| China | SD rats(40) | the portal venous and arterial blood supply to central and left liver lobes. | I90minR6h | MSC-exo(glycyrrhetinic acid(GA)) | intraperitoneal shot | NE | NE | NE |
- —National Natural Science Foundation of China10.13039/501100001809
- —Medical Innovation and Development Project of Lanzhou University
- —Major Science and Technology Projects of Gansu Province
- —Joint Research Fund General Projects of Gansu Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsExtracellular vesicles in disease · Organ Transplantation Techniques and Outcomes · Mesenchymal stem cell research
Introduction
Hepatic ischemia-reperfusion injury (HIRI) is a critical clinical challenge in liver surgery, frequently encountered in hepatectomy, liver transplantation, and hemorrhagic shock.1 HIRI is a major risk factor for primary graft dysfunction or failure and a key trigger for acute and chronic rejection.1 Hypoxia-induced HIRI involves an exogenous antigen-dependent local inflammatory response.2 Persistent HIRI not only causes hepatocyte apoptosis and necrosis, which can result in severe liver failure, but also provokes a substantial release of inflammatory mediators, thereby inducing systemic inflammatory response syndrome (SIRS). This further leads to multiple organ dysfunction syndrome (MODS), and may ultimately progress to multiple organ failure, significantly increasing the mortality rate of patients.
In recent years, stem cell-based therapies have emerged as a prominent strategy for mitigating HIRI, owing to their unique capacities for tissue repair and immunomodulation. Compared with conventional pharmacologic interventions, stem cell therapy offers multi-target and durable therapeutic benefits with an established safety profile in clinical applications. Among various stem cell populations, mesenchymal stem cells (MSCs) are regarded as the most clinically translatable cell type because of their broad tissue availability, ease of isolation, and robust immunoregulatory properties. MSCs reside in multiple mesenchymal tissues including bone marrow, adipose tissue, umbilical cord, and dental tissues and are characterized by adherence to plastic, expression of CD73, CD90, and CD105, lack of CD34, CD45, and HLA-DR expression, and tri-lineage differentiation potential toward osteogenic, chondrogenic, and adipogenic fates.3 Increasing evidence further indicates that the therapeutic actions of MSCs are predominantly mediated by their rich paracrine activity rather than by direct differentiation. MSCs-derived cytokines, chemokines, and extracellular vesicles, particularly exosomes, can reprogram innate and adaptive immune responses, attenuate excessive inflammation, and promote tissue repair and regeneration.3^,^4 Notably, MSCs derived from different tissues display substantial heterogeneity in proliferative potential, secretory profiles, and immunomodulatory potency; these variations are shaped by microenvironmental cues, inflammatory status, and metabolic reprogramming.5
Within the context of HIRI, MSCs-based therapy has gained considerable attention due to its pronounced immunomodulatory and regenerative capacities. Accumulating preclinical evidence demonstrates that MSCs rapidly home to the injured liver and ameliorate tissue damage by suppressing proinflammatory cytokine release, attenuating oxidative stress and apoptosis, and restoring microenvironmental homeostasis.6^,^7 Importantly, these therapeutic benefits also rely heavily on MSCs-derived paracrine mediators, including exosomes and diverse soluble factors that orchestrate immune regulation and tissue protection.8 Despite promising outcomes, substantial variation remains across studies regarding cell dose, timing, and route of administration, underscoring the need for greater standardization to optimize clinical translation.
This study aims to systematically review and meta-analyze existing rodent experimental evidence to evaluate the pooled effects of MSCs and their exosomes on liver function (ALT/AST) and histopathological outcomes across different ischemiareperfusion time points, and to explore potential sources of interstudy heterogeneity. The findings are expected to provide evidence-based insights to inform the design of future clinical trials and to facilitate the translation of stem cell therapy from preclinical research to precision medicine.
Materials and methods
Study selection
The inclusion criteria for the study are as follows: (1) In vivo rodent models (mice or rats); (2) The experimental group receive stem cell therapy; (3) The primary outcome index: HIRI; (4) The research was published in English. The exclusion criteria for the study are as follows: (1) There was no control group in the study; (2) Type of articles: Case reports, conference abstracts, posters, reviews, meta-analyses and letters from readers; (3) Lack of the full text; (4) No result or incomplete data; (5) Duplicate records. In this study, two researchers independently screened the titles, abstracts, and full texts, with any disagreements resolved through consensus discussion involving a third reviewer.
Protocol and registration
This study adhered to the methodological standards established by the EQUATOR Network (Enhancing the Quality and Transparency of Health Research), and reported according to PRISMA guidelines (Figure 1). The research protocol was prospectively registered in the PROSPERO international prospective register of systematic reviews (registration number: CRD420250638552).
Prisma flow diagram. Search strategy and selection process of the included studies.
Search strategy
This study adopted a systematic literature retrieval method, and cross-database retrieval was independently carried out by two researchers. The search scope includes the following core biomedical databases: PubMed, Web of Science, Embase, Cochrane Library, and ClinicalTrials.gov. Search strategies followed the PICOS framework using the following keywords: (1) “hepatic ischemia-reperfusion injury” OR “liver IRI” OR “HIRI”; (2) “stem cells” OR “mesenchymal stromal cells” OR “MSCs”; (3) “murine” OR “rodent” OR “animal model. The search was conducted from database inception to January 13, 2025, placing particular emphasis on studies published after January 2020. The complete details of the search strategy have been presented in detail in Supplementary Table S1.
Data extraction
A standardized data extraction process was applied. Two researchers independently extracted data based on the pre-designed PICOS framework table. Disputes arising during the extraction process were resolved through consultation or third-party arbitration. For data with opaque data or lacking relevant information, such as the mean, standard deviation, or standard error of the mean as described in the study, the missing data (including the sample size) were requested from the authors via email. Additionally, data were extracted from figures using WebPlotDigitizer (https://apps.automeris.io/wpd/) to extract data from studies where digital data were not available. Finally, we excluded studies from the meta-analysis that did not provide sample sizes even after contacting the authors. All steps followed PRISMA guidelines, with 10% of extracted data cross-checked for quality assurance.
Quality and risk of bias assessments
This study employed the SYRCLE animal experiment bias risk assessment tool.9 Two trained researchers independently evaluated the 10 key methodological areas included in the study, including random sequence generation, allocation concealment, and blinding. Each field is judged according to the three-level standards of low risk, uncertain risk, or high risk. Disagreements in the assessment shall be resolved through consultation or third-party arbitration, with final results recorded using a dual-entry system.
Statistical analysis
Data analysis was conducted using Review Manager software (Version 5.4.1) for statistical analyses and figure generation. Meta-analysis was conducted only when ≥3 studies were available. In the overall effect test, P<.05 was set as statistically significant. To account for methodological heterogeneity (e.g., differences in outcome assessment or interventions), a random-effects model was used for analysis. Since different measurement scales were used in each study to evaluate the same outcome measure, continuous data were summarized as standardized mean differences (SMD) with 95% confidence intervals (CI). Heterogeneity was assessed using the I^2^ statistic.10
Results
Search results
After screening 677 records (PubMed: 108, Embase: 219, Web of Science: 350; Cochrane Library and ClinicalTrials.gov: 0 each), 192 duplicate records were removed (Embase: 64, Web of Science: 128). Subsequently, 485 records were screened, of which 398 were excluded for failing to meet eligibility criteria. Ultimately, 87 full-text reports were retrieved for full-text assessment, of which 23 were unavailable. The remaining 64 reports underwent eligibility evaluation. Qualitative synthesis included 64 studies, comprising 39 stem cell-related studies (23 unmodified and 16 modified stem cells) and 25 studies on stem cell-derived exosomes/vesicles (16 unmodified exosomes/vesicles and 9 modified exosomes/vesicles) (Figure 1).
General characteristics of the included studies
The 64 eligible studies were published between 2012 and January 13, 2025. Approximately 53% (n = 34) were published in 2020 or later, indicating increasing research interest in stem cell-based therapies for HIRI. These studies originated from 11 countries, with China contributing the majority (72%, n = 46), followed by Japan, Iran, and the Netherlands (Table 1).
Characteristics of HIRI models
Animal species
All studies employed rodents, including mice (24 studies, 37.5%) and rats (40 studies, 62.5%), with males constituting at least 82.8% (53/64) of subjects. The C57BL/6 strain (17 studies) was predominant in mouse models due to its stable genetic background and suitability for metabolic studies. In contrast, rat studies primarily used Sprague-Dawley (SD, 23 studies) and Wistar (7 studies) strains, favored for their larger organ size and surgical feasibility.
Disease models
The animal models focused on HIRI (64 studies), with the following distribution: Isolated HIRI: 81.3% (52/64), HIRI + partial hepatectomy (PH): 12.5% (8/64), HIRI + PH + orthotopic liver transplantation: 1.6% (1/64), HIRI + hypoxia/reoxygenation (H/R): 4.7% (3/64). Among HIRI models, 51.6% (33/64) adopted the 70% liver ischemia standard.
HIRI models
There were significant differences in ischemia and reperfusion times in the model (Table 1). The ischemic time was most common at 60 minutes (48.4%,31/64), followed by 90 minutes (28.1%,18/64), ranging from 15 to 90 minutes. The reperfusion time was mainly 6 hours (40.6%,26/64) and 24 hours (51.6%,33/64), and in some studies, it was extended to 48 hours, 72 hours, or even 3 months. Chinese studies mostly adopt 60-minute ischemia + 6/24 hour reperfusion, while Japanese studies tend to favor short-term ischemia (15-45 minutes) and long-term reperfusion (1 day to several days).
Intervention characteristics
Type and source of stem cells
This study systematically analyzed the characteristics of stem cells type and their sources used for treating HIRI (Table 1). The results demonstrated that MSCs served as the primary therapeutic option (96.9%), with bone marrow-derived (BM-MSCs), adipose-derived (ADSCs), and umbilical cord-derived MSCs (UC-MSCs) being the most prevalent subtypes. Notably, approximately 25% (16/64) of the studies employed genetically modified or pharmacologically combined engineered stem cells to enhance therapeutic efficacy. Regarding cells origin, human-derived stem cells predominated in clinical applications, while rodent-derived cells were primarily utilized in preclinical investigations.
Stem cells and exosomes/vesicles modification
This meta-analysis evaluated 16 studies involving stem cells modification and 9 involving stem cell-derived exosome/vesicle modification for the treatment of HIRI. Current modification strategies can be broadly classified into four categories: (1) genetic engineering, including viral vector-mediated overexpression of protective genes (e.g., CD47, HO-1),33^,^36^,^40^,^47^,^67^,^69 miRNA modulation through gene transfection,70 and surface marker-based selection to enrich specific MSC subpopulations;37 (2) pharmacological combination therapies, involving immunosuppressants,34^,^42 active components of traditional Chinese medicine,38^,^64^,^72 anesthetics,45 cytokines,35^,^41 and metabolic regulators65 (3) physicochemical preconditioning, such as hypothermic culture,43 spheroid culture,44 heat shock,46 phosphatidylserine-mediated membrane modification,66 and hepatocyte-like cell-derived exosomes;71 and (4) nanomaterial composites, including nanozyme loading,39 biomembrane-nanocarrier hybrid systems,32 and exosome-drug complexes.68 These modifications enhanced targeting specificity, anti-inflammatory capacity, and cell survival. For instance, CD47 overexpression inhibiting pyroptosis and HO-1 overexpression enhancing anti-inflammatory efficacy.
Stem cells administration: route, dosage, and timing
Administration routes
The included studies employed diverse stem cells delivery methods, primarily intravenous (66%, 42/64) and intraperitoneal routes (Table 1). Intravenous administration included the tail vein (33 studies), peripheral veins,6^,^42^,^55 penile vein,19^,^30 inferior vena cava,60–62 and femoral vein,43 facilitating systemic distribution. Hepatic-targeted delivery (22%, 14/64) was achieved via portal vein (13 studies) and hepatic vein39 injections. Intraperitoneal administration27^,^37^,^44^,^72 was mainly used for modified stem cells (e.g., PDGFRα/Sca-1-sorted MSCs, spheroid-cultured UC-MSCs, glycyrrhetinic acid (GA)-modified MSC-exosomes), whereas localized injections (intrasplenic14 and intrahepatic13) were applied for adipose-derived stem cells (ADSCs). Notably, genetically modified stem cells (e.g., CD47-overexpressing MSCs, baicalin-combined BMSCs) predominantly utilized intravenous routes, whereas exosome-based therapies mainly administered tail vein delivery (64%, 16/25).
Stem cells dose
Stem cells dosage varied depending on cell type, animal model and modification status (Table 1): Unmodified stem cells: The typical dosage is 1 × 10^5^ to 3 × 10^6^ cells/animals, most commonly 1 × 10^6^ cells/mouse; Some studies used higher doses (e.g., 1 × 10^9^ cells for cold-treated ADSCs). Modified stem cells: The dose of engineered MSCs (e.g., SOD2-BMMSCs, TNF-α-pretreated BMSCs) is mostly 1–2 × 10^6^ cells, whereas exosome therapy required lower doses (such as 30–100 μg protein or 10^10^–10^12^ particles). Exosome/vesicle therapy: The dosage is usually standardized according to the protein content (such as 50 μg BMSC-EVs) or the number of particles. Modified exosomes (such as EVs carrying GAS6) may require a higher concentration (2 × 10^10^ particles).
Administration time
Interventions were classified based on timing relative to HIRI: Pretreatment (before ischemia, 25%, 16/64 studies), using strategies such as mild hypothermia combined ADMSCs, CD47-overexpressed MSCs, HO-1 BMMSCs. Interferon-γ (MenSCs) to enhance homing ability; Perioperative period (during HIRI): 22% (14/64) of the studies administered drugs (such as BM-MSCs, ADSCs) at the beginning of reperfusion to exert immediate anti-inflammatory effects; Delayed intervention (after HIRI): 42% (27/64) of the studies administered the drug several hours to several weeks after injury, with a focus on liver tissue regeneration and repair. Modified stem cells were often used in delayed regimens, whereas exosome therapies were predominantly administered perioperatively.
Stem cells surface markers
Stem cells express characteristic surface markers: the core positive markers are CD90, CD73 and CD105 constitute the core triad markers of mesenchymal stem cells; Genetically modified stem cells retain these core markers while exhibiting modification-specific markers (e.g., CD29/CD90 in HO-1-BMMSCs).
Outcome index
Liver function
MSCs markedly improved hepatic function following HIRI, as evidenced by reductions in serum biochemical markers (ALT, AST, LDH, and ALP). Across 64 included studies, 81% (52/64) demonstrated that MSCs and their exosomes effectively suppressed HIRI-induced elevation in liver enzymes and preserved hepatocyte functional homeostasis (Supplementary Table S2), highlighting their robust hepatoprotective effects.
Pathological features
Histopathological analyses consistently showed that MSCs alleviated HIRI-induced hepatic tissue injury. Zhang et al.53 reported improvements in ultrastructural hepatocyte injuries (e.g., nuclear condensation) and increased mitochondrial biogenesis. Other studies6^,^35^,^69 observed decreased Suzuki’s score. While kartal et al.13^,^21 noted attenuation of portal inflammation, hepatocyte edema, cytoplasmic deformation and coagulative necrosis. Additionally, Zare et al.18^,^26^,^28^,^33^,^35^,^38^,^54^,^55^,^61^,^62^,^68^,^71 reported reduced hepatocyte congestion, hepatocyte vacuolation, necrosis and inflammatory infiltration. Ultimately, multiple studies48^,^49 have shown that MSCs can significantly reduce the area of necrotic regions and maintain structural integrity; It simultaneously inhibits end-stage lesions such as coagulative necrosis13 and fibrosis.24^,^58
Oxidative stress and inflammation regulation
Meta-analysis of the included studies revealed that MSCs exert protective effects through the following key mechanisms: (1) Attenuation of oxidative stress7^,^16^,^38–42^,^53^,^59^,^62^,^63^,^65 through ROS scavenging,14^,^71 inhibition of mitochondrial fission,12 restoration of mitochondrial function,32 upregulation of PINK1-mediated mitophagy,6 and reduction of lipid peroxidation;51 (2) Modulation of inflammatory responses7^,^16^,^20^,^34^,^52^,^58^,^67 through downregulation of pro-inflammatory cytokines (TNF-α, IL-6, IL-1β),17^,^23^,^26^,^36^,^37^,^44^,^54^,^55^,^66^,^69^,^72 promotion of anti-inflammatory IL-10 secretion,37 reduction of neutrophil infiltration,21^,^26^,^59 regulation of Treg/Th17 balance,56^,^64 and reprogramming of intrahepatic transcriptional profiles of inflammation-related genes post-IRI,60 including downregulation of inflammatory genes (e.g., TNFA and IL1RN27)
Cell apoptosis and regeneration
MSCs improve liver ischemia-reperfusion injury through a dual mechanism: (1) Anti-apoptotic, as they significantly suppresses apoptosis6^,^7^,^11^,^18^,^20^,^32–36^,^38–40^,^44^,^48^,^49^,^53^,^55^,^57^,^58^,^62^,^63^,^66–71 by down-regulating pro-apoptotic proteins(Caspase-3 and BAX), upregulating anti-apoptotic protein Bcl-2 and reduce TUNEL-positive cells; and (2) Pro-regenerative, as they promotes hepatocyte regeneration20^,^22^,^27^,^29^,^31^,^47^,^50^,^52^,^60^,^61 by increasing PCNA-positive cells,28 elevating mitotic index30 and enhancing hepatocyte growth factor (HGF) expression.24 Additionally, inhibition of α-SMA expression24 mitigates progression toward liver fibrosis.
SYRCLE’s risk of bias tool
The SYRCLE-based risk of bias assessment for all 64 included studies is summarized in Figure 2. Overall, 45.3% of studies reported random sequence generation, with four studies specifically detailing their randomization methods.33^,^45^,^48^,^49 Baseline characteristics indicated a low risk of bias. However, none of articles described randomization of animal housing or allocation concealment, raising concerns about potential selection bias. One study had incomplete outcome data.25No evidence of selective outcome reporting or other systematic biases was identified. Notably, most studies inadequately reported key methodological elements, particularly randomization and blinding procedures.
The methodological quality of each animal study was examined through SYRCLE’s RoB tool.
Meta-analysis
A meta-analysis of 32 eligible studies (41 groups) using 70% hepatic ischemia-reperfusion injury (HIRI) models, evaluated the following outcome measures: ALT, AST, Suzuki’s score, necrotic area ratio per visual field, serum TNF-α levels, and related proteins (Bcl-2, BAX, c-Caspase 3). Studies were stratified by ischemia duration into I45min (1 group), I60min (29 groups), I80min (1 group), and I90min (10 groups). Given the sample size distribution, the analysis primarily focused on the I60min and I90min subgroups to ensure statistical power.
In 70%HIRI rodent models with 60-minute ischemia, MSCs significantly reducted ALT and AST compared with controls (ALT: SMD = 3.49, 95% CI: 2.80–4.17, P < .00001; AST: SMD = 3.86, 95% CI: 2.95-4.77, P<.00001) (Figures 3 and 4). High heterogeneity indices (ALT: I^2^ = 62%; AST: I^2^ = 76%) reflected variability in stem cells sources, preparation, and administration. ALT showed marked heterogeneity at 3 h (I^2^ = 78%) and 24 h (I^2^ = 60%) reperfusion timepoints, while AST showed heterogeneity at 3 h (I^2^ = 79%), 6 h (I^2^ = 80%), 12 h (I^2^ = 86%), and 24 h (I^2^ = 78%). MSCs also reduced Suzuki’s score (SMD = 3.12, 95% CI: 2.26-3.98, P<.00001, I^2^ = 52%), necrotic area ratio (SMD = 3.56, 95% CI: 2.49-4.62, P<.00001), serum TNF-α (SMD = 2.83, 95% CI: 1.94-3.72, P<.00001), and c-Caspase 3 expression (SMD = 2.70, 95% CI: 1.45-3.95, P<.0001) ([Figures 5–8](#szaf078-F5 szaf078-F6 szaf078-F7 szaf078-F8)). Funnel plots indicated no publication bias (Supplementary Figure S1).
*Forest plots showed that in 29 studies with 70%HIRI and 60 minutes of ischemia time, the liver function marker ALT was significantly reduced after stem cell intervention. Stands for exosomes/vesicles or exosomes/vesicles modification.
*Forest plots showed that in 28 studies with 70%HIRI and 60 minutes of ischemia time, the liver function marker AST was significantly reduced after stem cell intervention. Stands for exosomes/vesicles or exosomes/vesicles modification.
*Forest plots showed that in 15 studies with 70%HIRI and 60 minutes of ischemia time, the Suzuki’s score was significantly reduced after stem cell intervention. Stands for exosomes/vesicles or exosomes/vesicles modification.
*Forest plots showed that in 7 studies with 70%HIRI and 60 minutes of ischemia time, the necrotic area ratio per visual field was significantly reduced after stem cell intervention. Stands for exosomes/vesicles or exosomes/vesicles modification.
*Forest plots showed that in 9 studies with 70%HIRI and 60 minutes of ischemia time, the Serum TNF-α was significantly reduced after stem cell intervention. Stands for exosomes/vesicles or exosomes/vesicles modification.
*Forest plots showed that in 11 studies with 70%HIRI and 60 minutes of ischemia time, the c-Caspase 3 expression was significantly reduced after stem cell intervention. Stands for exosomes/vesicles or exosomes/vesicles modification.
Similarly, in 70%HIRI models with 90-minute ischemia, MSCs significantly reduced ALT (SMD = 4.09, 95% CI: 2.59–5.59, P < .00001, I^2^ = 75%) and AST (SMD = 3.78, 95% CI: 3.02–4.53, P<.00001, I^2^ = 19%) (Supplementary Figure S2). ALT showed high heterogeneity at 6 h (I^2^ = 81%) and 24 h (I^2^ = 62%). Funnel plots indicated no publication bias.
Discussion
Hepatic ischemia-reperfusion injury (HIRI) is a common and severe complication encountered during hepatic surgeries such as hepatectomy and liver transplantation. Although timely restoration of blood flow during surgery can partially mitigate ischemic damage, the reperfusion process itself paradoxically exacerbates hepatic injury by triggering oxidative stress, inflammatory cascades, and cell death, potentially leading to irreversible massive necrosis and severely compromising postoperative recovery and long-term outcomes. Currently, no specific and effective strategies are available for the prevention or treatment of HIRI in clinical practice. In recent years, therapeutic approaches based on MSCs and their derivatives (such as extracellular vesicles and conditioned media) have shown remarkable potential in preclinical studies, not only significantly reducing serum liver enzyme levels and ameliorating histopathological injury, but also exerting hepatoprotective effects through multiple synergistic mechanisms-including modulation of inflammatory responses, inhibition of apoptosis, attenuation of oxidative stress, and promotion of tissue regeneration. With the continuous advancement of stem cells engineering techniques and pharmacological preconditioning strategies, the targeting specificity and therapeutic efficacy of MSC-based therapies have been further enhanced, providing new avenues for their clinical translation. By combining quantitative synthesis with qualitative appraisal, our study not only confirms the therapeutic benefits but also delineates key mechanistic insights and translational challenges that will shape the next stage of HIRI research.
The efficacy of MSCs for HIRI: Insights from a meta-analysis
Our meta-analysis demonstrated that MSCs exert a significant therapeutic effect in alleviating HIRI, as evidenced by a marked reduction in serum transaminase levels (ALT and AST) and substantial improvements in histopathological injury severity. Specifically, the observed improvement in liver function indicators (60-minute group: ALT SMD = 3.49, AST SMD = 3.86) 90-minute group: ALT SMD = 4.09, AST SMD= 3.78) suggests a robust hepatoprotective effect. This finding aligns with previous studies reporting that MSCs restore liver function primarily through paracrine mechanisms that modulate the local microenvironment.55 Importantly, this protective efficacy was consistently observed across different ischemic durations (60 vs. 90 minutes) and reperfusion time points (3-24 hours), implying that MSCs may confer protection through multi-target and temporally sustained mechanisms rather than time-restricted actions.
Beyond biochemical indicators, histopathological and anti-apoptotic improvements further corroborate the therapeutic potential of MSCs. The significant reductions in Suzuki scores (SMD = 3.12) and necrotic area ratios (SMD = 3.56) indicate MSCs’ ability to directly mitigate parenchymal damage, while downregulation of apoptotic markers (e.g., c-Caspase 3 downregulation: SMD = 2.70) reveals their intrinsic anti-apoptotic mechanisms. Particularly noteworthy is the synergistic effect between inflammation suppression (e.g., TNF-α reduction: SMD = 2.83) and apoptosis inhibition, potentially mediated by the concurrent blockade of NF-κB45 and MAPK11 signaling pathways. This dual-pathway regulation underscores MSCs’ capacity to orchestrate a coordinated cytoprotective response, providing a mechanistic basis for designing future combination therapies targeting both inflammatory and apoptotic cascades in HIRI.
These findings provide pivotal insights with far-reaching implications for the field of HIRI research. While conventional strategies primarily aim to limit ischemic injury through surgical or pharmacological preconditioning, MSCs-based therapies offer a paradigm shift by actively promoting tissue repair and regeneration in addition to injury mitigation. This positions MSCs as a promising platform for precision regenerative therapy in liver surgery and transplantation. Moreover, the consistent efficacy across diverse experimental settings suggests a degree of robustness that strengthens the rationale for clinical translation.
However, the substantial heterogeneity observed across studies (I^2^ = 52-86%) highlights critical standardization deficiencies in current protocols. Variations in administration routes (e.g., intravenous vs. portal vein injection), dosage regimens (1 × 10^5^-1 × 10^9^ cells), discrepancies in cell sources (e.g., MSCs vs. exosomes) and preconditioning strategies (e.g., HSP-MSCs46) likely contribute to effect size variability. For instance, high-dose interventions (1 × 10^9^ ADMSCs43 or 3 × 10^9^ HLSC-EVs54) appear to enhance efficacy via tissue integrity preservation, transaminase suppression, and inflammatory cytokine modulation, while exosome-based therapies can achieve comparable therapeutic outcomes at significantly lower doses (100 μg),53^,^71 highlighting the concentration of bioactive components as a potential optimization target. Additionally, the temporal dynamics of reperfusion require careful consideration: early-phase improvements (3-6 h) may primarily reflect acute anti-inflammatory effects, whereas sustained benefits (>24 h) likely involve pro-regenerative processes, Extended follow-up periods are therefore warranted to fully assess the long-term tissue remodeling potential of MSCs-based therapies.
Multimodal protective mechanisms of MSCs in HIRI
The pathogenesis of HIRI is a dynamic and multifactorial process. In its early stage, ischemic insult disrupts hepatocyte microenvironmental homeostasis, leading to adenosine triphosphate (ATP) depletion, ion pump dysfunction, and calcium ion (Ca^2+^) imbalance, which cause cellular swelling and structural damage.73 Although subsequent reperfusion restores oxygen delivery, it paradoxically exacerbates cellular injury by inducing a burst of reactive oxygen species (ROS), increasing intracellular Ca^2+^ concentrations, and triggering endoplasmic reticulum (ER) stress, which collectively drive hepatocyte apoptosis and necrosis.74 ER stress, primarily induced by calcium dyshomeostasis and hypoxia-impaired protein synthesis,75 activates the unfolded protein response (UPR) sensors PERK, IRE1, and ATF6, further perturbing cellular proteostasis and promoting autophagy-related gene expression that may aggravate liver injury.76 While these findings have shed light on the pathophysiological underpinnings of HIRI, the complexity and redundancy of its regulatory networks remain incompletely understood, posing a major barrier to the development of targeted therapies.
Within this intricate pathological landscape, stem cell-based therapy emerges as a promising multi-targeted strategy by simultaneously mitigating oxidative stress, attenuating inflammation, inhibiting cell death, and enhancing liver regeneration. Our meta-analysis demonstrated that MSCs can effectively suppress oxidative injury by scavenging ROS, inhibiting excessive mitochondrial fission,12 and up-regulating Pink1-dependent mitophagy.6 Furthermore, paracrine exosomes from MSCs reduce lipid peroxidation by delivering miR-29a-3p69 or activating the AMPK/SIRT1 signaling pathway.65 Beyond their antioxidative roles, MSCs also exert profound immunomodulatory effects: they inhibit neutrophil infiltration,21 downregulate pro-inflammatory cytokines (TNF-α, IL-1β), and enhance the secretion of anti-inflammatory IL-10,37 thereby breaking the self-amplifying cycle between oxidative stress and inflammation that underlies HIRI progression.
Importantly, MSCs does not merely prevent further hepatocyte loss but also actively promotes tissue repair and regeneration. MSCs markedly reduce hepatocyte apoptosis, as evidenced by a decrease in TUNEL-positive cells through modulation of the Bcl-2/Bax axis18 and downregulation of Caspase-3 expression70 (SMD = 2.70, P<.0001). Exosome-mediated miR-25-3p has also been shown to synergistically suppress apoptosis by targeting the p53/PTEN pathway.49 In parallel, MSCs stimulate hepatocyte proliferation via secretion of hepatocyte growth factor (HGF)24 and activation of sphingosine kinase signaling.61 Genetic and engineering advances have further potentiated these therapeutic effects, for example, HO-1-overexpressing bone marrow MSCs attenuate ferroptosis through the AMPK-Nrf2-FTH1 axis,36 while 3D-cultured umbilical cord MSCs enhance reparative efficacy by dampening inflammation-related gene expression.44
Collectively, these findings suggest that MSCs-based therapies act through an integrated network of antioxidative, anti-inflammatory, anti-apoptotic, and regenerative pathways to counteract the multifaceted injury mechanisms of HIRI. This multimodal mode of action underscores their potential to overcome the limitations of conventional single-target strategies, which have thus far shown limited efficacy in clinical settings. More importantly, by elucidating the diverse protective mechanisms mediated by MSCs, these studies not only provide critical mechanistic insights but also lay the groundwork for the rational design of next-generation cell-based or cell-free therapies, thereby facilitating the translation of MSCs from experimental models to clinical practice in the management of HIRI.
Optimization of the therapeutic effect of MSCs strategies
To enhance the therapeutic efficacy of MSCs in HIRI, current research in stem cell engineering primarily focuses on improving their precision, stability, and lesion-targeting capacity, thereby laying the foundation for future clinical translation and application. Gene modification approaches exemplify this trend, for instance, bone marrow-derived MSCs (BM-MSCs) overexpressing superoxide dismutase 2 (SOD2) significantly attenuate liver injury by boosting cellular antioxidant defenses,40 while exosomes enriched with growth arrest-specific 6 (GAS6) promote macrophage efferocytosis via the MerTK-ERK-COX2 signaling cascade.67 Similarly, pharmacological pretreatment of MSCs can fine-tune their immunoregulatory properties; for example, baicalin-preconditioned MSCs enhance the Treg/Th17 balance through activation of the FGF21 axis.64 Beyond biological modification, bioengineering strategies such as nanoenzyme loading39 and phosphatidylserine (PS) surface modification66 have been shown to significantly increase MSC survival, homing efficiency, and therapeutic potency in ischemic liver tissue.
These engineering strategies are not merely incremental enhancements but represent a paradigm shift toward precision-tailored cell therapies for complex liver injuries. By enabling MSCs to more effectively withstand the hostile oxidative and inflammatory microenvironment of the reperfused liver, these modifications amplify their anti-oxidative, anti-inflammatory, and pro-regenerative effects in a synergistic manner. More importantly, they offer a proof-of-concept that MSCs can be rationally redesigned into “smart” therapeutic agents with disease-adaptive functions. However, to facilitate their clinical translation, future work must focus on establishing standardized modification protocols, scalable manufacturing systems, and rigorous safety assessments to prevent unforeseen risks such as oncogenic transformation or immunogenicity.
Exosomes, as the principal paracrine effectors of stem cells, have also garnered increasing interest as a promising cell-free therapeutic alternative. They not only recapitulate most of the beneficial effects of MSCs but also exhibit unique translational advantages, including lower immunogenicity, higher storage stability, and facile bioengineering potential.77 Meta-analysis evidence indicates that exosomes derived from adipose-derived stem cells (ADSCs) suppress hepatic inflammation via the miR-183/ALOX5 axis,50 PS-coated extracellular vesicles (EVs) enhance hepatocyte uptake efficiency,66 and hiPSC-MSC-derived exosomes stimulate liver regeneration through the sphingosine-1-phosphate pathway.61 Nevertheless, emerging data also highlight their inherent heterogeneity: for example, BMSC-EVs containing miR-27b-3p have been shown to induce abnormal hepatocyte proliferation through the Wnt/β-catenin pathway,48 raising safety concerns about unregulated miRNA cargo profiles.
Therefore, while exosome-based therapies hold tremendous translational promise, their clinical development must be accompanied by systematic efforts to standardize their production, dosing, and quality control, as well as to develop targeted delivery systems that can enhance lesion specificity while minimizing off-target risks. Equally crucial is the need for long-term safety evaluations in preclinical and clinical settings to identify potential tumorigenic or immunogenic effects. Addressing these challenges will be pivotal for transforming exosomes from an experimental tool into a reproducible, safe, and scalable therapeutic platform for HIRI.
Taken together, these optimized strategies spanning genetic engineering, pharmacological preconditioning, biomaterial modification, and exosome-based delivery represent a crucial step toward overcoming the current efficacy bottleneck in MSCs for HIRI. They not only deepen our mechanistic understanding of how stem cells interventions can be fine-tuned to meet the pathological complexity of HIRI but also lay a conceptual and technological foundation for the next generation of precision cell and cell-free therapies in liver surgery and transplantation settings.
Limitations and future
Although the research results are encouraging, there are also certain limitations: Model heterogeneity: The ischemic time varies from 15 to 90 minutes, and the experimental animals include mice and rats; Insufficient reporting: Only approximately 45% (29/64) of the studies clearly described the randomization process (according to the SYRCLE bias assessment tool). Additionally, the heterogeneity of MSCs sources represents an important but unresolved issue. Although bone marrow-, adipose-, and umbilical cord-derived MSCs were included, uneven distribution across subtypes and inconsistencies in cell preparation, dosing, and administration made it difficult to determine whether intrinsic biological differences among MSCs sources affected therapeutic efficacy.
Future research should prioritize standardized HIRI models, harmonized MSCs preparation protocols, and head-to-head comparisons of MSCs sources to clarify source-dependent effects. Dose-response relationships, especially for exosomes, require systematic evaluation. Mechanistic studies integrating multi-omics analyses and comprehensive functional assessments beyond ALT/AST are needed. Ultimately, multi-center preclinical and clinical studies will be essential to develop standardized, safe, and clinically translatable MSCs-based therapies for HIRI.
Conclusion
This meta-analysis synthesizes preclinical evidence on MSCs for HIRI, demonstrating its ability to significantly improve liver function parameters and histopathological outcomes through multimodal mechanisms. Engineered MSCs and exosomes offer promising avenues for enhanced therapeutic efficacy. However, inter-model heterogeneity and reporting inconsistencies necessitate resolution through standardized methodologies. Although our findings are derived from rodent studies, they provide essential groundwork for subsequent large-animal and human studies to establish clinically translatable strategies for HIRI treatment. Future research should prioritize dose optimization, mechanistic exploration, and preclinical validation to develop clinically viable cell-based strategies for HIRI management.
Supplementary Material
szaf078_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhai Y , Petrowsky H, Hong JC, et al. Ischaemia-reperfusion injury in liver transplantation—from bench to bedside. Nat Rev Gastroenterol Hepatol. 2013;10:79-89.23229329 10.1038/nrgastro.2012.225PMC 3577927 · doi ↗ · pubmed ↗
- 2Liu Y , Lu T, Zhang C, et al. Activation of YAP attenuates hepatic damage and fibrosis in liver ischemia-reperfusion injury. J Hepatol. 2019;71:719-730.31201834 10.1016/j.jhep.2019.05.029PMC 6773499 · doi ↗ · pubmed ↗
- 3Lu W , Allickson J. Mesenchymal stromal cell therapy: progress to date and future outlook. Mol Ther. 2025;33:2679-2688.39916329 10.1016/j.ymthe.2025.02.003PMC 12172195 · doi ↗ · pubmed ↗
- 4Li P , Ou Q, Shi S, et al. Immunomodulatory properties of mesenchymal stem cells/dental stem cells and their therapeutic applications. Cell Mol Immunol. 2023;20:558-569.36973490 10.1038/s 41423-023-00998-y PMC 10040934 · doi ↗ · pubmed ↗
- 5Hoseini SM , Montazeri F. Cell origin and microenvironment: the players of differentiation capacity in human mesenchymal stem cells. Tissue Cell. 2025;93:102709.39765135 10.1016/j.tice.2024.102709 · doi ↗ · pubmed ↗
- 6Zheng J , Chen L, Lu T, et al. MS Cs ameliorate hepatocellular apoptosis mediated by PINK 1-dependent mitophagy in liver ischemia/reperfusion injury through AMPKα activation. Cell Death Dis. 2020;11:256.32312955 10.1038/s 41419-020-2424-1PMC 7171190 · doi ↗ · pubmed ↗
- 7Sun C-K , Chang C-L, Lin Y-C, et al. Systemic administration of autologous adipose-derived mesenchymal stem cells alleviates hepatic ischemia-reperfusion injury in rats. Crit Care Med. 2012;40:1279-1290.22336724 10.1097/CCM.0b 013e 31823 dae 23 · doi ↗ · pubmed ↗
- 8Ankrum JA , Ong JF, Karp JM. Mesenchymal stem cells: immune evasive, not immune privileged. Nat Biotechnol. 2014;32:252-260.24561556 10.1038/nbt.2816 PMC 4320647 · doi ↗ · pubmed ↗