Evolutionary dynamics of the proanthocyanidin biosynthesis gene LAR
Maria F. Marin-Recinos, Boas Pucker

TL;DR
This paper studies how the LAR gene, important for plant pigment production, evolved differently across plant species.
Contribution
The study reveals lineage-specific evolutionary patterns and functional divergence in LAR genes across major plant clades.
Findings
LAR genes show multiple independent duplication events and lineage-specific expansions in dicots and gymnosperms.
Differential retention and diversification of LAR1 and LAR2 in dicots suggest functional specialization.
Promoter and synteny analyses indicate regulatory and structural divergence in LAR genes.
Abstract
Leucoanthocyanidin reductase (LAR) is a key enzyme in proanthocyanidin (PAs) biosynthesis, catalyzing the conversion of leucoanthocyanidins to catechins. While early steps in the flavonoid pathway are broadly conserved across plant lineages, increasing evidence demonstrates lineage-specific evolutionary trajectories and functional diversification in its terminal branches, particularly in the case of LAR. To explore the evolutionary dynamics and functional divergence of LAR genes, we conducted large-scale comparative and phylogenetic analyses across major plant clades. The phylogenetic analysis revealed multiple independent duplication events and lineage-specific expansions of LAR lineages, particularly among dicots and gymnosperms. In dicots, LAR1 and LAR2 were differentially retained and diversified, whereas gymnosperm LAR homologs formed early-diverging clades, suggesting an ancient…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
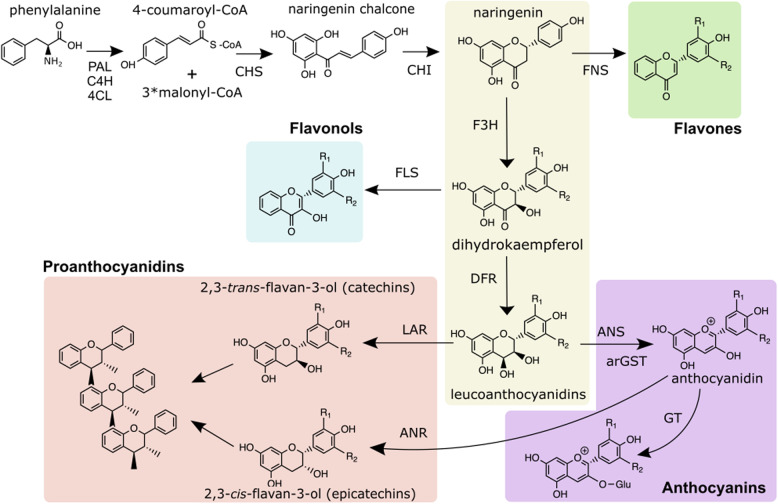 Figure 1
Figure 1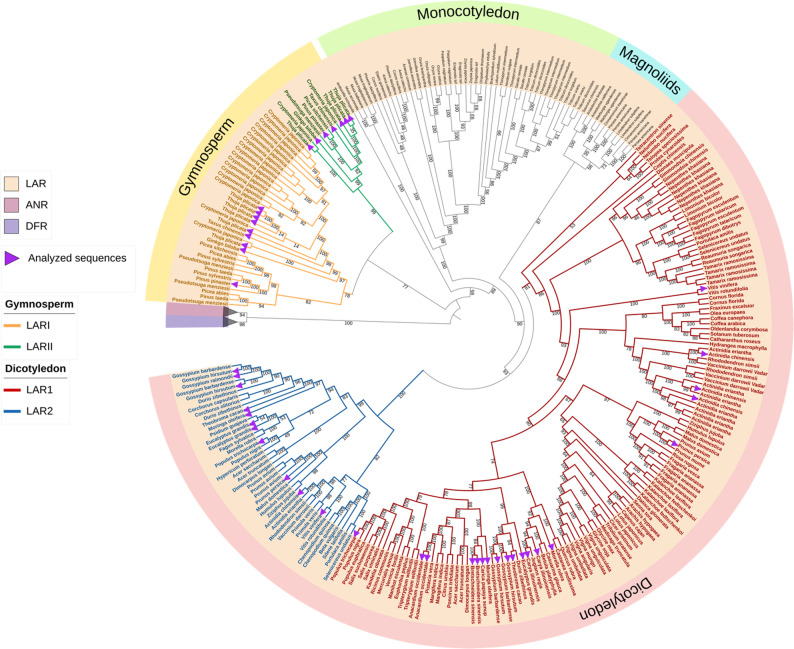 Figure 2
Figure 2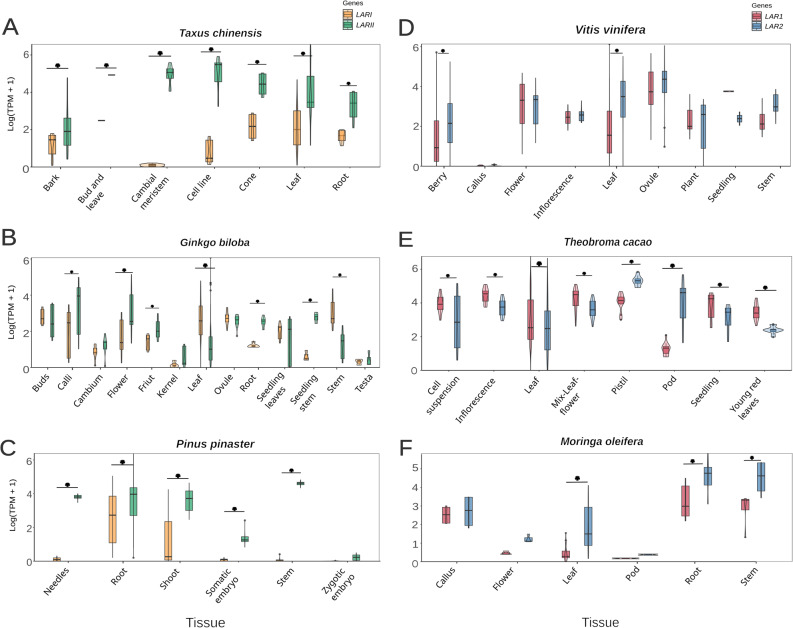 Figure 3
Figure 3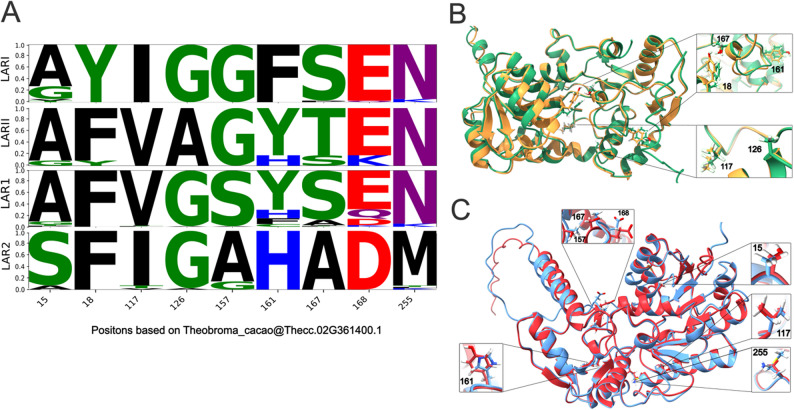 Figure 4
Figure 4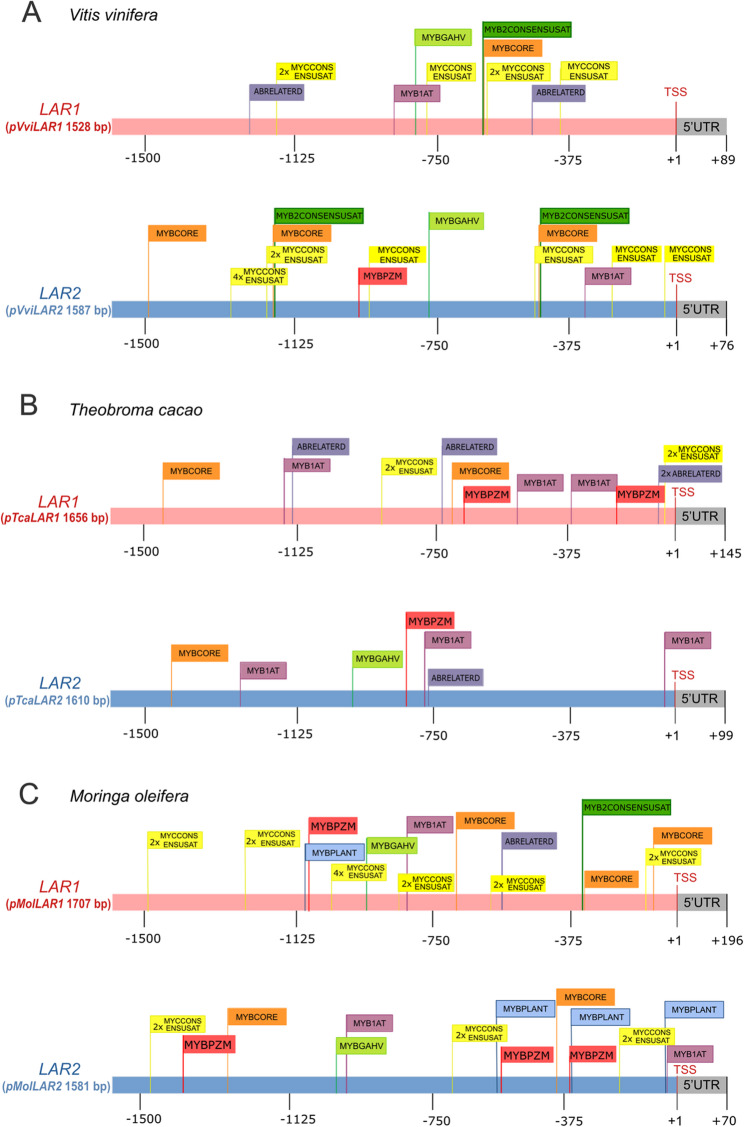 Figure 5
Figure 5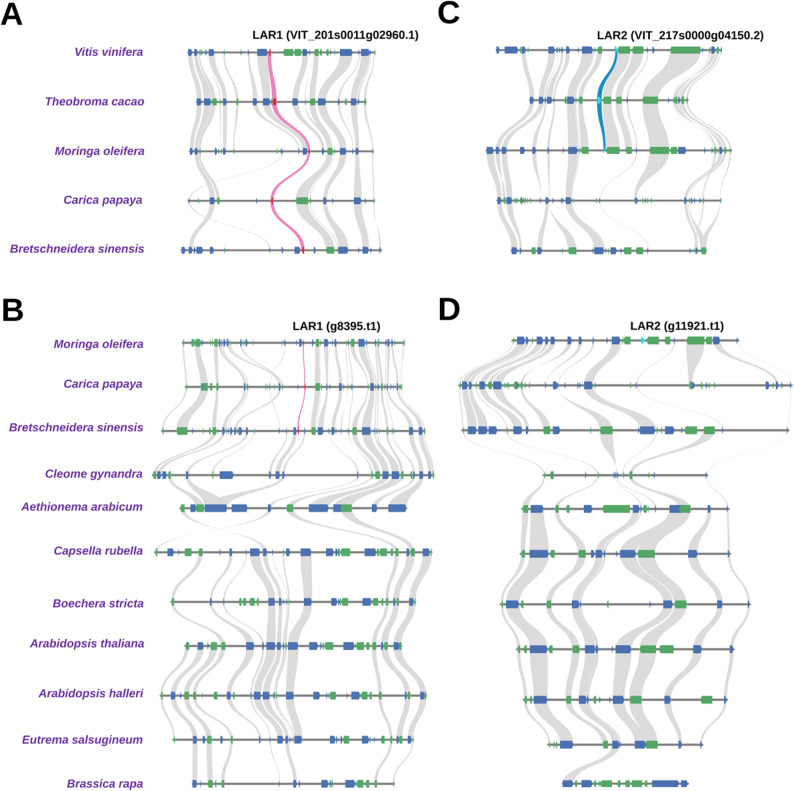 Figure 6
Figure 6- —Rheinische Friedrich-Wilhelms-Universität Bonn (1040)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Gene Expression Analysis · Plant biochemistry and biosynthesis · Horticultural and Viticultural Research
Introduction
Proanthocyanidins (PAs), also referred to as condensed tannins, constitute a group of polyphenolic compounds within the flavonoid biosynthesis pathway. As secondary metabolites, PAs play an important role in plant defense mechanisms against insect herbivores and microbial pathogens. Their contribution to herbivore deterrence is largely attributed to the astringent and bitter taste they confer to various plant tissues, which reduces palatability and impairs digestibility in insect pests [1–3]. In addition, PAs exhibit antimicrobial activity against pathogens, including bacteria [4–6] and fungi [7]. Historically, PAs have been utilized as tanning agents for leather preservation and in the regulation of different flavor qualities and profiles in beverages such as wine, teas, and fruit juices [8, 9]. Furthermore, their antioxidant properties have been reported for their potential health benefits in both human and ruminant animal consumption [10]. In addition to their role in plant defense, PAs have been extensively studied for their impact on seed coat pigmentation [11, 12], influencing seed dispersal [13] and dormancy mechanisms [14], thereby contributing to plant reproductive efficacy [15, 16].
PA synthesis represents one branch of the flavonoid biosynthesis pathway, a process extensively elucidated across a wide range of plant species and notably conserved among diverse taxa [17–19]. The synthesis of PAs and anthocyanins, as competing branches of the pathway, share common precursors which involve a series of enzymatic reactions. The key enzymes involved in PA synthesis start with the enzyme dihydroflavonol 4-reductase (DFR) catalyzing the production of leucoanthocyanidins which are then processed by leucoanthocyanidin reductase (LAR) for the conversion into 2,3-trans-flavan-3-ols, commonly known as the monomer 2,3-trans-(+)-catechin. In a parallel route, anthocyanidin synthase (ANS) converts leucoanthocyanidins to anthocyanidins, which can be reduced by anthocyanidin reductase (ANR) to form 2,3-cis-flavan-3-ols, also known as the monomer 2,3-cis-(-)-epicatechin. These monomeric units then polymerize to form PA oligomers (Fig. 1).
Fig. 1. Schematic representation of the general flavonoid biosynthesis pathway. Enzyme names are abbreviated as follows: PAL - phenylalanine ammonia-lyase, C4H - cinnamic acid 4-hydroxylase, 4CL − 4-coumarate-CoA ligase, CHS - chalcone synthase, CHI - chalcone isomerase, FNS - flavone synthase, F3H - flavanone 3-hydroxylase, FLS - flavonol synthase, DFR - dihydroflavonol 4-reductase, ANS - anthocyanidin synthase, arGST – anthocyanin-related glutathione S-transferase, GT - UDP-dependent anthocyanidin- 3-O-glucosyltransferase, ANR - anthocyanidin reductase
LAR and ANR play a central role in defining PA composition by synthesizing the flavan-3-ol monomers that act as starter and extension units. Canonically, the primary constituents of these polymers are (+)-catechin, produced by LAR, and (-)-epicatechin, synthesized by ANR. These two monomers serve as the basic building blocks in the formation of proanthocyanidin polymers. However, recent studies in Medicago truncatula have reported that the function of LAR extends beyond its traditional role in the synthesis of (+)-catechins [20, 21]. Specifically, LAR has been shown to be responsible for converting 4β-(S-cysteinyl) epicatechin back to epicatechin, a reaction that was previously attributed exclusively to ANR [20, 21]. This dual LAR activity may explain the observed increase in catechin and epicatechin levels under overexpression of LAR, indicating that LAR is crucial for regulating both monomer types that serve as starter and extension units in the polymerization process of PAs [20, 21]. Furthermore, Jun et al. demonstrated that leucoanthocyanidin dioxygenase (LDOX) in M. truncatula plays a role in the formation of epicatechin starter units from catechin, contrasting with the function of ANS in the conversion of leucocyanidin to cyanidin [21]. Meanwhile, ANR is involved in the synthesis of the extension epicatechin units [21].
In addition to M. truncatula, LAR genes have been characterized in multiple economically important plants such as Vitis vinifera [22], Theobroma cacao [23], Gossypium hirsutum [24], Malus domestica [25], Pyrus communis [26], and Camellia sinensis [27], all of which belong to a diverse range of plant families and orders. Interestingly, despite the widespread presence of LAR genes in many plants, it has been known for years that the model plant Arabidopsis thaliana does not harbor a LAR gene [19, 28]. However, A. thaliana is still capable of synthesizing proanthocyanidins, which provide a dark color to its seed coat [12]. Knowledge obtained from studying the transparent testa (tt) and tannin-deficient seed (tds) mutants collection in A. thaliana have been essential in understanding the synthesis, regulation, and transport mechanisms involved in the PA production, even in the absence of a functional LAR gene [12, 19, 29–32]. The regulation of flavonoid biosynthesis, including the PA branch, is controlled by a well-characterized complex of transcription factors (TFs) known as the MBW complex, consisting of MYB, basic helix-loop-helix (bHLH), and WD40 proteins [33]. In A. thaliana, key TFs such as TT2 (R2R3-MYB), TT8 (bHLH), and TTG1 (WD40) regulate the expression of BANYULS (BAN) which encodes for ANR involved in the PA production in seeds [12, 34, 35].
Given the widespread occurrence of LAR in various plant lineages and its absence in A. thaliana, understanding the evolutionary trajectory of LAR genes can provide valuable insights into their functional diversification and potential compensatory mechanisms in species lacking LAR. Here, we investigate the evolutionary dynamics of LAR through a systematic analysis of its presence/absence across land plant lineages. Our findings reveal a deep duplication event at the base of the evolutionary split between gymnosperms and angiosperms that has resulted in many plant species harboring two LAR copies, suggesting potential subfunctionalization or neofunctionalization within these paralogs. However, we also identified entire plant lineages without LAR. Most notably, LAR appears to be largely missing within the Brassicaceae family. Additionally, we explored the functional divergence of the two LAR lineages in plants with multiple copies, aiming to understand how gene duplication has influenced the evolution and PA biosynthesis.
Results
Phylogenetic analysis of leucoanthocyanidin reductase (LAR)
This study explored the phylogenetic relationships of LAR genes across a wide range of plant taxa. In total, 287 LAR, 268 DFR, and 112 ANR sequences were retained for alignment and phylogenetic analysis. Four primary clades were identified in the LAR portion of the tree, corresponding to the major plant lineages: gymnosperms, monocotyledons, magnoliids, and dicotyledons (Fig. 2). Within both the gymnosperm and dicotyledon clades, two distinct subclades were identified. These will be referred to here as LARI and LARII for those in the gymnosperm clade (shown in orange and green in Fig. 2, respectively) and LAR1 and LAR2 for those in the dicotyledon clade (shown in red and blue in Fig. 2, respectively). These distinctions are necessary to highlight clear phylogenetic divergence within each lineage and to avoid confusion in downstream analysis.
Interestingly, the presence of separate LAR gene copies from the same species in both subclades of gymnosperms (LARI and LARII), as well as in both subclades of dicotyledons (LAR1 and LAR2), implies that an ancient gene duplication event occurred prior to the diversification of each of these groups (Fig. 2). This deep duplication appears to have given rise to two major evolutionary lineages of LAR genes in both gymnosperms and dicots. Following this deep duplication, additional lineage-specific duplication events occurred at the family, genus, or species level independently within each clade. For example, in gymnosperms, species such as Thuja plicata,* Picea sitchensis*, and Cryptomeria japonica present multiple LAR copies distributed across the subclades LARI and LARII. In dicots, the LAR1 subclade represents a particularly large and diverse collection of gene copies. Many species exhibit multiple paralogs within this subclade, pointing to repeated gene duplication events within specific lineages. Families such as Theaceae,* Fabaceae*,* Rosaceae*, and Malvaceae are especially well represented in LAR1, often with two or more copies per species. The LAR2 subclade, while smaller, also includes multiple gene copies of species present in LAR1. Notably, species such as Bretschneidera sinensis (Akaniaceae) and Carica papaya (Caricaceae) both belonging to the Brassicales order, contain LAR gene copies only in LAR1, suggesting a potential absence of the LAR2 copy in these lineages. This pattern further supports the idea that, following the initial ancient duplication, LAR1 and LAR2 have undergone independent expansions in different lineages, leading to a potential clade-specific diversification of the LAR genes.
Fig. 2. Phylogenetic analysis of LAR, DFR, and ANR sequences in different plant species. Sequences were collected via KIPEs3 results, codon-based alignments were generated with MACSE [36], and the phylogeny was inferred with IQ-TREE2 [37, 38] based on maximum likelihood. Distinct clades of LARI (orange) and LARII (green) in the Gymnosperm group as well as LAR1 (red) and LAR2 (blue) in the Dicotyledon group are highlighted. The purple arrows indicate the sequences used for the synteny and gene expression analysis. The complete tree, including all non-collapsed clades, is available in Additional File 1C [39]
Leucoanthocyanidin reductase duplication event and functional divergence
The phylogenetic analysis (Fig. 2) revealed the presence of multiple LAR gene copies in several species within both gymnosperms and dicotyledons. This pattern points to an ancestral gene duplication event, followed by divergence within each lineage. Not only these gene copies are phylogenetically distinct, but also, they are not located near each other in the genome (See Additional file 1D [39]). This suggests that they did not arise from recent tandem duplication but rather from older duplication events followed by independent evolutionary paths. Over time, these copies have likely diverged in sequence and regulation, which may reflect differences in function or tissue-specific roles. To explore this potential functional divergence, gene expression patterns of the different LAR paralogs were examined across multiple tissues in representative species presented in the phylogenetic tree. Among gymnosperms, Taxus chinensis, Ginkgo biloba, Pinus pinaster,* Picea sitchensis*, and Thuja plicata were analyzed (Fig. 3 [39]), and among dicotyledons the analyzed species included: Vitis vinifera, Theobroma cacao, Moringa oleifera,* Gossypium hirsutum*,* Actinidia chinensis*,* Eucalyptus grandis*,* Populus trichocarpa*,* Carya illinoinensis*, and Morella rubra (Fig. 4; [39]). T. chinensis showed a significant difference in expression between TchLARI (KAH9310580.1) and TchLARII (KAH9295439.1). TchLARII showed consistently higher expression across all the sampled tissues (Fig. 3A). A similar pattern was observed in G. biloba, where GbiLARII (GBI00018207) had a significantly higher overall expression than GbiLARI (GBI00017466) with tissue-specific expression patterns showing stronger GbiLARII expression in seedling stem tissues (p-value = 0.0004) and root tissues (p-value = 0.002), and a stronger GbiLARI expression in stem tissues (p-value < 0.0001) (Fig. 3B). In P. pinaster, a significant overall expression difference was also detected between PpiLAR1 (PPI00016665) and PpiLARII (PPI00057446), with PpiLARII being more highly expressed in needles (p-value < 0.0001), shoots (p-value < 0.0001), and stem tissues (p-value < 0.0001) (Fig. 3C).
In dicots, clear expression differences between paralogs were also identified. In V. vinifera,* VviLAR2* (VIT_217s0000g04150.2) showed significantly higher expression than VviLAR1 (VIT_201s0011g02960.1) in leaf tissues (p-value < 0.0001) and berry tissues (p-value < 0.0001) (Fig. 3D). In T. cacao, the gene expression of TcaLAR1 (Thecc.02G361400.1) was dominant in cells, inflorescence, leaf, seedlings, young red leaves, and mix of leaf and flower tissues while TcaLAR2 (Thecc.03G028300.1) presented higher expression in pod (p-value < 0.0001) and pistil tissues (p-value < 0.0001) (Fig. 3E). Similarly, in M. oleifera, a significant overall expression difference was detected between MolLAR1 (g8395.t1) and MolLAR2 (g11921.t1), with MolLAR2 exhibiting higher expression in roots (p-value = 0.002), leaf (p-value < 0.0001), and stem tissues (p-value = 0.005) (Fig. 3F).
Fig. 3. Comparison of the expression of LARI (orange) and LARII (green) genes in the species Taxus chinensis (A), Ginkgo biloba (B) and Pinus pinaster (C), and LAR1 (red) and LAR2 (blue) in the species Vitis vinifera (D), Theobroma cacao (E), and Moringa oleifera (F) genes across different plant tissues. An asterisk above the violin plot indicates a statistically significant difference (p < 0.05) between LARs in that specific tissue, based on a linear mixed-effects model followed by estimated marginal means analysis. The complete results of the LMM and EMMs are provided online [39].
When comparing the 36 sequences of LARI and the 11 sequences of LARII in gymnosperms, both groups showed strong conservation at many positions, numbered according to the Theobroma cacao LAR1 (Thecc.02G361400.1) reference sequence (Fig. 4A [39]). Particularly interesting are positions with contrasting differences between LARI and LARII (Fig. 4A). In dicotyledonous species, 106 sequences of LAR1 and 43 sequences of LAR2 were analyzed, revealing comparable conservation with notable divergences at several key residues (Fig. 4A [39]). Conserved regions suggest key functional or structural roles. Amino acid substitutions at five specific positions in gymnosperms (18, 117, 126, 161, 167) and seven in dicots (15, 117, 157, 161, 167, 168, 255) clearly differentiate the gymnosperm LARs (LARI, LARII) from their dicot counterparts (LAR1, LAR2). At position 15, a consistent presence of alanine (A) was found in LARI, LARII, and LAR1, while a substitution to serine (S) was observed in LAR2. At position 18, tyrosine (Y) was exclusive to LARI, while phenylalanine (F) was dominant in LARII and dicots (Fig. 4B). At position 117, valine (V) was retained in both LARII and LAR1, while isoleucine (I) was found in LARI and LAR2. A unique alanine (A) in LARII was observed in position 126, substituting glycine (G) in LARI and dicots. The ICCN motif region centered on position 157 showed a particular divergence in dicots; while serine (S) is highly conserved in LAR1, alanine (A) is more conserved in LAR2. Similar results can be observed at position 167: serine (S) was conserved in LARI and LAR1, while threonine (T) appeared in LARII, and alanine (A) was found in LAR2. Given the proximity of this position to the active site and the polarity differences, this variation was interpreted as functionally relevant (Fig. 4C). An additional substitution at position 168 within a conserved motif replaces glutamate (E) in LARI, LARII, and LAR1 with aspartate (D) in LAR2, reducing the side chain length and potentially altering charge distribution or hydrogen bonding patterns. Finally, position 255, while this has not been reported as part of any LAR-specific motif, showed a predominant conservation of asparagine (N) in all groups except LAR2, which presented a substitution to methionine (M). This polar-to-nonpolar shift, occurring near the active site, was interpreted as likely contributing to a reduction in the catalytic efficiency of LAR2 as reported by the gene expression in Fig. 3E for T. cacao. The proximity of several variable positions to conserved motifs and the active site further supports their relevance in the functional diversification of LAR enzymes.
Fig. 4. Functional divergence in LAR proteins. A Comparative sequence logo showing conservation and variability across LARI and LARII sequences in gymnosperm species, and LAR1 and LAR2 sequences in dicotyledon species. The X-axis represents the position based on Theobroma cacao LAR1 (Thecc.02G361400.1) reference sequence, while the Y-axis indicates the frequency in percentage, reflecting sequence conservation at each position. Individual amino acids are color-coded based on their chemical properties: nonpolar/hydrophobic residues (black), polar amino acids with hydroxyl groups (green), polar amino acids with amide residues (purple), positively charged residues (blue), and negatively charged residues (red). The height of each letter within a column corresponds to its relative frequency at that position with taller letters indicating more conserved regions and shorter ones reflecting variability. B Structural model of LARI (orange) and LARII (green) of the gymnosperm species Taxus chinensis, highlighting five positions with amino acid substitution near the active site of the protein. C Structural model of LAR1 (red) and LAR2 (blue) of the dicotyledon species Theobroma cacao, highlighting seven positions with amino acid substitution near the active site of the protein
The combined analysis of gene expression patterns and polypeptide seq uence conservation indicates a clear functional differentiation between the LAR gene groups. LAR1 and LAR2 in dicots, as well as LARI and LARII in gymnosperms, exhibit distinct tissue-specific expression profiles. These expression patterns are supported at the molecular level by systematic amino acid substitutions revealed through the comparative analysis of LAR sequences. These substitutions differentiate dicots and gymnosperm LARs, as well as within each LAR lineage. Mutations such as Y18F in gymnosperms LARII, and S157A and E168D, both occurring in dicots LAR2, present particular interest due to occurring within conserved motifs.
Expression analysis among LAR1 and LAR2 genes
Given the distinct expression patterns and systematic amino acid differences previously identified between LAR1 and LAR2, a coexpression analysis was conducted to further investigate their potential functional divergence and relationships with other metabolic pathways. The results are presented in two tables, in which each LAR gene is depicted along with a list of coexpressed genes. For each gene, the Spearman correlation coefficient and gene expression value are provided (See [39]). In V. vinifera,* VviLAR1* (VIT_201s0011g02960.1) was notably coexpressed with the genes; VIT_206s0004g08150.1 (r = 0.710) encoding trans-cinnamate 4-monooxygenase, a key enzyme in the biosynthesis of flavonoids and lignin, and VIT_213s0067g02870.2 (r = 0.657) encoding the enzyme chalcone-flavone isomerase, involved in the flavonoid biosynthesis. In T. cacao the analysis identified strong correlations between TcaLAR1 (Thecc.02G361400.1) and a diverse set of high expressed protein-coding genes involved in stress responses such as proline-rich protein-1 (Thecc.04G253900.1, r = 0.666) and the bax inhibitor-1 (Thecc.04G271000.1, r = 0.683). Additionally, genes involved in signaling pathways like Calmodulin7 (Thecc.09G265600.2, r = 0.763) which plays an important role in seedling development, and vesicle trafficking like ADP-ribosylation factor (Thecc.01G320100.1, r = 0.708) and ESCRT-related protein CHMP1B (Thecc.06G106700.1, r = 0.695). MolLAR1 (g8395.t1) in M. oleifera, presented strong correlation with genes encoding metallothionein-like protein 1 (g23076.t1, r = 0.702), copper transport protein ATX1 (g22635.t1, r = 0.701), and Profilin2 (g19932.t1, r = 0.682).
Regarding the genes coexpressed with LAR2, in V. vinifera the gene expression of VviLAR2 (VIT_217s0000g04150.2) showed stronger correlation with an isoform of the same gene (VIT_217s0000g04150.7, r = 0.741), followed by genes encoding sigma intracellular receptor 2 (VIT_212s0059g02290.1, r = 0.664), sucrose transport protein (VIT_218s0076g00250.1, r = 0.658), and patatin-like protein 2 (VIT_218s0001g10830.1, r = 0.651). In T.cacao, TcaLAR2 (Thecc.03G028300.1) coexpressed strongly with the genes encoding the protein SPIRAL1 (Thecc.02G328900.1, r = 0.681), GTP-binding nuclear protein (Thecc.09G120500.1, r = 0.722), and 60 S ribosomal protein (Thecc.01G108600.1, r = 0.707). Lastly, MolLAR2 (g11921.t1) in M. oleifera showed coexpression with genes encoding proteins such as WALLS ARE THIN1 (g3827.t1, r = 0.684), Major latex protein-like 43 (g15381.t1, r = 0.672), and triose-phosphate/phosphate translocator (g12330.t1, r = 0.665).
Coexpression analyses further supported divergence among LAR paralogs, with TcaLAR1 and MolLAR1 showing strong positive correlations with genes involved in protein regulation and stress response. Likewise, the coexpression profile of TcaLAR2 was strongly related to tissue-specific expression, specifically to reproductive development, consistent with its higher expression in pods and flower tissues.
Regulatory divergence between LAR1 and LAR2 gene promoters and identification of transposable elements
To explore the regulatory divergence between the two LAR gene copies, promoter sequences from V. vinifera,* T. cacao*, and M. oleifera were extracted and analyzed for transcription factor binding motifs and transposable elements (TE) insertions. This species-specific comparison provides additional insights into the functional differentiation of these LAR paralogs and highlights the potential influence of TEs on LAR gene regulation, particularly in the context of proanthocyanidin biosynthesis.
In V. vinifera, both promoters pVviLAR1 and pVviLAR2 included motifs associated with flavonoid biosynthesis gene regulation, notably those bound by MYB transcription factors. pVviLAR1 exclusively contained ABRELATERD1 motifs and shared with pVviLAR2 the presence of MYCCONSENSUSAT, MYBCORE, MYB1AT, MYBGAHV, and MYB2CONSENSUSAT. In contrast, pVviLAR2 uniquely harbored a MYBPZM motif. While both genes showed considerable overlap in motif composition, pVviLAR2 presented elements in closer proximity to the transcription start site (TSS) and a more varied motif composition, which may reflect a broader regulatory role (Fig. 5A). Additionally, TEs annotated with EDTA revealed a Helitron TE insertion in pVviLAR1 and a Mutator-like TIR transposon in pVviLAR2.
In T. cacao, both promoter regions displayed a diverse motif landscape sharing elements such as MYB1AT, MYBCORE, MYBPZM, and ABRELATERD1. Notably, pTcaLAR1 presented a higher number of motifs overall and uniquely included multiple MYCCONSENSUSAT elements, while pTcaLAR2 was characterized by the presence of MYBGAH (Fig. 5B). In M. oleifera, the promoter region pMolLAR1 presented a richer and more dense motif profile compared to pMolLAR2. Shared motifs included MYB1AT, MYBCORE, MYCCONSENSUSAT, MYBPZM, and MYBPLANT. In addition, ABRELATERD1 and MYB2CONSENSUSAT were exclusively found in pMolLAR1. (Fig. 5C). No TE insertions were detected in the promoter regions of either LAR gene in T. cacao and M. oleifera. Collectively, these findings reveal both conservation and divergence in the regulatory landscapes of LAR1 and LAR2 across species. MYB and MYC motifs were consistently found and are likely essential for proanthocyanidin biosynthesis regulation. However, the presence of TEs in the LAR promoters of V. vinifera, in contrast with their absence in T. cacao and M. oleifera, point toward species-specific regulatory adaptations, supporting the hypothesis that promoter evolution via motif variation and TE insertions may have contributed to species-specific expression patterns of LAR paralogs.
Fig. 5. Promoter region constructs with putative cis-acting regulatory elements (CREs) predicted by the PLACE database [40]. A pVviLAR1 and *pVviLAR2 in Vitis vinifera. *B pTcaLAR1 and pTcaLAR2 in Theobroma cacao. C pMolLAR1 and pMolLAR2 in Moringa oleifera. CREs are displayed above each promoter schematic, and transcription start sites (TSS) are indicated in red. The 5’ untranslated region (5’ UTR), shown in gray, is a schematic representation and not proportional to bp length. A comprehensive list of CREs for each promoter is available online [39].
Loss of LAR in families Cleomaceae and Brassicaceae
To confirm the absence of LAR in the Cleomaceae and Brassicaceae families we performed a synteny analysis that first compared the genome sequences of three species belonging to different Brassicales families: Moringa oleifera (Moringaceae), Carica papaya (Caricaceae), and Bretschneidera sinensis (Akaniaceae) in context with the outgroup species Theobroma cacao (Malvaceae) and Vitis vinifera (Vitaceae) (Fig. 6A). Microsynteny confirmed the presence of LAR1 among above-mentioned species of the Brassicales order and the outgroups. However, when repeating the analysis only among members of the Brassicales order, this time including Cleome gynandra (family Cleomaceae), a close relative of the Brassicaceae, as well as the Brassicaceae species: Aethionema arabicum,* Capsella rubella*,* Boechera stricta*,* Arabidopsis thaliana*,* Arabidopsis halleri*,* Eutrema salsugineum*, and Brassica rapa, no LAR1 gene was detected among the Cleomaceae and Brassicaceae species (Fig. 6B).
After discovering two independent LAR clades through the phylogenetic analysis, it was necessary to determine whether LAR2 is present among the Brassicales families. This synteny analysis included as reference the genome sequence of V. vinifera with the region where LAR2 is localized. LAR2 was only observed in the syntenic position between V. vinifera, T. cacao, and M. oleifera, corresponding to the LAR2 presence in these species as revealed by the phylogenetic analysis (Fig. 6C). Like LAR1, LAR2 was also not detected in multiple Brassicales families in this synteny analysis (Fig. 6D).
Fig. 6. Synteny gene identification of species in the Brassicales order. A Synteny around LAR1 (indicated in pink) among the genome sequences of V. vinifera,* T. cacao*,* M. oleifera*,* C. papaya*, and B. sinensis. B Synteny around LAR1 among the genome sequences of M. oleifera,* C. papaya*,* B. sinensis*,* C. gynandra*,* A. arabicum*,* C. rubella*,* B. stricta*,* A. thaliana*,* A. halleri*,* E. salsugineum*, and B. rapa. C Synteny around LAR2 (indicated in light blue) among the genome sequences of V. vinifera,* T. cacao*,* M. oleifera*,* C. papaya*, and B. sinensis. D Synteny around LAR2 among the genome sequences of M. oleifera,* C. papaya*,* B. sinensis*,* C. gynandra*,* A. arabicum*,* C. rubella*,* B. stricta*,* A. thaliana*,* A. halleri*,* E. salsugineum*, and B. rapa. A complete list of syntenic gene positions and coordinates used for this analysis is provided in [39]
Discussion
The flavonoid biosynthesis pathway is generally believed to be well conserved across different plant taxa [18, 41]. The evolution of flavonoids has been extensively studied, including the distinct evolutionary mechanisms that led to some of the individual branch products of the pathway [42–44]. Evolution of this biosynthetic pathway and novel traits are often achieved by changes in transcription factors [45–47]. Such findings have provided significant insights into the genetic and enzymatic variations that contribute to the diversity of flavonoid compounds among plant species [48–50]. Research has revealed that gene duplications and subsequent functional divergences are key processes in the evolution of flavonoid biosynthesis [45, 51]. These genetic variations result in a wide array of flavonoid structures, many of which have well-reported biological functions, such as antioxidant activity [52], UV protection [53], and attraction of pollinators and seed dispersers [54]. Additionally, studies have highlighted the role of regulatory genes in controlling the expression of genes encoding enzymes involved in flavonoid production [3, 47, 55]. Our findings on LAR highlight the mechanism by which gene duplication, divergence, and loss of the LAR gene have driven functional diversification in the context of proanthocyanidin biosynthesis across plant species.
Evolutionary Dynamics of LAR Gene Duplications
The phylogenetic analysis revealed that LAR has undergone complex evolutionary trajectories in both gymnosperms and dicotyledons. Within the dicotyledon group, LAR genes are segregated into two major clades, referred to as LAR1 and LAR2, which likely originated from an ancient duplication event in the common ancestor of the eudicotyledons. This bifurcation of LAR genes into distinct evolutionary paths denotes selective pressure favoring the maintenance of both paralogs, possibly due to their acquisition of distinct or complementary functions.
Evidence of this phylogeny pattern in LAR has been previously reported by Wang et al. [27], who reported that plant LARs present three distinctive groups, including monocotyledons, gymnosperms, and dicotyledons, with the latter further splitting into two clades. The authors also noted the presence of sequence-level divergence due to a DNA transversion and transition in a codon of the LAR-specific motif ICCN [27]. This finding was corroborated by our results, which also contain a serine residue in LAR1 substituted by alanine in LAR2. However, we also observed a similar duplication pattern in gymnosperms, with gene copies clustering into two distinct clades, LARI and LARII. Similar to dicots, these clades do not correspond to recent tandem duplication, as the copies are found in separate genomic regions, suggesting an ancient duplication event followed by long-term divergence. Notably, LARII in gymnosperms shows strong amino acid sequence similarity to the LAR paralogs in dicots, supporting the hypothesis that the duplication originated in a common ancestor of seed plants [56]. Under this scenario, one of the ancestral copies was potentially retained and further duplicated within dicots, giving rise to LAR1 and LAR2, while no such duplication has been observed in monocots. This evolutionary model implies that the LAR gene family underwent differential retention and lineage-specific diversification across major plant groups. Additionally, LARII in gymnosperms tend to show higher expression across several tissues compared to LARI, inferring the possibility that regulatory divergence may also have contributed to their functional differentiation and evolutionary paths.
Functional divergence between LAR1 and LAR2
The functional differentiation of LAR1 and LAR2 is supported not only by their phylogenetic divergence but also by their distinct expression profiles. Gene expression analyses across several dicot species revealed consistent differences between the two copies. For instance, TcaLAR1 is predominantly expressed in vegetative tissues such as leaves and seedlings, highlighting its role in primary metabolism and general stress responses. This is further supported by coexpression of TcaLAR1 with stress-related genes, including proline-rich protein-1, bax inhibitor-1 and Calmodulin7, which are known to be involved in abiotic stress tolerance [57–59]. In contrast, TcaLAR2 presented higher expression in reproductive tissues, such as pods and pistils, indicating a more specialized function in these organs. TcaLAR2 was found to be coexpressed with genes involved in structural and regulatory processes, including the protein SPIRAL1, known for its expression in tissues undergoing rapid cell expansion [60], these findings support its potential role in reproductive tissue development. This pattern of tissue-specific functional differentiation points to a potential adaptive significance of maintaining multiple LAR copies within a given species. This divergence is also evident in gymnosperms. For example, in T. chinensis and G. biloba, LARII consistently showed higher expression across several tissues compared to LARI. In P. pinaster, LARII was strongly expressed in needles and shoots, reinforcing the notion that regulatory divergence is an independent outcome following LAR duplication in gymnosperms and dicots. Variation in expression patterns among species suggests that these LAR duplicates may have undergone lineage-specific adaptations shaped by ecological or developmental constraints. These findings align with previous studies that have demonstrated how gene duplication events can give rise to paralogs through subfunctionalization or neofunctionalization allowing species to optimize their resources and improve their environmental adaptation [61, 62].
Sequence comparisons among LAR paralogs across gymnosperms and dicots further revealed both conserved patterns and lineage-specific divergences, particularly at functionally relevant motifs. Among gymnosperm copies, LARII exhibited sequence similarity to dicot LARs (LAR1 and LAR2), especially at conserved residues such as positions 157 and 168. This pattern, along with the consistently higher expression levels of LARII compared to LARI, supports the hypothesis that LARII may have retained more ancestral structural features, making it a more functionally conserved paralog than LARI. Additionally, the sequence variations identified between LAR1 and LAR2 reinforce the evidence for their functional divergence. One of the most prominent mutations involves S157A within the conserved ICCN motif, a region implicated in substrate binding and catalytic specificity of LAR enzymes [27, 63]. Polar amino acid residues like serine are known to facilitate hydrogen bonding with substrates or cofactors, supporting catalytic activity and proper substrate binding [64, 65]. In contrast, nonpolar residues such as alanine contribute primarily to structural stability and maintaining the integrity of the protein through hydrophobic interactions. The substitutions S157A and S167A in LAR2 may therefore alter the enzyme’s interaction dynamics or catalytic behavior. Although changes in protein structure do not directly control gene expression, structural divergence can result in functional specialization, which may in turn be accompanied by differential expression in different tissues. In this context, the specialized expression of TcaLAR2 in reproductive tissues could reflect a shift in its biochemical role, favored by changes in substrate preferences, activity, or protein-protein interactions. Supporting this, Liu et al. [23] reported a similar expression profile of a T. cacao LAR gene located on chromosome 3, which matches the position of TcaLAR2 in this study. Liu et al. reported a higher expression in pods and seeds, supporting the idea that TcaLAR2 may be functionally specialized for reproductive tissue processes such as proanthocyanidin biosynthesis during pod development [23]. Another substitution potentially linked to tissue-specific expression is E168D in LAR2, a variation that also lies within a conserved motif. Although both glutamate and aspartate are negatively charged, the shorter side chain of aspartate can reduce enzyme efficacy and alter substrate specificity. While these effects have not been specifically demonstrated in LAR proteins, similar glutamate-aspartate substitutions in other enzymes have been shown to impair catalytic activity and binding properties [66–68]. In addition, the substitution N255M, which lies adjacent to the active site, was also identified. Asparagine (N), conserved in LARI, LARII, and LAR1, is replaced by methionine (M) in LAR2. As a polar residue, N255 commonly participates in hydrogen bonding, these bonds help stabilize the configuration of the active site [69]. Additionally, the presence of a polar sidechain can contribute to the potential electrostatic interactions, enhancing substrate binding [70]. Contrarily, M255 in LAR2 is larger and has a nonpolar sidechain, which may disrupt these interactions with a possible decrease in catalytic efficiency through altered substrate positioning or transition state stabilization [71, 72]. The proximity of these changes to conserved motifs and active sites suggests that they may impact enzyme stability as well. These findings align with a previous report [27], which highlighted the importance of specific amino acid changes in the evolution of LAR in C. sinensis.
Together, these amino acid substitutions in LAR sequences point to biochemical divergence that may contribute to tissue-specific functions. Their location within or in proximity to conserved motifs can be interpreted as indicative of evolutionary shifts likely affecting substrate affinity or motif function. While our findings provide evidence for structural differences, they also propose new directions for understanding how such molecular changes might relate to the distinct functional roles observed in LAR across species.
Role of CREs and TEs in divergence of LAR1 and LAR2 regulation
The diversification of the flavonoid biosynthesis pathway in angiosperms is closely tied to the evolutionary plasticity of its regulatory hotspots [48, 73], including those involved in the reduction of leucoanthocyanidins to catechins via LAR [74, 75]. In this study, the promoter regions of LAR1 and LAR2 from V. vinifera,* T. cacao*, and M. oleifera were analyzed to assess regulatory divergence through the identification of cis-regulatory motifs (CREs) and transposable elements (TEs). Our findings suggest that promoter divergence, modulated by cis-element variation and TE insertions, may explain functional specialization of the two LAR paralogs in a lineage-specific manner. Promoter region analysis revealed a common pattern involving MYB, and bHLH binding elements across all species, consistent with the well-documented role of members of these TFs families in regulating flavonoid biosynthesis [76]. Several MYB TFs are established regulators of the phenylpropanoid and flavonoid pathways [77, 78], specific members such as MYBPA1 and MYBPA2 in V. vinifera, and MYB5 and MYB14 in M. truncatula have been shown to activate the expression of LAR and ANR genes, thereby controlling proanthocyanidin biosynthesis [79–81]. Similarly, bHLH transcription factors, which function as part of the MBW complex, not only assist MYBs in regulating anthocyanin and proanthocyanidin production but also modulate gene expression dynamically in response to developmental stages and environmental stimuli [82]. In this study, the VviLAR2 promoter presented a higher abundance of MYC-related cis-elements (E-boxes), which are canonical binding sites for bHLH transcription factors such as MYC1, MYC2, and MYCA1 [83–85]. Notably, MYC1 has been shown to interact with R2R3-MYB proteins to coregulate anthocyanins and PA biosynthesis during berry development [84]. The enrichment of bHLH-binding motifs in the VviLAR2 promoter may therefore contribute to tissue-specific expression of VviLAR2, with particularly high levels in grape berries. Additionally, previous studies of the promoter regions of both LAR genes in V. vinifera have shown that VviLAR1 expression is sensitive to light conditions, with higher expression under light and repression in the dark, correlating with the activity of bHLH transcription factors such as MYC2 and MYCA1, the latter being upregulated in callus tissues under dark conditions [86]. Our promoter analysis supports these regulatory patterns; pVviLAR1 presents multiple light-responsive elements (LREs), such as GT1-related motifs [39]. While the presence of MYB and bHLH binding sites in LAR promoters does not provide functional evidence, it suggests the potential regulation by these TF families. This supports the hypothesis that LAR expression, and by extension, PA biosynthesis may be modulated in response to developmental and environmental cues, consistent with the dual roles of these specialized metabolites in pigmentation and protection [87]. Overall, these findings suggest that VviLAR1 is predominantly regulated by light-responsive pathways, whereas the enrichment of MYC-binding motifs in pVviLAR2 implies that this paralog may be more responsive to bHLH-mediated regulation.
In T. cacao,* pTcaLAR1* exhibited a higher overall abundance of MYB motifs. Among these elements is MYBCORE (CNGTTR), a canonical target for R2R3-MYB proteins. This motif is recognized by MYB.Ph3 in Petunia hybrida, known to be involved in regulating anthocyanins and PA biosynthetic genes in petal epidermal cells [88]. Moreover, pTcaLAR2 contains the MYBGAHV motif (TAACAAA) which is not found in pTcaLAR1. This element is the specific binding site for GAMYB, a gibberellin (GA) inducible MYB transcription factor, shown to activate GA-responsive genes such as high-pl α-amylase in barley aleurone cells [89]. The presence of this cis-element in pTcaLAR2 suggests that this paralog may be selectively responsive to GA signaling, potentially linking its regulation to flower and seed development. GA is known to play critical roles in the transition from vegetative to reproductive phase including the formation of floral organs such as seed and pollen tube growth [90, 91]. This aligns with the enrichment of POLLEN1LELAT52 motif in pTcaLAR2, a cis-element known for pollen-specific expression [92]. These findings further support the hypothesis that TcaLAR2 may be subject to a more restricted regulatory profile, with activity concentrated mostly around reproductive organs. In M. oleifera, pMolLAR2 is characterized by a higher number of flavonoid-related motifs, including MYBPZM and MYBPLANT with the latter being reported as binding site for transcription factors such as AmMYB305 and AmMYB340 in Antirrhinum majus, which regulate phenylpropanoid and lignin biosynthesis [93].
The differential presence of TE insertions within the LAR promoter regions may also play a significant role in the regulatory divergence between LAR1 and LAR2. In V. vinifera, both promoters contain TE insertions, but differ in their composition and position. These elements are inserted in different regions within the promoters and genes, likely influencing their expression. TE insertion can influence gene regulation by introducing or removing cis-regulatory elements, modulating chromatin accessibility, or even acting as alternative enhancers or repressors [94]. For instance, LTR retrotransposons have been associated with pigmentation loss in crops such as grape, blood orange, and apple by modulating the expression of anthocyanin biosynthesis genes [95–97]. However, the extent to which these regulatory disruptions in the anthocyanin pathway influence proanthocyanidin production remains unclear. By contrast, no transposable elements were detected in the LAR promoters of T. cacao and M. oleifera, indicating a more conserved promoter architecture.
Across all three species, promoter region analyses revealed the presence of MYB and bHLH binding motifs associated with LAR regulation, suggesting that particular members of these TF families contribute to the conserved control of PA biosynthesis. However, the distribution and abundance of these conserved motifs, along with their proximity to the TSS and presence of species-specific or paralog-specific elements, supports a model in which **LAR **gene duplicates have acquired distinct regulatory profiles.
Insights into the loss of LAR in the families Cleomaceae and Brassicaceae
The absence of LAR genes in the Cleomaceae and Brassicaceae families, as revealed by the synteny analysis, presents an interesting case of metabolic adaptation. While previous studies have not reported the presence of LAR homologs in A. thaliana [20] or in species of the Brassica genus [98], our results extend this observation to a broader sampling within the family and beyond. Given that A. thaliana predominantly accumulates PAs composed of (-)-epicatechin, the presence of (+)-catechins, and consequently a functional LAR gene, has not yet been reported in this species [20]. The retention of other proanthocyanidin biosynthesis genes in Brassicaceae [98] points to a functional compensation mechanism or a shift in metabolic compounds that led to LAR not being necessary in the production of proanthocyanidins. It is possible that the pathway in Brassicaceae species has evolved to rely solely on ANR activity for the synthesis of PAs, unlike M. truncatula, which dual activity of LAR has been reported in the production of catechins and its involvement with ANR in the synthesis of epicatechins [21]. This selective loss within the common ancestor of the Cleomaceae and Brassicaceae families hints at the likelihood that alternative enzymatic pathways or redundant mechanisms might compensate for the absence of LAR. Similar patterns of LAR loss have been observed in other lineages. For example, both Zea mays and Sorghum bicolor lack apparent LAR homologs [99]. While maize shows low PA levels and relies solely on ANR to produce epicatechin-based PAs, sorghum is a PA-rich species with a more complete set of regulatory transcription factors, including TT2-type MYBs, synthesizing catechin and procyanidin B3 dimers as major PA components [99]. This divergence illustrates that although both species share the absence of LAR, their metabolic compensation strategies and PA profiles are lineage-specific. Such metabolic shifts are not uncommon in plant specialized metabolism. Similar cases include the mutual exclusivity of anthocyanin and betalain pathways in the Caryophyllales order [100, 101], or the presence of glucosinolates that are largely confined to Brassicales [102]. These examples highlight how evolutionary pressures shape biosynthetic pathways differently across taxa.
Although A. thaliana has been instrumental in elucidating the core steps of the flavonoid biosynthesis, its limitations, such as the lack of LAR, highlight the importance of exploring additional model species. Recent reviews have proposed new model systems better suited for studying the full diversity of PA biosynthetic pathway, particularly in species where both LAR and ANR are present [31, 103]. In this context, crops such as V. vinifera and T. cacao, where the LAR function is well-documented and linked to seed development and PA accumulation [22, 23], could provide opportunities to better understand species-specific enzymatic adaptations.
This study confirms and adds to previous findings by providing additional evidence that LAR is likely absent from a broader range of families within the Brassicales order. The release of new genome sequences from additional Brassicales species in the next years will allow to narrow down these loss events, which might affect more families beyond Cleomaceae and Brassicaceae. Together, these findings highlight the dynamic nature of plant specialized metabolism emphasizing the value of integrating model and non-model systems to fully grasp the evolution and diversification of biosynthetic pathways.
Conclusion
We conclude that the four distinct LAR clades have functional differences i.e., with respect to gene expression and enzymatic properties. However, a universal pattern of functional difference was not discovered and might not exist due to secondary evolutionary events. Interestingly, LAR1 was represented by a higher number of sequences compared to LAR2. While this may partly reflect differences in species-richness, genome sequence availability, and annotation quality, it could also indicate that LAR1 is more broadly retained and functionally important across dicots. This overrepresentation of LAR1 may reflect a central role in proanthocyanidin biosynthesis and stress responses as indicated by coexpression results. Future research could focus on exploring the biochemical properties of LAR1 and LAR2 enzymes in greater detail to better understand whether these differences result from subfunctionalization or neofunctionalization. Additionally, investigating the interaction dynamics influencing substrate specificity could provide valuable insights into the evolutionary and ecological significance of proanthocyanidin metabolism in plants. Most importantly, future studies are encouraged to differentiate between members of the LAR1 and LAR2 clade in dicots or LARI and LARII clade in gymnosperms, respectively, when reporting about LAR.
Methods
Data collection
Two primary types of sequence datasets were collected for this study: genome assemblies and corresponding annotations for synteny analysis, and LAR, ANR, and DFR coding and peptide sequences for phylogenetic construction.
Genomic data for synteny analysis
The genome assemblies and corresponding gene annotation of 11 species were retrieved from Phytozome and NCBI genome database (Table 1). These data were used for synteny analysis to evaluate genomic neighborhood conservation of the LAR gene and support putative loss events (See methods Sect. Synteny analysis).
Candidate sequences for phylogenetic analysis
To reconstruct the evolutionary history of LAR, DFR, and ANR genes, candidate orthologs were identified across a broad range of plant species using KIPEs v3 with default parameters [104]. Protein databases were screened using the bait and reference data FlavonoidBioSynBaits_v3.3 provided by KIPEs3, and only hits showing a 100% match in the conserved amino acid residues were retained to infer presence in a plant lineage. Partial candidates were inspected when inferring absence of LAR. The resulting sequences were then cleaned and mapped to their corresponding coding sequences (CDS) using a custom pipeline (see methods Sect. Alignment and phylogenetic tree construction). The species tree used to guide synteny comparisons and phylogenetic interpretation was obtained from [105].
Gene annotation
Since no annotation of the coding sequences was publicly available for Moringa oleifera, BRAKER v3.0.8 [106] with default settings was applied to generate a structural annotation of protein-encoding genes. To support the annotation process with hints, several RNA-seq data sets were retrieved from the Sequence Read Archive [107, 108] and aligned to the genome sequence with HISAT v2.2.1 [109] using default parameters. Samtools v1.20 [110] was used to sort the resulting BAM file which was then provided to BRAKER3 [106].
Alignment and phylogenetic tree construction
A collection of LAR, DFR, and ANR sequences were identified using KIPEs v3 [111]. Sequences were only considered if a strict match of 100% was found in the set of conserved amino acid residues defined in the KIPEs3 bait file, which ensures a high-confidence candidate identification. These sequences were selected for further analysis. To improve tree readability, sequences labeled as “putative”, “predicted”, or repetitive sequences were removed using a custom python script (clean_fasta.py), which is publicly available on GitHub: https://github.com/mariamarinr/LAR_Evolution/. This filtering step was designed to preserve taxonomic representation, retaining at least one high-confidence LAR sequence per major plant lineage. After the sequence cleaning, the corresponding coding sequence (CDS) was retrieved for each polypeptide sequence to enable codon-aware alignments. This was done using a custom pipeline that cross-referenced gene identifiers from the amino acid FASTA headers with available genome sequence annotations and CDS files (extract_kipes_cds.py) on GitHub: https://github.com/mariamarinr/LAR_Evolution/. Only sequences with matching identifiers to their corresponding polypeptide sequences were retained. In cases where formatting differences in the headers occurred between CDS and polypeptide sequence files, identifiers were standardized to ensure a perfect match. This ensured consistency between protein-level and nucleotide-level data for downstream phylogenetic analysis (see [39] for a complete list). Codon-aware multiple sequence alignments were constructed using MACSE v2.07 [36, 112] with default parameters. An initial phylogenetic tree was constructed with FastTree v2.1.10 [113] using the WAG substitution model and without bootstrap support for rapid visualization. To confirm the phylogenetic structure and arranged relationships, a more robust analysis was performed using IQ-TREE v2.0.7 [37, 38], with ultrafast bootstrap and 1000 replicates. The best-fit substitution model was identified using IQ-TREE’s ModelFinder function and selected according to the Bayesian Information Criterion (BIC). The chosen model was GTR + F + R10, which appropriately accounts for base frequency heterogeneity and rate variation across the different positions in the alignment. Tree visualization and annotation were conducted using the Interactive Tree Of Life (iTOL) v.7 [114].
Gene expression analysis
Paired-end RNA-seq datasets were retrieved from the Sequence Read Archive using fastq-dump v2.8.1 [107]. Gene expression quantification, including raw counts and Transcripts Per Million (TPMs), were obtained using kallisto v0.44 [115] with default parameters. Customs R scripts (prep_expression_matrix.R and LMM_expression_analysis.R) available through GitHub (https://github.com/mariamarinr/LAR_Evolution/) were developed to prepare the expression data matrix and generate violin plots using the ggplot2 package v3.5.1 [116] illustrating the variation in gene expression (TPMs) across multiple samples [39]. These plots were specifically used to visualize the expression levels of LAR across specific plant tissues in different species, aiming to identify patterns in gene expression and to gain insights into potential functions. To assess the statistical significance of the difference in gene expression levels between the two LAR copies, a linear mixed-effects model (LMM) was employed. The model was formulated as:
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$$TPM\_log\sim genes^\ast tissue+(1\vert sample)$$\end{document}where TPM_log represents the log-transformed transcript abundance; genes and tissue are fixed effects; and sample is included as a random effect to account for variability across biological replicates and sample-specific factors (e.g., physiological condition, developmental stage, or sequencing platform). This structure ensures more accurate estimates of fixed effects and reduces the risk of false positives. The model was fitted using restricted maximum likelihood (REML), and statistical significance for the fixed effects was assessed using t-tests with Satterthwaite’s method, as implemented in the lmerTest package v3.1–3 [117]. To further explore differences in gene expression across plant tissues and provide a more detailed evaluation of tissue-specific patterns for each LAR copy, a post-hoc pairwise comparison was conducted. This analysis was performed using the emmeans function v1.10.1 [118], which computed estimated marginal means (EMMs) [119] and pairwise contrasts for the interaction between genes and tissue. Specifically, pairwise comparisons were made for the genes within each level of the tissue factor, using t-tests to assess the significance of differences between the two LAR copies across the various tissues. To account for multiple comparisons, p-values were adjusted using the Bonferroni method. All statistical analyses were performed using R v4.3.3 [120].
Functionalization analysis
In order to explore the functional divergence of the LAR gene copies in the same species, a screen for evidence of subfunctionalization and neofunctionalization was conducted. This involved comparing the sequence profiles of the two LAR copies to identify potential nonsynonymous substitutions that would lead to novel functions that had evolved since the duplication event. Specifically, we utilized sequence alignments using MAFFT v7.490 [121] and visualized in JalView v.2.11.4.1 [122] and ChimeraX v.1.8 [123] to detect differences in the amino acid positions within the LAR proteins, which could indicate changes in functional domains or active sites. Additionally, the alignments were subjected to an analysis with DivAAs (https://github.com/k-georgi/DivAAs) to identify specific amino acid positions that show systematic differences between the two groups of LAR sequences.
Coexpression analysis
Coexpression analysis was performed to identify genes that present similarity in expression with the LAR genes of interest. A list with the LAR genes and a count table with normalized TPM values were used for this analysis. Pairwise Spearman correlation coefficients for gene expression values across all samples were calculated for each LAR gene in the list. The coexpressed genes were assessed based on their Spearman correlation values, adjusted p-values, and total expression level. Genes with low expression levels were excluded, and only gene pairs with a correlation coefficient > 0.65 and an adjusted p-value < 0.05 were considered significant. This coexpression analysis was implemented in a Python script - coexp3.py [51]. The functional annotation of the genes belonging to V. vinifera (Vvinifera_457_v2.1), T. cacao (Tcacao_523_v2.1), and M. oleifera (GCA_021397835.1) were derived from M. truncatula and A. thaliana using the Python script - construct_anno.py [51] available at: https://github.com/bpucker/ApiaceaeFNS1.
Identification of cis-regulatory elements (CREs) in LAR promoters
To investigate the differences in the regulatory mechanisms of LAR1 and LAR2 gene copies, the promoter region of each gene was extracted and analyzed for cis-regulatory elements. Promoter and transposable element (TEs) analyses were performed for V. vinifera, T. cacao, and M. oleifera, which were selected to represent distinct eudicot lineages (Vitales,* Malvales*,* Brassicales*, respectively) and to ensure the use of high-quality genome assemblies with reliable gene annotations. For V. vinifera, the LAR1 and LAR2 promoters previously reported by [86] were used, available in GenBank under accession numbers MT586116 and MT586117, respectively. For T. cacao and M. oleifera, paired-end RNA-seq reads were retrieved from the Sequence Read Archive (SRA) using fasterq-dump v.3.0.3 [124]. The complete list of SRR accessions is provided in [39]. To ensure accuracy in the identification of the transcription start sites (TSS), an alignment of these reads to their respective genome assemblies was performed using HISAT v2.2.1 [109], a splice-aware aligner optimized for short RNA-seq reads. This mapping workflow was automated using a custom Bash script (hisat2_mapping.sh) available at the GitHub repository: https://github.com/mariamarinr/LAR_Evolution/. This pipeline also handled the conversion, merging, sorting, and indexing of alignment files with Samtools v1.15 [110] for each species. The resulting sorted BAM files (available at [ 39 ]) were visualized using the Integrative Genomics Viewer (IGV) [125] to identify the regions with transcriptional activity upstream of LAR encoding sequences. Based on RNA-seq coverage patterns, the transcription start site (TSS) was inferred for each LAR gene copy, and promoter regions were defined as approximately 1500 bp upstream of the TSS. These promoter sequences were extracted from the genome assemblies using BEDtools v2.30 [126] (See additional file 4 [ 39 ]) and analyzed for potential cis-regulatory elements using the PLACE database [40] at https://www.dna.affrc.go.jp/PLACE/.
Identification of transposable elements (TEs)
To evaluate whether transposable elements (TEs) may contribute to the regulation of LAR gene expression, genome-wide annotation of TEs was performed for the species: V. vinifera, T. cacao, and M. oleifera. TE annotation was conducted for each genome sequence using the Extensive de-novo TE Annotator (EDTA) v.2.2.2 [127]. EDTA was run in sensitive mode with both annotation and evaluation options enabled to ensure a complete detection of both abundant and low-copy TEs. To identify overlaps between TEs and LAR loci, the intersect function of BEDTools v2.30 [126] was employed. This approach was intended to reveal TE insertions located within or adjacent to LAR genes or their promoter regions across the three species. Each overlapping TE was classified by type (e.g., LTR retrotransposons, TIR DNA transposons), and family, allowing further exploration of whether particular TE lineages were commonly linked with LAR regulatory regions. The complete list of overlapping TE insertions and their annotations is provided in [39].
Synteny analysis
Synteny analysis using JCVI/MCscan [128] was performed to visualize conserved genomic regions across the genome sequences of multiple species (Table 1). Flanking regions surrounding the identified LAR loci, typically including 10 to 15 protein-coding genes upstream and downstream, were manually selected based on their physical proximity to LAR and the preservation of local gene order, serving as reference points. Gene connections between species were manually validated and revised by comparing the predicted syntenic blocks with the phylogenetic relationships reported by [105], ensuring consistency between gene conservation and species divergence. To identify homologous genes or syntenic regions that might be missing in current annotations, BLAST v.2.13.0 [129] databases were constructed for multiple species using the ‘makeblastdb’ command, specifying nucleotide sequence types. Query sequences were then searched against these databases using ‘tblastn’.
Table 1. List of species used for synteny analysisSpeciesFamilyOrderData set IDDatabaseReference Vitis vinifera
Vitaceae
Vitales Vvinifera_457_v2.1Phytozome [130] Theobroma cacao
Malvaceae
Malvales Tcacao_523_v2.1Phytozome [131] Bretschneidera sinensis
Akaniaceae
Brassicales GCA_018105755.1NCBI [132] Moringa oleifera
Moringaceae
Brassicales GCA_021397835.1NCBI [133] Carica papaya
Caricaceae
Brassicales Cpapaya_113_ASGPBv0.4Phytozome [134] Eutrema salsugineum
Brassicaceae
Brassicales Esalsugineum_173_v1.0Phytozome [135] Capsella rubella
Brassicaceae
Brassicales Crubella_474_v1.1Phytozome [136] Boechera stricta
Brassicaceae
Brassicales Bstricta_278_ v1.2Phytozome [137] Arabidopsis halleri
Brassicaceae
Brassicales Ahalleri_765_v2.1.0Phytozome [138] Arabidopsis thaliana
Brassicaceae
Brassicales Athaliana_447_Araport11Phytozome [139, 140] Brassica rapa
Brassicaceae
Brassicales GCA_003434825.1NCBI [141] Cleome gynandra
Cleomaceae
Brassicales GCA_036759665.1NCBI [142] Aethionema arabicum
Brassicaceae
Brassicales Ae.arabicum_v3.1- [143]
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ullah C, Unsicker SB, Reichelt M, Gershenzon J, Hammerbacher A. Accumulation of Catechin and proanthocyanidins in black Poplar stems after infection by plectosphaerella populi: hormonal Regulation, biosynthesis and antifungal activity. Front Plant Sci. 2019;10. 10.3389/fpls.2019.01441.10.3389/fpls.2019.01441 PMC 687335231803202 · doi ↗ · pubmed ↗
- 2Yang Y, Yao G, Yue W, Zhang S, Wu J. Transcriptome profiling reveals differential gene expression in Proanthocyanidin biosynthesis associated with red/green skin color mutant of Pear (Pyrus communis L). Front Plant Sci. 2015;6. 10.3389/fpls.2015.00795.10.3389/fpls.2015.00795 PMC 458870126483812 · doi ↗ · pubmed ↗
- 3Marín Recinos MF, Pucker B. Supplementary data to evolutionary dynamics of the Proanthocyanidin biosynthesis gene LAR. 2025. 10.60507/FK 2/UKAAO 010.1186/s 12864-025-12429-5PMC 1283135541420151 · doi ↗ · pubmed ↗
- 4Quattrocchio F, Wing JF, Leppen HTC, Mol JNM, Koes RE. Regulatory genes controlling anthocyanin pigmentation are functionally conserved among plant species and have distinct sets of Target Genes.:17. 10.1105/tpc.5.11.149710.1105/tpc.5.11.1497 PMC 16038112271045 · doi ↗ · pubmed ↗
- 5Yang L, Zhang S, Chu D, Wang X. Exploring the evolution of CHS gene family in plants. Front Genet. 2024;15. 10.3389/fgene.2024.1368358.10.3389/fgene.2024.1368358 PMC 1109133438746055 · doi ↗ · pubmed ↗
- 6Davies KM, Jibran R, Zhou Y, Albert NW, Brummell DA, Jordan BR, et al. The evolution of flavonoid biosynthesis: A bryophyte perspective. Front Plant Sci. 2020;11. 10.3389/fpls.2020.00007.10.3389/fpls.2020.00007 PMC 701083332117358 · doi ↗ · pubmed ↗
- 7Khoo HE, Azlan A, Tang ST, Lim SM. Anthocyanidins and anthocyanins: colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr Res. 2017. 10.1080/16546628.2017.136177910.1080/16546628.2017.1361779 PMC 561390228970777 · doi ↗ · pubmed ↗
- 8Liu X, Lu Y, Yan M, Sun D, Hu X, Liu S, et al. Genome-Wide Identification, Localization, and expression analysis of Proanthocyanidin-Associated genes in brassica. Front Plant Sci. 2016;7. 10.3389/fpls.2016.01831.10.3389/fpls.2016.01831 PMC 514588128018375 · doi ↗ · pubmed ↗