The Role of Campus as an Urban Multiuse Protected Area in Bird Nocturnal Roosting Habitat Function
Meng He, Bo Li, Wei Du, Chunlan Du

TL;DR
Campus green spaces serve as important nocturnal habitats for birds, with preferences for certain tree species and seasonal changes in roosting behavior.
Contribution
This study reveals how campus green spaces function as nocturnal roosting habitats and identifies specific tree species and seasonal patterns that influence bird behavior.
Findings
Birds prefer Ficus concinna and Ficus virens for roosting due to dense foliage and concealment.
Roosting patterns are more dispersed in summer and concentrated in autumn and winter.
Thermal hotspots indicate higher bird activity in summer compared to other seasons.
Abstract
Campus green spaces are vital components of urban green infrastructure, providing crucial habitats for wildlife, especially birds, in fragmented urban environments. However, their role in supporting nocturnal bird habitats remains underexplored. This study investigates nocturnal bird roosting habitat selection in the green spaces of a university campus in western China. The seasonal surveys were conducted along fixed routes to track the distribution of birds' nocturnal roosts on campus. Infrared hotspots were detected using thermal imaging night‐vision cameras. Roosting tree species were identified, and roosting point heights were estimated visually. The survey results identified eight nocturnal roosting sites and common campus vegetation, which were used for in‐depth analysis of bird roosting habitat characteristics. The study found that: (1) Campus birds preferred Ficus concinna and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
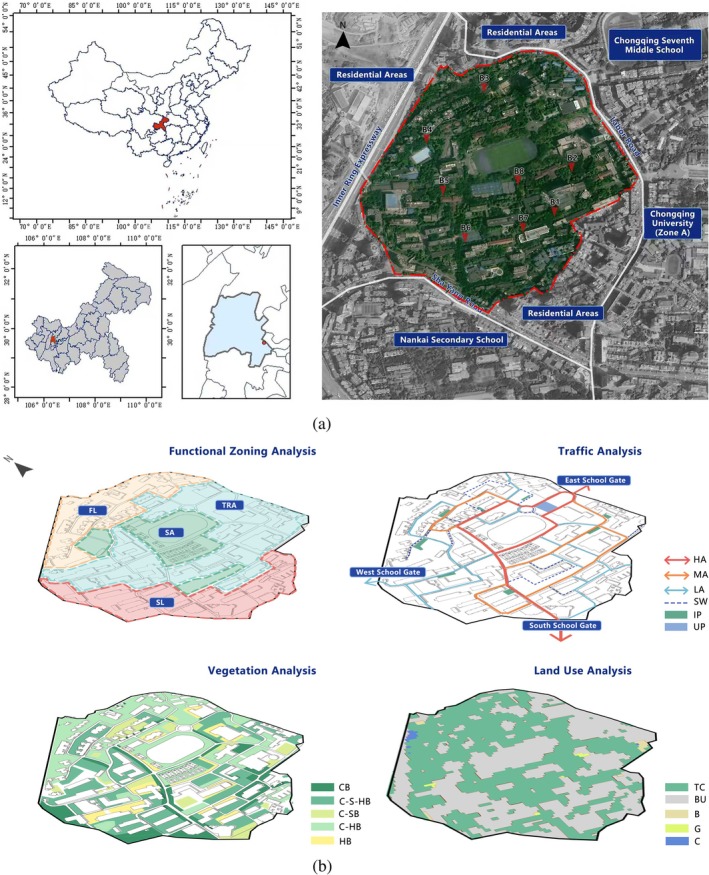 FIGURE 1
FIGURE 1 FIGURE 2
FIGURE 2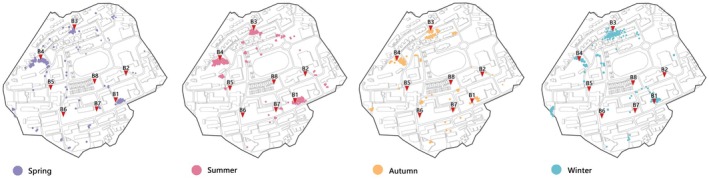 FIGURE 3
FIGURE 3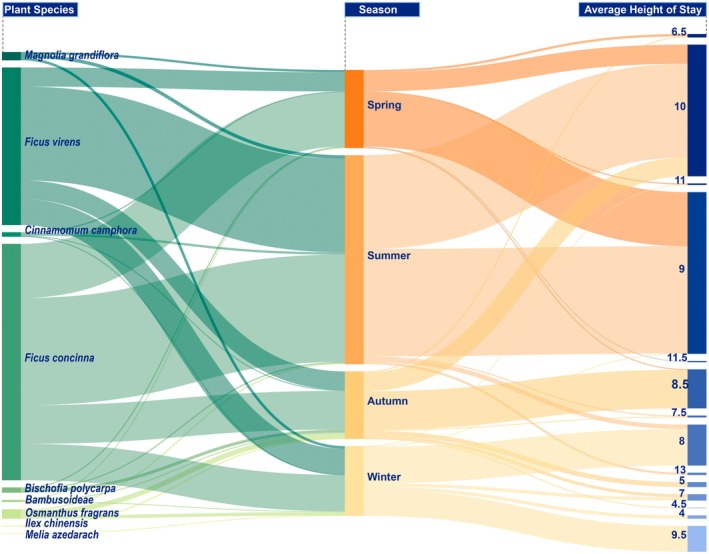 FIGURE 4
FIGURE 4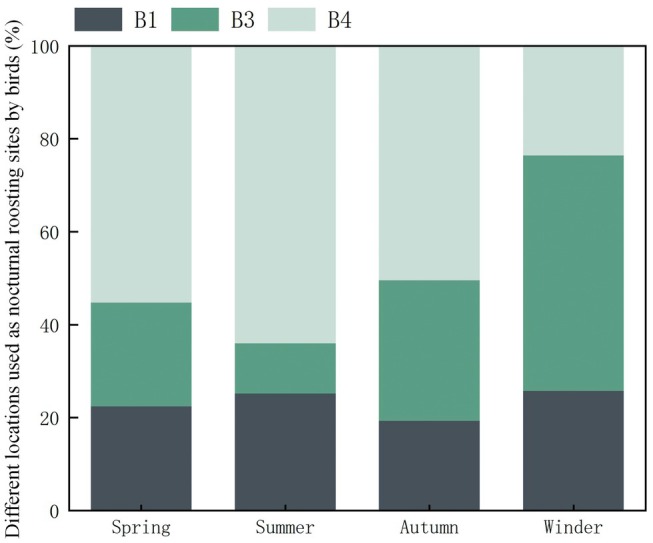 FIGURE 5
FIGURE 5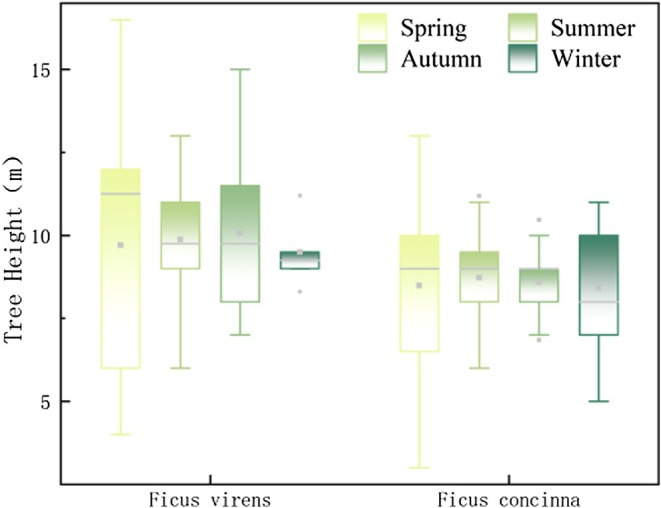 FIGURE 6
FIGURE 6 FIGURE 7
FIGURE 7 FIGURE 8
FIGURE 8| Functional partition | Area/m2 | Frequency of evening activities (times/h·observation point) | Typical activities | Activity intensity | Vegetation structure and type | Includes sample plots |
|---|---|---|---|---|---|---|
| Faculty and staff living area | 90,972 | 133 | Strolling, commuting, walking dogs | Low | Natural greening; arbor + herbaceous vegetation | B3\B4 |
| Student living area | 72,393 | 74 | Commuting, walking | Medium | Functional greening oriented toward circulation; mixed structure of trees and herbs | B6 |
| Teaching and research Area | 158,124 | 452 | Commuting, walking | Medium | Functional greening; mixed tree‐herbaceous structure with guidance‐based layout | B1\B2\B5\B7 |
| Sports and recreation area | 40,030 | 122 | Strolling, Ball games, jumping exercises | High | Dominated by hard pavement and partially covered by arbor vegetation | B8 |
| Plot ID | Area (m2) | Tree species | Tree canopy cover (%) | Shrub cover (%) | Herbaceous cover (%) | Hard surface cover (%) | Canopy density (%) |
|---|---|---|---|---|---|---|---|
| B1 | 1400 |
| 90 | 0 | 50 | 20% | 95 |
| B2 | 1400 |
| 60 | 0 | 0 | 100 | 65 |
| B3 | 1000 |
| 86 | 40 | 0 | 20 | 90 |
| B4 | 1000 |
| 69 | 40 | 55 | 50 | 75 |
| B5 | 1100 |
| 40 | 30 | 0 | 40 | 47 |
| B6 | 1100 |
| 80 | 75 | 0 | 68 | 85 |
| B7 | 1100 |
| 40 | 8 | 37 | 55 | 40 |
| B8 | 1100 |
| 90 | 10 | 50 | 40 | 94 |
| Season | Site | Tree species | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B1 | B3 | B4 | Total | Mean ± SD |
|
|
|
|
|
|
|
|
| Total | Mean ± SD | |
| Spring | 34 | 34 | 84 | 152 | 50.67 ± 28.87 | 0 | 6 | 51 | 3 | 143 | 3 | 0 | 0 | 1 | 207 | 23 ± 47.89 |
| Summer | 128 | 55 | 326 | 509 | 169.67 ± 140.22 | 3 | 10 | 250 | 6 | 286 | 3 | 0 | 0 | 0 | 558 | 62 ± 117.18 |
| Autumn | 21 | 33 | 55 | 109 | 36.33 ± 17.24 | 13 | 0 | 50 | 2 | 103 | 8 | 1 | 0 | 3 | 180 | 20 ± 34.92 |
| Winter | 37 | 73 | 34 | 144 | 48 ± 21.7 | 9 | 7 | 69 | 1 | 97 | 0 | 0 | 1 | 2 | 186 | 20.67 ± 36.16 |
| Source | Likelihood ratio | df |
|
|---|---|---|---|
| Intercept | 11.840 | 1 | 0.001 |
| Species | 30.167 | 8 | < 0.001 |
| Season | 5.019 | 3 | 0.170 |
| Species × Season | 17.018 | 14 | 0.255 |
| Area | Strategies | Optimization module diagram | Goals |
|---|---|---|---|
| Academic and student living |
Retain existing trees; add diverse shrub/herb layers—Prune trees near teaching buildings and study areas—Zone green spaces and walkways |
|
Guide pedestrian flow, reduce disturbance to bird habitats—Minimize bird song interference in study areas |
| Staff residential |
Keep large‐canopy trees; mix evergreen and deciduous—Limit branch pruning—Preserve natural vegetation |
|
Provide shelter and nesting sites—Maintain seasonal resources and roosting stability—Enhance biodiversity |
| Sports |
Increase open areas, separate activity zones from habitats—Use low‐attractiveness plants for birds |
|
Reduce bird disturbance from human activity—Ensure cleanliness of sports facilities |
- —National Natural Science Foundation of China10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsUrban Green Space and Health · Wildlife-Road Interactions and Conservation · Urban Heat Island Mitigation
Introduction
1
Urban green spaces provide essential habitats for wildlife, playing a crucial role in biodiversity conservation (Davis and Glick 1978). However, accelerated urbanization has led to habitat fragmentation and declining habitat quality, posing significant threats to biodiversity (Zhu et al. 2020). Birds, as sensitive indicators of environmental change (Sulaiman et al. 2013), play a key role in assessing urban habitat quality and biodiversity (Callaghan et al. 2018). Consequently, they are frequently studied, including in research on ecological habits (Wang and Chu 2021), distribution characteristics (Sandström et al. 2006), temporal dynamics (Chen et al. 2023), and environmental factors influencing urban bird habitats (Da Silva et al. 2021).
While university campuses primarily serve educational purposes, they are also crucial components of urban green space and hold significant value for bird diversity conservation (Zhang et al. 2018). Cerwinka's (2019) survey at the University of Pennsylvania showed that campuses offer vital stopover habitats for migratory birds and provide resident birds with foraging and nesting sites. Campus green spaces offer birds foraging, refuge, and roosting sites (Rajashekara and Venkatesha 2017; Xiao et al. 2024). Additionally, these spaces provide students, faculty, and the public with direct interactions with nature, while pleasant soundscapes like birdsong can boost positive emotions (Chen et al. 2022).
Current campus management often overlooks the effective protection of biological habitats, especially for birds (Elmqvist et al. 2013). For example, improper plant management and frequent pruning can negatively affect bird diversity (Dearborn and Kark 2010). Such practices may weaken campus green spaces' ecological role in supporting bird diversity. Scientific planning, design, and management are essential to support campus green spaces' ecosystem services in education and bird conservation, but research on this remains limited.
These habitats are essential for bird populations and, to some extent, influence the overall status of bird diversity in the region (Gou et al. 2022). While the term “nocturnal habitat” is sometimes used in the literature, we focus here on the roosting sites used by diurnal birds during the night. In this study, we did not target nocturnal bird species; instead, we surveyed diurnal birds at their nighttime roosts. To avoid confusion, we use the term “birds' nighttime roosts” throughout. A number of scholars have studied the nocturnal habitats of birds and their nocturnal behaviors (Prior and Weatherhead 1991; Smith et al. 2008). For example, Liao et al. (2008) examined the nocturnal behaviors of the Arborophila rufipectus in Laojunshan Nature Reserve, and Wang and Chu (2021) studied how seasonal changes affect the nocturnal roosting behavior of urban cockatoos regarding microhabitat characteristics. Generally, most studies focus on nature reserves or, less often, urban green spaces and are typically species‐specific. However, research on the role of campus green spaces as nocturnal habitats supporting bird biodiversity is scarce.
Understanding nocturnal roosting site selection by birds in campus green spaces is crucial for optimizing green space management and enhancing the functional assessment of urban ecosystems. This study investigates birds' nighttime roosts within the green spaces of Chongqing University to quantify temporal and spatial variation in roost‐site selection, identify key factors influencing roosting site choice, and propose management strategies to enhance the campus's ecological value for biodiversity conservation.
Methods
2
Study Site
2.1
The study area is located within Campus B of Chongqing University, a prominent university in Chongqing, situated in the Shapingba District (29°33′46″ N, 106°28′10″ E, Figure 1). The region experiences an annual average temperature of approximately 17.4°C, with January, the coldest month, averaging 7.8°C, and July, the hottest month, averaging 28.5°C. The area also has a rainy season primarily spanning from May to September. Annual precipitation ranges between 1, 000 mm and 1, 300 mm. Campus B encompasses an area of roughly 33.556 ha and is endowed with rich green spaces, predominantly characterized by tree species such as Ficus virens, Cinnamomum camphora , and Ficus concinna. The overall greenery coverage rate reaches 54.8%. According to functional distinctions, the campus is divided into four main areas: the Faculty and Staff Living Area, Student Living Area, Teaching and Research Area, and Sports and Recreation Area (Table 1). Eight survey points (B1–B8) were established across four functional zones, each with distinct habitat and disturbance profiles. Points B1 and B2, located in the core teaching area adjacent to main roads, experience high‐intensity human commuting and feature manicured, functional greening. The northern points B3 and B4 (Faculty Living Area) offer quieter conditions with more natural vegetation. Points B5 and B7, situated at the periphery of the teaching zone, represent a transition from functional to natural greening. The western point B6 (Student Living Area) is characterized by medium‐intensity, circulation‐dominated activity. The southern point B8 (Sports Area) is dominated by extensive hard pavement and high‐intensity sports activities, representing a highly disturbed, open habitat.
Campus use of space and vegetation analysis. (a) Geographic location of the study site in Chongqing, China, and satellite image of the campus with boundary (red line) and eight sampling plots (B1–B8), adjacent to residential areas and schools. (b) Functional zoning (SA, sports area; TRA, teaching and research area; SL, student living area; FL, faculty living area), traffic hierarchy (HA, high‐traffic artery; MA, medium‐traffic artery; LA, local access; SW, sidewalk; IP, internal path; UP, underground passage), vegetation types (AB, arbor–bush; A‐S‐HB, arbor–shrub–herbaceous; A‐SB, arbor–shrub; A‐HB, arbor–herbaceous; HB, herbaceous), and land‐use categories (TC, teaching/cultural; BU, buildings; B, bare land; G, green space; C, circulation).
Data Collection
2.2
Birds generally enter a resting state at night, during which their activity levels significantly decrease. The nocturnal roosting locations are typically established after sunset and remain stable until birds resume activity at sunrise the following day. Identification of roosting trees was primarily conducted shortly after sunset or just before dawn, when birds had settled into their nocturnal resting locations.
Survey Timing and Frequency
2.2.1
Field surveys for this research were conducted from August 2018 to July 2019, encompassing both bird roosting behavior observations and vegetation surveys within selected plots. To systematically capture seasonal differences in bird roosting behavior, surveys were carried out in four representative months: spring (May), summer (August), autumn (November), and winter (January of the following year). Each month, surveys were conducted continuously for three consecutive days. Monitoring began approximately 1 h after sunset and lasted for 2–3 h, a period when birds generally enter and stabilize at their roosting sites. To control for potential inaccuracies arising from nocturnal movement, supplementary observations were occasionally conducted just before dawn.
Survey Routes and Plot Selection
2.2.2
Multiple survey routes were planned along the primary road network on the campus to ensure coverage of different vegetation types and potential bird activity areas. Observers moved slowly along the predetermined routes while systematically scanning the vegetation on both sides. Necessary foot surveys were supplemented for internal green spaces not fully visible from the roads to minimize observational gaps. Areas with conspicuous nocturnal bird aggregations and representative vegetation structures were identified as fixed study plots through initial reconnaissance.
The measurement of vegetation parameters focused on vegetation type and vertical coverage. Within the selected plots, all major tree species were identified in situ during daytime. Identification was based on key morphological characteristics such as leaf morphology and bark texture, with reference to authoritative taxonomic works like Flora of China. The percent cover (%) of the tree layer, shrub layer, and herbaceous layer was visually estimated to quantify the three‐dimensional vegetation structure.
Equipment and Observation Procedures
2.2.3
During data collection, an LM6P thermal imager was employed to scan individual trees and shrubs along each survey route. A night vision device was used to confirm the presence of birds at nighttime roosts (hotspots) (Figure 2). The detailed operational procedures were as follows:
Instruments used in nocturnal surveys. (a) LM6P handheld thermal imager; inset shows canopy thermal “hotspots.” (b) NV400 digital night‐vision device; inset shows the verified outlines/positions of roosting birds under low light.
Multi‐Angle Detection
2.2.3.1
Observations were conducted from a suitable distance (approximately 5–10 m or adjusted according to field conditions), scanning individual tree crowns from multiple angles, from bottom to top, to minimize missed detections due to foliage obstruction. Potential roosting bird locations were identified by real‐time “hotspots” on the thermal imaging screen.
Hotspot Verification
2.2.3.2
Upon detecting a hotspot, a night vision device was immediately used to confirm the hotspot's outline and activity. In cases of uncertainty, brief illumination with a flashlight was employed to further verify the presence of birds while minimizing disturbance. Subsequently, the tree species or specific branch location associated with the hotspot was recorded.
Recording Standards
2.2.3.3
Confirmed bird roosting locations were documented, including time, specific location, tree species or vegetation type, estimated height, and the number of bird hotspots. Due to operational challenges posed by nighttime conditions and the specific nature of nocturnal roosting (e.g., high numbers of roosting birds, dense foliage), direct measurement of roosting height was difficult. Therefore, the heights of roosting branches were visually estimated (Hou et al. 2012). Although visual estimation may introduce potential errors, consistency was ensured by repeated calibrations and observations conducted by the same researcher, providing valid reference data for studying spatial patterns of bird nocturnal roosting.
Data Analysis
2.3
This study employed multiple statistical models to examine the effects of various factors on roosting behavior, complemented by visualization techniques to interpret and present the results. First, a one‐way analysis of variance (ANOVA) was used to test for seasonal differences in roosting height; as the assumption of homogeneity of variances was violated (Levene's test, p < 0.05), Tamhane's T2 post hoc test, which does not assume equal variances, was applied for pairwise comparisons between seasons. Second, a generalized linear model (GLM) was employed to analyze the count data of roosting hotspots. Due to detected overdispersion in an initial Poisson model, a negative binomial distribution was ultimately selected for the final model, which included the main effects of tree species and season, as well as their interaction term. Furthermore, to intuitively reveal spatial patterns and statistical trends, the spatial distribution of roosting sites was mapped using ArcGIS, and various statistical graphs, including boxplots, Sankey diagrams, and stacked bar charts, were generated using OriginPro.
Results
3
Nocturnal Observations and Spatial Distribution of Campus Birds
3.1
A total of 1131 nocturnal bird hotspots were recorded in this survey, indicating the nocturnal roosting site preferences of campus bird species (Figure 3). Of these, 914 hotspots (81% of the total) were concentrated in areas B1, B4, and B5. Fewer birds roosted in street trees or other isolated trees within the campus. Seasonal variations significantly influenced the choice of nocturnal roosting sites. In spring, summer, and autumn, area B5 was the primary roosting site, while in winter, area B4 was preferred.
Seasonal spatial distribution of thermal hotspots corresponding to birds' nighttime roosts. The dots represent the number of hotspots for birds, with denser dots representing a higher number of observed hotspots for birds.
Analysis of the characteristics of bird‐preferred nocturnal roosts (Table 2) revealed that these sites were typically pure forests with a “tree‐herb” structure (B1, B3, B4), dominated by tall trees, a high degree of depression, and minimal hard‐surface cover in the understorey.
Plant Species Preferences for Birds' Nighttime Roosts
3.2
A total of nine nocturnal roosting plant species, spanning eight families and eight genera, were recorded in Area B of Chongqing University. These included Ficus concinna, Ficus virens, Magnolia grandiflora , Osmanthus fragrans , Cinnamomum camphora , Bischofia polycarpa, Bambusoideae, Ilex chinensis, and Melia azedarach . Ficus concinna and Ficus virens were the primary choices for nocturnal roosting, attracting a total of 1050 birds hotspots, which represented 93% of all birds recorded (Table 3). Notably, birds roosted at higher elevations on these plants during spring and summer compared to autumn and winter.
To assess the influence of plant species and season on the distribution of nocturnal bird roosting hotspots, we fitted a generalized linear model (GLM). The model showed a significant overall fit, indicating that the predictors jointly affected roosting site selection (Table 4). Plant species had a significant effect on hotspot selection (Likelihood Ratio χ ^2^ = 30.167, df = 8, p < 0.001), consistent with distinct preferences among tree species. These results imply that bird preferences for specific tree species were consistent across seasons.
Interestingly, birds were observed roosting at higher canopy positions during spring and summer compared with autumn and winter (Figure 4). This seasonal shift may reflect changes in microclimatic conditions or predation risk, although such effects were not captured by the species–season interaction in the current model.
Associations between birds' nighttime roosts and roosting plant species. Sankey diagram linking plant species used by birds as nighttime roosts (left) to roosting height (m) (right), the width of the line indicates the number of bird hotspots, and the more hotspots that are recorded, the wider the line is.
Seasonal Differences in the Nocturnal Behavior of Birds
3.3
Seasonal Differences in Site Selection for Birds at Nighttime Roosts
3.3.1
Bird selection for the main nocturnal roosts (B1, B3, and B4) varied seasonally (Figure 5). In spring and summer, B4 was highly preferred, reaching over 80% of selections in summer. In autumn, B4 usage declined while B3 increased to around 40%. By winter, B3 became the main roosting site, attracting 90% of birds, while B1 and B4 showed a marked decrease in selection.
Proportion of birds' nighttime roosts across habitat types by season.
Seasonal Differences in the Height of Nocturnal Roosting Sites for Birds
3.3.2
Analysis of the two primary roosting species, Ficus concinna and Ficus virens, revealed significant seasonal variation in roosting height (Figure 6). One‐way ANOVA indicated a significant main effect of season (F = 2.512, p = 0.060). Due to a violation of the homogeneity of variances assumption (Levene's test, p < 0.05), Tamhane's T2 post hoc test was employed for pairwise comparisons. This analysis showed that the roosting height in winter was significantly lower than in summer (Mean Difference = 1.166, p = 0.032). Although not all pairwise comparisons with winter reached statistical significance, a general trend of lower roosting heights in winter was observed (Table S1).
Bird nocturnal vegetation height box line plot. The vertical axis indicates tree height (M), while the horizontal axis distinguishes the two tree species. The box‐and‐whisker plots illustrate the range and median values of tree heights for each season.
Specifically, the distribution of roosting heights was widest in spring, particularly on Ficus virens, indicating more diverse height selectivity during this season. In contrast, roosting heights were more clustered in summer and autumn, reflecting more consistent height preferences. Thermal imaging observations corroborated these findings (Figure 7), showing that birds were more spatially dispersed in spring and summer, but formed denser clusters in autumn and winter.
Seasonal patterns in nocturnal roosting hotspots. Panels show canopy‐level thermal detections of hotspots (red dots) for (a) spring, (b) summer, (c) autumn, and (d) winter. The observations indicate more dispersed hotspots in spring and summer and more clustered hotspots in autumn and winter.
Discussion
4
Habitat Preferences for Birds at Nighttime Roosts
4.1
Campus green spaces not only provide pleasant living environments for humans but also serve as essential habitats for urban birds. This study reveals distinct seasonal preferences in nocturnal roosting site selection by campus birds. Birds select their roosting sites carefully based on ecological needs and physiological traits, typically demonstrating strong site fidelity once a roost is chosen (Wang and Chu 2021). Across all seasons, approximately 81% of nighttime roost detections were concentrated in three consistently used plots (B1, B3, and B4). Specifically, plot B4, located in the retired faculty residential area with extensive vegetation cover, provided ample shelter and water resources, making it favorable for bird roosting. These results align with Yuan Baodong's findings (Yuan et al. 2018), which reported long‐term site fidelity in Hume's pheasant ( Syrmaticus humiae ). Notably, although B6 and B8 have high canopy cover (Table 2), both sites are dominated by non‐Ficus species, whereas hotspots in our study were concentrated in Ficus‐dominated plots. Comparative analyses (Table 2) suggest that this stable selection is associated with differences in tree height, plant assemblage, and the intensity of human disturbance, indicating that Ficus canopies with dense, horizontally tiered branches may provide more effective nocturnal shelter.
Regarding habitat structure, analyses revealed that preferred nocturnal roosting plots (B1, B3, and B4) had higher canopy coverage formed by trees like Ficus concinna and Ficus virens, creating relatively safe roosting environments. Dense tree crowns offered birds protection and vigilance opportunities by minimizing predation risks and human disturbances. Notably, most birds preferred roosting at the junction of two trees (Figure 8), balancing visibility and ventilation while reducing susceptibility to disturbance.
Schematic diagram of nocturnal roosting for birds. Dotted circles mark observed roosting birds within the canopy; most birds perch on outer branch tips in the upper middle crown, a position that likely reduces predation risk. Diagram is illustrative and not to scale.
Similar conclusions have been reported by Clergeau and Quenot (2007), who noted birds favor densely foliated trees accommodating multiple individuals, thereby enhancing predator defense capabilities. Pan et al. (2019) also found concealment of roosting branches critical for the safety of Copsychus saularis . Additionally, species like silver pheasants (Lophura nycthemera, Shao and Hu 2005) and barn swallows ( Hirundo rustica , Verma 2010) exhibit preferences for dense foliage and concealment. Species‐specific differences also occur, with some birds, such as blue‐eared pheasants ( Crossoptilon auritum ), favoring dense shrub habitats for increased ground‐level safety (Zhang et al. 2023). Consequently, varied tree heights or ground vegetation configurations in campus landscapes may lead to differentiated roosting patterns across bird communities, which accords with our observation that hotspots were concentrated in Ficus‐dominated plots with dense, horizontally tiered crowns.
In summary, this study's statistical analysis and quantified habitat assessments preliminarily confirm that tree‐herbaceous forest structures, high canopy coverage, minimal hardscape cover, and moderate concealment significantly influence nocturnal bird roosting preferences. Furthermore, the intensity of human disturbance also influences roost site distributions; quieter environments, like around plot B4, reduce birds' vigilance and enhance perceived safety. These findings provide empirical support for subsequent exploration of ecological functions and management optimization strategies in campus green spaces. By retaining key trees, maintaining canopy coverage, and controlling nocturnal human activities, campuses can better meet avian requirements for secure roosting habitats.
Seasonal Dynamics of Nocturnal Roosting Behavior
4.2
The study indicated that while birds often demonstrate fidelity to specific roost sites, bird abundance varied significantly across seasons. For example, bird numbers peaked in summer (breeding season), while roosting distributions differed during spring, autumn, and winter.
Seasonal Distribution Patterns of Nocturnal Roosting Locations
4.2.1
High summer temperatures necessitate thermoregulation for birds, causing dispersion into more open, ventilated vegetation areas to reduce heat stress and energy costs related to thermoregulation (Zhao et al. 2023). According to heat‐balance theory, birds select roost sites dynamically to optimize local microclimates and minimize metabolic expenditures (Walsberg 1986). Empirical evidence supports this theory, such as spotted owls ( Strix occidentalis ) preferring sparse canopies with good airflow during hot months to mitigate heat stress (Barrows 1981), and Eurasian magpies (Pica pica, Crosbie et al. 2006) displaying individual roosting patterns correlated positively with temperature, reducing thermal interactions (Crosbie et al. 2006). Compared with typical urban green spaces, university campuses feature higher vegetation coverage and diversity, offering abundant microhabitats and behavioral flexibility crucial for mitigating thermal stress and optimizing resource acquisition (Sanllorente et al. 2023).
As temperatures decrease, birds face physiological pressures for heat retention, often forming communal roosts or selecting highly concealed environments. Studies on brown‐headed cowbirds (Molothrus ater, Francis 1976) indicate communal roosting reduces heat loss and enhances predator vigilance. Although oriental magpie robins (Pan et al. 2019) typically roost alone, occasional communal roosting occurs under low temperatures. Hence, clustered roost distributions during autumn and winter on campus green spaces represent adaptive responses to cold conditions.
Seasonal Adjustments in Tree Species Preferences and Roosting Height
4.2.2
Trees like Ficus virens and Ficus concinna attract birds to varying degrees seasonally, influenced by leaf shedding cycles and vegetation density. For example, leaf shedding in spring reduces canopy density at plot B3, temporarily diminishing its attractiveness until foliage regeneration in winter restores its status as a hotspot.
Regarding roosting height, birds generally prefer relatively elevated positions but avoid the highest canopy points (Sedgeley 2001), balancing microclimate conditions and predator avoidance (Harris and McMurry 2024). Observations indicate average roosting heights were higher in spring, summer, and autumn compared to winter (Figure 5), potentially related to seasonal temperature variations and dynamic canopy structures. Kelty and Lustick (1977) observed similar patterns, with birds exploiting upper canopies for cooling in warmer seasons and lower branches during cold months to reduce heat loss.
In conclusion, seasonal dynamics in nocturnal roosting behavior are shaped by multiple ecological factors, including thermal balance, reproductive scheduling, and vegetation structure. Because our nocturnal protocol focused on thermal detections in trees and shrubs, ground‐dwelling or low‐roosting species (e.g., pheasants) may have been underdetected. Future work should incorporate ground‐level and understory assessments to better capture these taxa. Campus green‐space management strategies that account for these seasonal patterns by enhancing tree and shrub‐layer configuration could substantially strengthen the ecological services of campus environments, particularly roost provision and avian biodiversity conservation.
Human Disturbance and Birds at Nighttime Roosts Behavior
4.3
Despite environmental disturbances like artificial lighting at plot B3 and periodic academic bell sounds near plot B4, no significant impacts on bird roosting were detected. Previous studies indicate urban green spaces, often characterized by simple vegetation structures and limited diversity, force birds into restricted habitats where they adapt gradually to disturbances such as noise and lighting (Liu et al. 2023). Hence, noise and illumination are typically secondary disturbances for roosting birds. For instance, oriental magpie robins tolerate urban noise levels exceeding 40 dB (Pan et al. 2019), while rainbow lorikeets (Trichoglossus moluccanus, Daoud‐Opit and Jones 2016) prefer brightly illuminated areas, exemplifying urban bird resilience to disturbances.
Nevertheless, bird tolerance to disturbances has limits (Gorenzel and Salmon 1995). Despite suitable habitats, plots B7 and B8 near sports facilities showed significantly reduced roosting bird numbers due to persistent high human activity and late‐night illumination. This underscores that continuous or intense human disturbances negatively influence bird roosting behavior.
Future research should quantitatively assess disturbance thresholds through noise monitoring and human traffic analyses. Moreover, potential understudied disturbances like vehicle traffic and wildlife interactions require further exploration to comprehensively evaluate impacts on avian roosting behavior.
A Green Space Design Strategy for University Campuses Based on the Conservation of Nocturnal Roosting Sites for Birds
4.4
University campuses serve not only as vital spaces for study, research, and residence but also as essential ecological areas where humans and nature coexist harmoniously. Studies indicate that engaging in physical activities in green environments can significantly alleviate depression and stress (van den Bosch and Meyer‐Lindenberg 2019), particularly on campuses where academic and work pressures are prevalent. Thus, strategic planning and placement of green spaces across campus can help alleviate psychological stress. Additionally, campuses support substantial biodiversity, particularly in bird species richness (Ahir and Singh 2024). Campus green spaces provide essential bird habitats and play a key role in sustaining bird populations (Table S2, Zhang et al. 2023). However, some urban‐tolerant species frequently use campus roosts and may pose management challenges. Prolonged communal roosting can lead to guano accumulation that soils vehicles along roads and in parking areas, and vocal activity can disrupt study environments (Barcan and Johnston 2024). In response, this study proposes green‐space optimization strategies (Table 5) that balance human use with habitat preservation and foster a symbiotic campus ecosystem. Where intervention is necessary, actions should prioritize nonlethal, habitat‐based measures, such as periodic pruning scheduled outside the breeding season, selective crown lifting near entrances and roadways to discourage large aggregations, and routine cleaning, rather than removals that could cause avoidable ecological harm.
For routine maintenance, it is recommended to retain perching branches of various sizes and heights to support birds' nocturnal concealment and roosting needs. Meanwhile, low branches near campus roads, parking areas, and entrances should be moderately pruned to limit bird congregation and reduce guano accumulation. Additionally, faculty and students from related disciplines are encouraged to participate in ecological monitoring and maintenance programs to assess vegetation growth and bird activity on campus, ensuring green spaces meet bird roosting needs while maintaining a clean campus environment. This process also provides valuable educational opportunities for students and promotes the harmonious coexistence of humans and nature.
Conclusions
5
Campus green spaces, as crucial ecological areas in urban settings, not only create a pleasant environment for learning and living but also serve as essential bird habitats, offering key areas for foraging, breeding, and predator avoidance. Research indicates that birds' choice of nocturnal roosts is stable and largely influenced by habitat characteristics; while roosting behaviors vary significantly with the seasons, reflecting birds' adaptability to environmental changes. Campus green spaces not only provide suitable habitats for birds but also significantly enhance bird diversity and support campus and urban biodiversity. Therefore, this study can help optimize the ecological functions of campus green spaces and provide a scientific basis for urban biodiversity conservation, supporting the healthy and sustainable development of urban ecosystems.
Author Contributions
Meng He: conceptualization (equal), data curation (equal), investigation (equal), methodology (equal), writing – original draft (equal). Bo Li: funding acquisition (equal), investigation (equal), methodology (equal), writing – review and editing (equal). Wei Du: investigation (equal), visualization (equal). Chunlan Du: funding acquisition (equal), supervision (equal).
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 52238003 and 52078074).
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Table S1: Pairwise seasonal comparisons of roosting height (m) for Ficus concinna and F. virens (Tamhane's T2, unequal variances). Table S2: Daytime bird checklist for the campus.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahir, A. , and Y. Singh . 2024. “The Diversity and Abundance of Bird Species in Urban Landscape; Barkatullah University Campus, Bhopal, India.” Species 25, no. 75: 1–9. 10.54905/disssi.v 25i 75.e 11s 1633. · doi ↗
- 2Barcan, R. , and J. Johnston . 2024. “Birds as Buddies: The Politics of Sentimentality in the Birds in Backyards (Australia) Facebook Site.” Continuum 38, no. 1: 98–110. 10.1080/10304312.2023.2234106. · doi ↗
- 3Barrows, C. W. 1981. “Roost Selection by Spotted Owls: An Adaptation to Heat Stress.” Condor 83, no. 4: 302–309. 10.2307/1367496. · doi ↗
- 4Callaghan, C. T. , R. E. Major , M. B. Lyons , J. M. Martin , and R. T. Kingsford . 2018. “The Effects of Local and Landscape Habitat Attributes on Bird Diversity in Urban Greenspaces.” Ecosphere 9, no. 7: e 02347. 10.1002/ecs 2.2347. · doi ↗
- 5Cerwinka, C. 2019. “Birds as Ecological Indicators at the University of Pennsylvania (Doctoral dissertation, University of Pennsylvania)”.
- 6Chen, H. , B. Hong , H. Qu , Y. Geng , and M. Su . 2022. “Effects of Acoustic Perception on Outdoor Thermal Comfort in Campus Open Spaces in China's Cold Region.” Buildings 12, no. 10: Article 10. 10.3390/buildings 12101518. · doi ↗
- 7Chen, Y. , M. A. Rasool , S. Hussain , et al. 2023. “Bird Community Structure Is Driven by Urbanization Level, Blue‐Green Infrastructure Configuration and Precision Farming in Taizhou, China.” Science of the Total Environment 859, no. Pt 1: 160096. 10.1016/j.scitotenv.2022.160096.36372169 · doi ↗ · pubmed ↗
- 8Clergeau, P. , and F. Quenot . 2007. “Roost Selection Flexibility of European Starlings Aids Invasion of Urban Landscape.” Landscape and Urban Planning 80, no. 1: 56–62. 10.1016/j.landurbplan.2006.06.002. · doi ↗