Intestinal parasite prevalences in dogs and cats: a decade of retrospective data from a reference veterinary laboratory in Madrid, Spain
J. P. Barrera, A. Montoya, V. Marino, J. Sarquis, R. Checa, D. Carmena, E. Estévez-Sánchez, C. Gómez-Velasco, P. Moraleda, L. Cano, I. Fuentes, G. Miró

TL;DR
This study analyzed over 15,000 pet fecal samples in Madrid to track intestinal parasite trends in dogs and cats over a decade, finding high prevalence of zoonotic parasites like Giardia and Toxocara.
Contribution
The study provides the first comprehensive, decade-long analysis of intestinal parasite prevalence in pets in Madrid using modern diagnostic techniques.
Findings
Giardia duodenalis was the most prevalent parasite in dogs, while Cystoisospora spp. was most common in cats.
Puppies and kittens had higher infection rates, and seasonal patterns were observed for different parasites.
Infection rates increased in shelter animals and over time for Giardia, while Cystoisospora decreased.
Abstract
Spain’s recent abrupt rise in numbers of registered pet dogs and cats has intensified the need for proper animal health care, as 60% of infectious diseases are zoonotic. While pathogen detection has improved through advances in molecular techniques, pet owners often fail to adhere to veterinary guidelines, increasing infection risks. Among the diagnostic tools available, faecal analysis plays a key role in detecting zoonotic parasites such as Giardia duodenalis, Cryptosporidium spp. and Toxocara spp. This study was designed to assess intestinal parasite prevalence in dogs and cats along with epidemiological trends. Between 2013 and 2023, a total of 15,899 faecal samples from dogs and cats submitted to a reference laboratory of parasitology in Madrid (Spain) were analysed using Mini-FLOTAC®, merthiolate–iodine–formalin (MIF), Baermann–Wetzel (for lungworms and S. stercoralis when…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2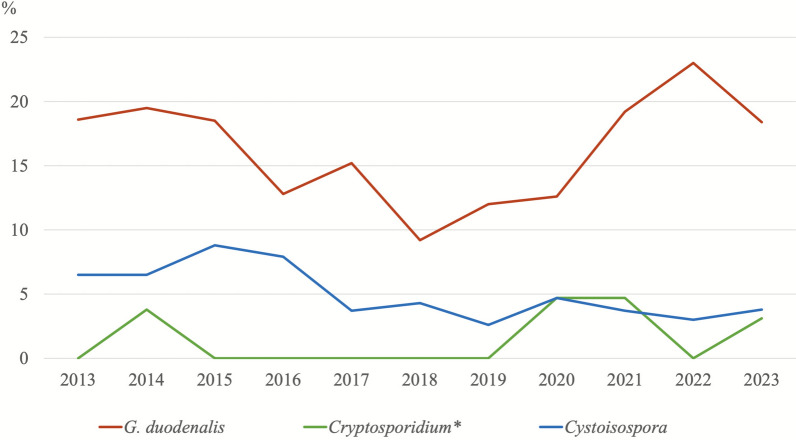 Figure 3
Figure 3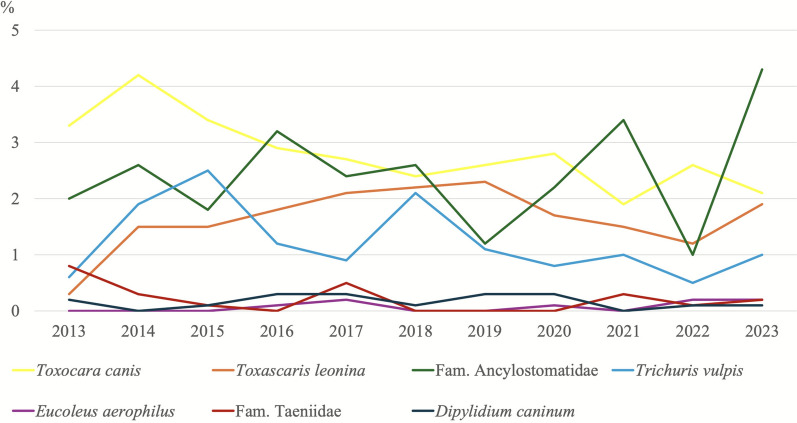 Figure 4
Figure 4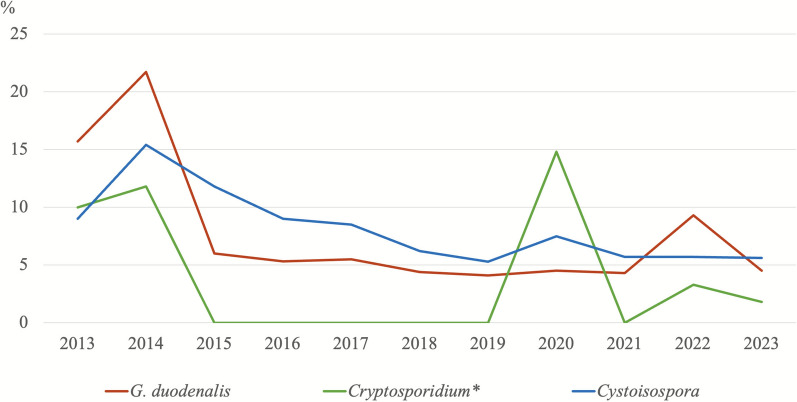 Figure 5
Figure 5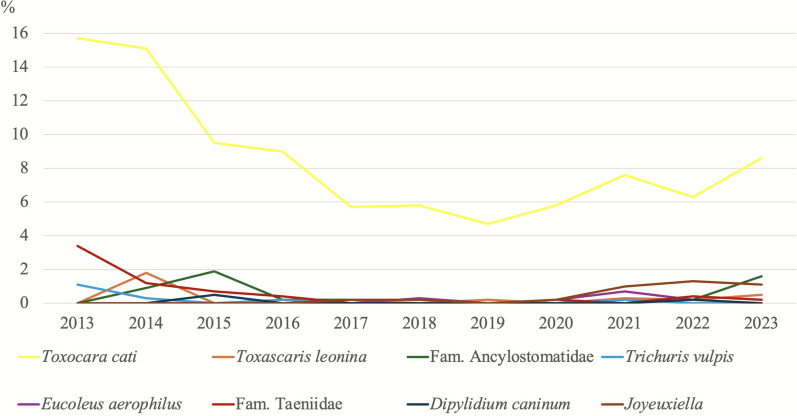 Figure 6
Figure 6 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasitic Infections and Diagnostics · Parasites and Host Interactions · Helminth infection and control
Background
Global figures of companion animals have shown a steady increase, especially since the 2019 coronavirus disease (COVID-19) pandemic. For instance, according to data published in 2023 [1], Spain had 9.3 million and 5.8 million registered dogs and cats, respectively, notably exceeding the population of children under the age of 15 years. This rising trend has been attributed to factors such as remote work, more time spent at home and changes in leisure activities during the pandemic [1]. Consequently, proper health check-ups in pets are crucial, as 60% of infectious diseases are zoonotic [2]. Advances in molecular techniques have improved the detection of pathogens, which can now be identified with greater precision [3].
Advances have been made both in the diagnosis of diseases in pets and in prevention, particularly through the implementation of healthcare plans that reinforce early diagnosis in small animal practices [4]. However, owners do not always adhere strictly to veterinarian’s recommendations, thereby increasing the risk of infection for both pets and people [5]. Among the tests usually included in healthcare plans, a coprological examination should always be undertaken owing to the ease and non-invasive nature of sample collection and the significance of the diagnosis obtained, as among the detectable infectious agents that affect dogs and cats, several zoonotic agents can be detected [6–9].
Faecal analysis is a routine procedure in many veterinary practices. However, it is often performed without sufficient attention paid to proper methodologies. While coprological tests are best performed by veterinarians or experienced veterinary technicians, this task is often delegated to the less experienced team members, usually with minimal instruction or emphasis on its significance [10]. The need should, therefore, be stressed to submit faecal samples to specialized veterinary reference parasitology laboratories, where staff are properly trained in these procedures.
Among the most relevant parasites that can be found in dog and cat faeces in Spain and other Mediterranean countries are protozoa (e.g. Giardia duodenalis, Cryptosporidium spp. and Cystoisospora spp.), nematodes (e.g. Toxocara canis, Toxocara cati, Toxascaris leonina, the Ancylostomatidae family, Trichuris vulpis, Eucoleus aerophilus and Strongyloides stercoralis) and cestodes (e.g. the Taeniidae family, Dipylidium caninum and Joyeuxiella spp.). In addition, some bronchopulmonary nematodes can be detected in faecal samples from carnivores (e.g. Angiostrongylus vasorum, Aelurostrongylus abstrusus and Troglostrongylus spp.) [11].
In general, animals infected with intestinal parasites show a wide range of clinical signs, ranging from subclinical infections, moderate to severe diarrhoea or, in extreme cases, even death [12]. Migrating parasites (such as Toxocara spp. and Ancylostoma spp.) are particularly worrying, as massive parasitism in animals under 1 year of age can lead to verminous pneumonia or intestinal intussusception [13]. Similarly, some of the zoonotic parasites mentioned can cause a similar range of clinical signs in humans, with more severe symptoms occurring in vulnerable populations, such as children aged under 5 years, the older population and immunocompromised individuals [6]. This is particularly true for infections caused by G. duodenalis and Cryptosporidium spp. Migratory parasites can lead to conditions such as cutaneous larva migrans (ancylostomids), larva currens (S. stercoralis) and ocular and visceral larva migrans (T. canis and T. cati) [14]. This highlights a need to detect intestinal and/or respiratory parasites, not only because of their impacts on public and animal health but also because prompt treatment of animals will prevent the environmental infection [15].
In the present study, we examined data derived from different coprological methods used on canine and feline faeces samples submitted to a veterinary parasitology reference laboratory by veterinary practices, breeding sites, stray cats and animal protection shelters located in the Community of Madrid (Spain). Our aims were to assess the prevalence of the most common intestinal parasite infections, to explore their potential associations with epidemiological variables and to examine trends over a 10-year period in dogs and cats.
Methods
Study design and sample collection
In this retrospective study, faecal samples from dogs and cats collected from 2013 to 2023 were analysed at PetParasiteLab, a reference laboratory for the diagnosis of infectious diseases in carnivores (Veterinary Faculty, Universidad Complutense de Madrid, Madrid, Spain). Only faecal samples from animals not previously registered as patients in the laboratory were included, corresponding to their first coprological analysis. These analyses were performed mainly as part of annual health check-ups covered by pet insurance, during the introduction of new animals into shelters or as control measures in cat colonies. This criterion was applied to avoid potential bias from prior administration of antiparasitic treatments that could alter true prevalence estimates. Samples collected during post-treatment follow-up were excluded, as persistent cyst excretion after effective therapy (e.g. in G. duodenalis infections) could lead to an overestimation of prevalence.
Veterinarians completed a clinical form (CRF) and submitted the faecal samples. Three consecutive daily samples per animal were analysed whenever available; in cases where this was not possible, a single sample was examined. Every CRF included the signalment of each animal, and the variables analysed were species (dog/cat), age (< 1 year/≥ 1 year), sex (male/female), origin (breeding dog/stray cats, owner and shelter), season (winter, spring, summer and autumn) and faecal consistency. The latter was determined by laboratory personnel using the Bristol Stool Chart (1–7) [16, 17], where a score of 1 corresponded to hard/dry stools and 7 indicated liquid diarrhoea.
Routine laboratory protocol for coprological analysis
Faecal samples were analysed using different techniques, depending on the type of coprological analysis requested, following internal validated standard operating procedures (SOPs). Samples were first macroscopically inspected. For the coprological analysis, Mini-FLOTAC^®^ was used to detect oocysts, eggs and/or larvae of the main intestinal parasites, and the merthiolate–iodine–formalin (MIF) method was carried out to detect G. duodenalis cysts.
When requested, an immunodiagnostic test was performed by direct immunofluorescence assay (DFA) to detect cysts of G. duodenalis and oocysts of Cryptosporidium spp. along with a molecular diagnosis to detect infection by G. duodenalis, Toxoplasma gondii, S. stercoralis (real-time polymerase chain reaction [PCR]) and Cryptosporidium spp. (conventional nested PCR). A G. duodenalis infection was considered present when at least one positive result was obtained through the MIF and/or DFA methods. Likewise, Cryptosporidium infection was recorded on the basis of a positive DFA result. Molecular techniques for G. duodenalis and Cryptosporidium spp. were employed only for confirmation and diagnostic support and not as the main diagnostic method.
The Baermann–Wetzel method was also used to detect lungworms and S. stercoralis when there was clinical suspicion on the basis of clinical signs such as coughing and/or lifestyle factors (e.g. habitat, aptitude, etc.). In consequence, the number of samples analysed with this technique was lower.
Flotation method (Mini-FLOTAC®)
The flotation method requiring a Mini-FLOTAC^®^ device was used on cat and dog faecal samples according to the manufacturer’s instructions [18, 19].
Merthiolate–iodine–formalin (MIF) method
This procedure is particularly useful to detect G. duodenalis cysts in faecal concentrates prepared from pooled samples collected over 3–5 consecutive days from the host. This method [20] is useful to partially distinguish between active cysts (Fig. 1) and degenerated ones on the basis of their morphology [21].Fig. 1Giardia duodenalis cyst (15 × 12 μm) detected by the MIF method. 1000×
Direct immunofluorescence assay (DFA)
The commercial Crypto/Giardia Cel IF^®^ Kit (CeLLabs, Brookvale, Australia) was used following the manufacturer’s manual. The resulting slides were examined under a Nikon Eclipse Ci-S fluorescence microscope (Nikon, Tokyo, Japan) at 400× magnification. Round to oval structures of appropriate size (G. duodenalis cysts: 8–12 μm; Cryptosporidium oocysts: 4–6 μm) visible as bright apple-green fluorescence were identified as positive (Fig. 2).Fig. 2Giardia duodenalis cyst (narrow-headed white arrow) and Cryptosporidium spp. oocyst (wide-headed white arrow) detected by the DFA method. 400×
Baermann–Wetzel method
This method relies on the movement of living nematode larvae from faeces into the surrounding water, where they are collected in a funnel. A plastic funnel was attached to a 10-cm-long piece of soft silicone tube, which was sealed with a pinch clamp positioned at an angle to the tube’s tip. Water was added to the funnel together with a surfactant solution, and 5 g of faecal sample was placed on a gauze and wrapped up. The funnel was left at room temperature for 8 h, allowing the larvae to leave the faeces and settle at the tip of the tube by sedimentation. Next, the pinch clamp was opened and the liquid drained into a 10-ml tube. This tube was then centrifuged at 1500 rpm for 10 min. Next, the supernatant was discarded and the sediment examined under a microscope [11]. Pulmonary nematodes and compatible S. stercoralis larvae were identified on the basis of keys [22, 23].
Faecal DNA extraction and purification
DNA was extracted from 200 mg of each concentrated faecal sample using the QIAamp DNA Stool Mini Kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol. The samples, mixed with InhibitEX buffer, were incubated for 10 min at 95 °C. The purified DNA was then eluted in 200 μl of PCR-grade water and stored at 4 °C until further PCR analysis.
PCR methods
For the detection of G. duodenalis, T. gondii and S. stercoralis, real-time PCR (qPCR) assays were used, targeting highly conserved genes such as 18S rRNA gene for G. duodenalis and S. stercoralis, and the B1 gene for T. gondii, allowing for rapid and sensitive identification. For Cryptosporidium spp., a nested PCR was performed to amplify a 587 bp fragment of the 18S rRNA gene, which enhances diagnostic sensitivity in low-load infections. Primer sequences, amplicon sizes, PCR methods and the references are presented in Table 1. Table 1. Oligonucleotides and PCR conditions used for the molecular detection of intestinal parasitesTarget genePrimers (nucleotide sequence 5′-3′)Amplicon sizeMethodReferenceG. duodenalis**18S rRNAGd-80F: 5′-GACGGCTCAGGACAACGGTT-3′Gd-127R: 5′-TTGCCAGCGGTGTCCG-3′62 bpReal-time PCR[24]Cryptosporidium spp.18S rRNACR-P1: 5′-CAGGGAGGTAGTGACAAGAA-3′CR-P2: 5′-TCAGCCTTGCGACCATACTC-3′CR-P3: 5′-ATTGGAGGGCAAGTCTGGTG-3′CPB-DIAGR: 5′-TAAGGTGCTGAAGGAGTAAGG-3′652 bp652 bpNested PCR[25]T. gondii**B1 gene23-mer 5′-GGAGGACTGGCAACCTGGTGTCG-3′25-mer 5′-TTGTTTCACCCGGACCGTTTAGCAG-3′126 bpReal-time PCR[26]S. stercoralis**18S rRNAStro18S-1530F: 5′-GAATTCCAAGTAAACGTAAGTCATTAGC-3′Stro18S-1630R: 5′-TGCCTCTGGATATTGCTCAGTTC-3′101 bpReal-time PCR[27]rRNA ribosomal ribonucleic acidbp base pair
Sequencing analyses
PCR products were sequenced in both directions using the corresponding internal primer sets described above, employing BigDye^™^ chemistry and an ABI 3730xl sequencer (Applied Biosystems^®^). Raw sequencing data from both forward and reverse directions were analysed using Chromas Lite version 2.1. Nucleotide sequences were compared with reference sequences from the National Center for Biotechnology Information (NCBI) database using the Basic Local Alignment Search Tool (BLAST) tool. The resulting DNA consensus sequences were aligned with reference sequences using MEGA 11 software to confirm species identity.
Phylogenetic relations between the Cryptosporidium sequences identified in this study and those retrieved from the NCBI repository were analysed using the neighbour-joining (NJ) method in MEGA 11 [28, 29]. Genetic distances were calculated with the Kimura 2-parameter model, while rate variation among sites was modelled using a Gamma distribution (shape parameter = 2).
Statistical analysis
To assess associations between the variables examined and infections by the different species of the parasite, the chi-squared test was employed. All statistical tests were conducted using the SPSS Statistics package version 17.0 (IBM, Chicago, IL, USA). Significance was set at a confidence level of 95% (P < 0.05).
Results
Prevalence rates
Over a 10-year period (2013–2023), 15,889 faecal samples (10,813 from dogs and 5086 from cats) were analysed along with several epidemiological variables (Table 2). The global prevalence of detected parasites is presented in Table 3. Using previously described coprological methods, 26% (2801/10,813) of the dog samples and 21.4% (1090/5086) of the cat samples tested positive for at least one parasite. Table 2. Distribution of dogs and cats by sex, age, origin, faecal consistency, season and year of the analysisVariableDogs (n = 10,813)Cats (n = 5086)Total (n = 15,889)n%n%n%Age< 1 year4894.52765.47654.8≥ 1 year416238.5105320.7521532.8Total465113295980SexMale468243.3222343.7690543.5Female480644.4235746.3716345.1Total9488458014,068OriginBreeders/stray cats229421.276215.0305619.2Owned297727.5107021.0404725.5Shelters539949.9325464.0865354.5Total10,670508615,756Faecal consistency1–2610.61943.82551.63–4282926.2141427.8424326.75–7188417.43296.5221313.9Total4774193738.1671142.2SeasonWinter267424.7109521.5376923.7Spring296927.5115722.7412626.0Summer209119.3143828.3352922.2Autumn307928.5139627.4447528.1Total10,813508615,899Years20136666.2891.77554.820147957.43376.611327.220157266.74318.511577.32016119011.056811.2175811.12017110710.254310.7165010.42018116310.864312.6180611.4201910039.351410.115179.620209428.74669.214088.92021116410.857911.4174311.02022109910.24739.3157210.020239588.94438.714018.9Total10,813508615,889n number of animalsFaecal consistency (Bristol Stool Chart): scale from 1 to 7; where 1–2 indicates very firm stools, 3–4 normal stools and 5–7 soft to liquid stoolsTotal may vary owing to missing data in some categoriesTable 3Global prevalence of intestinal parasites detected in canine and feline faecal samples analysedDogs (N = 10,813)Cats (N = 5086)Total (N = 15,889)n%95% CIn%95% CIn%95% CIProtozoaG. duodenalis173216.015.3–16.73326.55.8–7.2206413.012.5–13.5Cryptosporidium spp.^^191.80.99–2.6153.51.7–5.2342.31.5–3.0Cystoisospora* spp.5264.94.4–5.33957.87.1–8.59215.85.4–6.2NematodaToxocara canis/cati2962.72.3–3.03887.66.9–8.36844.34.0–4.6Toxascaris leonina1841.71.4–1.9130.30.1–0.41971.21.0–1.4Fam. Ancylostomatidae2672.52.2–2.7230.50.3–0.72901.81.6–2.0Trichuris vulpis1331.20.9–1.440.10.0–0.21370.90.8–1.0Eucoleus aerophilus80.10.0–0.280.20.1–0.3160.10.0–0.1Angiostrongylus vasorum^♦^73.30.9–5.7NDNDNDNDNDNDAelurostrongylus abstrusus^♦^NDNDND4213.910.0–17.8NDNDNDTroglostongylus spp.^♦^NDNDND20.70.0–1.6NDNDNDStrongyloides stercoralis^♦^94.21.5–6.9NDNDNDNDNDNDCestodaFam. Taeniidae190.20.1–0.3160.30.2–0.5350.20.1–0.3Dipylidium caninum180.20.1–0.330.10.0–0.2210.10.0–0.1Joyeuxiella spp.NDNDND200.40.2–0.6200.10.0–0.1^^number of samples analysed for Cryptosporidium spp. by DFA: dogs: 1046; cats: 428; total: 1474N*: total number of samples from dogs and catsn: number of positive animals^♦^number of samples analysed for lungworms and S. stercoralis by the Baermann–Wetzel method: dogs: 213; cats: 303; total: 516CI confidence interval, ND not detected
The most prevalent parasites were G. duodenalis (16.0%; 1732/10,813)) infecting dogs and Cystoisospora spp. (7.8%; 395/5086) and T. cati (7.6%; 388/5086) infecting cats. The least prevalent parasites were E. aerophilus (0.1%; 8/10,813) in dogs and D. caninum (0.1%; 3/5086) in cats.
Of the 3891 samples testing positive in dogs and cats, 531 (13.6%) were co-infections with more than one parasite. The most common co-infections were G. duodenalis/Cystoisospora spp. (30.5%; 162/531) in dogs, followed by Cystoisospora spp./T. cati (13.4%; 71/531) and G. duodenalis/T. cati (13.2%; 70/531) in cats. No oocysts compatible with T. gondii or DNA were detected in the faecal samples from 108 cats analysed by qPCR.
Associated epidemiological variables
Our statistical analysis results regarding associations between infection by each parasite and epidemiological variables are presented in Tables 4 and 5. Table 4. Intestinal parasites detected in dogs by sex, age, origin, faecal consistency and season of the analysisVariableG. duodenalisCryptosporidium spp. % (+/n)^^Cystoisospora spp.T. canis**T. leoninaFam. AncylostomatidaeT. vulpisE. aerophilusFam. TaeniidaeD. caninumA. vasorum* % (+/n)^^S. stercoralis % (+/n)^^Age < 1 (n = 489)31.7 (155) ^a^10 (6/60) ^a^14.5 (71)^a^5.5 (27) ^a^1.4 (7)2.2 (11)0.8 (4)0 (0)0 (0)0 (0)0 (0/5)40.0 (2/5) ^a^ ≥ 1 (n = 4162)19.7 (819) ^b^1.0 (7/685) ^b^3.8 (157)^b^1.4 (59) ^b^0.9 (38)1.9 (77)0.5 (22)0.02 (1)0.3 (12)0.02 (1)0 (0/26)0 (0/26) ^b^Total (n = 4651)20.9 (974)1.7 (13/745)4.9 (228)1.8 (86)1.0 (45)1.9 (88)0.6 (26)0.02 (1)0.3 (12)0.02 (1)0 (0/31)6.5 (2/31)P-value** < 0.001**** < 0.001**** < 0.001**** < 0.0010.260.530.410.10.230.73– < 0.001SexMale (n = 4682)15.8 (738)2.9 (14/488) ^a^4.8 (223)3.1 (144)1.9 (87)2.2 (102)1.3 (62)0 (0)0.2 (8)0.3 (12)3.8 (4/106)1.9 (2/104)Female (n = 4806)15.2 (729)0.9 (4/458) ^b^5.2 (249)2.6 (125)1.7 (83)2.8 (133)1.2 (57)0.02 (1)0.2 (8)0.1 (5)3.1 (3/97)2.1 (2/95)Total (n = 9488)15.5 (1467)1.9 (18/946)5.0 (472)2.8 (269)1.8 (170)2.5 (235)1.3 (119)0.01 (1)0.2 (16)0.2 (17)3.4 (7/203)4.4 (9/203)P-value0.420.0240.30.160.630.0650.540.450.960.080.790.92OriginBreeding (n = 2294)22.5 (517) ^a^0.2 (1/482) ^b^4.7 (107)0.7 (16) ^b^0.3 (7) ^b^1.2 (27)0.1 (3) ^b^0 (0)0.1 (2)0 (0) ^b^0 (0/7)0 (0/7)Owned (n = 2977)15.8 (470)3.3 (16/480) ^a^5.3 (158)2.9 (87)1.0 (29) ^b^1.6 (49)b0.5 (16) ^b^0.03 (1)0.1 (3)0.4 (11) ^a^3.4 (7/203)4.4 (9/203)Sheltered (n = 5399)12.8 (693) ^b^1.4 (1/74)4.8 (258)3.6 (192) ^a^2.7 (148) ^a^3.5 (189) ^a^2.0 (110) ^a^0.13 (7) ^a^0.3 (14) ^a^0.1 (7)0 (0/3)0 (0/3)Total (n = 10,670)15.7 (1680)1.7 (18/1036)4.9 (523)2.8 (295)1.7 (184)2.5 (265)1.2 (129)0.07 (8)0.2 (19)0.2 (18)3.3 (7/213)4.2 (9/213)P-value < 0.001**** < 0.0010.5 < 0.001**** < 0.001**** < 0.001**** < 0.0010.10.130.0030.840.8Faecal consistency1–2 (n = 61)6.6 (4) ^b^0.0 (0/)0 (0)0 (0)0 (0)0 (0)0 (0)0 (0)0 (0)0 (0)0 (0/2)0 (0/2)3–4 (n = 2829)16.2 (458) ^b^1.3 (8/632)2.3 (66) ^b^1.1 (31) ^b^0.8 (23) ^b^2.0 (57)0.6 (17)0.1 (2)0.1 (4)0.07 (2)0 (0/58)3.4 (2/58)5–7 (n = 1884)22.6 (425) ^a^2.7 (10/377)6.9 (130) ^a^2.9 (55) ^a^1.8 (34) ^a^2.7 (50)0.7 (13)0.1 (2)0.1 (1)0 (0)3.8 (2/52)3.8 (2/52)Total (n = 4774)18.6 (887)1.7 (18/)4.1 (196)1.8 (86)1.2 (57)2.2 (107)0.6 (30)0.1 (4)0.1 (5)0.04 (2)1.8 (2/112)3.6 (4/112)P-value < 0.0010.145 < 0.001**** < 0.0010.0060.170.760.890.630.50.30.95SeasonWinter (n = 2674)18.1 (483) ^a^3.9 (12/310) ^a^3.7 (100) ^b^3.0 (79)1.7 (46)2.4 (63)1.5 (39)0.0 (1)0.1 (2)0.1 (3)5.8 (3/52)3.8 (2/52)Spring (n = 2969)16.9 (501)0.3 (1/363) ^b^4.3 (129)2.0 (59) ^b^1.4 (41)2.0 (59)1.2 (35)0.1 (3)0.2 (7)0.3 (10)3.9 (2/51)5.9 (3/51)Summer (n = 2091)12.6 (263) ^b^0.7 (1/135)5.4 (112)2.9 (61)1.5 (32)2.4 (50)0.8 (16) ^b^0.0 (0)0.2 (5)0.1 (3)0 (0/56)7.1 (4/56)Autumn (n = 3079)15.8 (485)2.1 (5/238)6.0 (185) ^b^3.2 (97)2.1 (65) ^a^3.1 (95) ^a^1.4 (43)0.1 (4)0.2 (5)0.1 (2)3.7 (2/54)0 (0/54) ^b^Total (n = 10,813)11.6 (1249)0.7 (7/1046)3.9 (426)2.0 (217)1.3 (138)1.9 (204)0.9 (94)0.1 (7)0.2 (17)0.1 (15)1.9 (4/213)3.3 (7/213)P-value < 0.001 < 0.001**** < 0.001**** < 0.001**** < 0.001**** < 0.001**** < 0.0010.230.350.020.0580.13Bold indicates statistical significancen: number of animals +: number of positive animalsFaecal consistency: scale from 1 to 7, where 1–2 indicates very firm stools, 3–4 normal stools and 5–7 soft to liquid stools^a^adjusted residual > 2: statistically significantly higher than expected^b^adjusted residual ≤ 2: statistically significantly lower than expectedTable 5Intestinal parasites detected in cats by sex, age, origin, faecal consistency and season of the analysisVariable*G. duodenalisCryptosporidium* spp. % (+/n)^^Cystoisospora spp.T. cati**T. leoninaFam. AncylostomatidaeT. vulpisE. aerophilusFam. TaeniidaeD. caninumJoyeuxiella* spp.A. abstrusus % (+/n)^^Troglostrongylus spp. % (+/n)^^Age < 1 (n = 276)11.6 (32)8.0 (2/25)16.3 (45)^a^11.2 (31)^a^0.4 (1)0 (0)0 (0)0.7 (2)^a^0 (0)0 (0)0 (0)25.0 (3/12)0 (0/12) ≥ 1 (n = 1053)13.2 (139)3.4 (11/321)2.6 (27)^b^4.4 (46)^b^0.5 (5)0.9 (9)0.09 (1)0 (0)^b^0.3 (3)0 (0)0.2 (2)7.6 (8/105)0 (0/105)Total (n = 1329)12.9 (171/)3.8 (13/346)5.4 (72)5.8 (77)0.5 (6)0.7 (9)0.08 (1)0.2 (2)0.2 (3)0 (0)0.2 (2)9.4 (11/117)0 (0/117)P-value0.470.24** < 0.001**** < 0.0010.80.120.60.0060.37–0.4680.05–SexMale (n = 2223)7.1 (158)2.8 (6/212)8.2 (182)8.8 (196)^a^0.2 (5)0.3 (7)0.04 (1)0.22 (5)0.1 (2)^b^0.09 (2)0.6 (14)13.1 (19/145)0 (0/145)Female (n = 2357)6.2 (147)4.9 (9/184)7.6 (179)7.1 (168)^b^0.3 (8)0.5 (12)0.08 (2)0.08 (2)0.4 (10)^a^0.04 (1)0.3 (6)13.1 (17/130)1.5 (0/130)Total (n = 4580)6.7 (305)3.8 (15/396)7.9 (3)7.9 (364)0.3 (13)0.4 (19)0.07 (3)0.15 (7)0.3 (12)0.07 (3)0.4 (20)13.1 (36/275)0.7 (2/275)P-value0.230.280.450.030.460.3060.590.220.030.530.0540.990.133OriginStray cats (n = 762)12.7 (97)^a^1.9 (4/214)1.7 (13) ^b^2.6 (20)^b^0.7 (5)^a^0.8 (6)0 (0)0 (0)0.1 (1)0 (0)0.1 (1)5.6 (5/90)^b^0 (0/90)Owned (n = 1070)6.1 (65)3.9 (6/153)5.3 (57)^b^7.3 (78)0.3 (3)0.4 (4)0.09 (1)0.3 (3)0.2 (2)0 (0)1.0 (11)^a^17.7 (29/164)0.6 (1/164)Sheltered (n = 3254)5.2 (170)^b^8.2 (5/41)^a^10.0 (325) ^a^8.9 (290)^a^0.2 (5)0.4 (13)0.09 (1)0.2 (5)0.4 (13)0. 09 (3)0.3 (9)^b^16.3 (8/49)^a^2.0 (1/49)Total (n = 5086)6.5 (332)3.5 (15/428)7.8 (395)7.6 (388)0.3 (13)0.5 (23)0.08 (4)0.2 (8)0.3 (16)0.06 (3)0.4 (21)13.9 (42/303)0.7 (2/303)P-value < 0.0010.056 < 0.001**** < 0.0010.0460.3240.70.320.350.430.00160.0240.36Faecal consistency1–2 (n = 194)3.1 (6)12.5 (1/8)9.8 (19) ^a^9.3 (18)^a^0 (0)0.5 (1)0 (0)0.5 (1)0 (0)0 (0)1.5 (3)23.5 (4/17)0 (0/17)3–4 (n = 1414)5.7 (81)3.1 (1/32)4.1 (58)5.5 (78)0.3 (4)0.4 (6)0.07 (1)0.1 (2)0.1 (2)0 (0)0.7 (10)10.7 (15/140)0 (0/140)5–7 (n = 329)13.7 (45)^a^0 (0/4)2.7 (9)5.5 (18)0.3 (1)0.3 (1)0 (0)0.6 (2)0.3 (1)0 (0)1.2 (4)21.1 (4/19)5.3 (1/19)Total (n = 1937)6.8 (132)4.5 (2/44)4.4 (86)5.9 (114)0.3 (5)0.4 (8)0.05 (1)0.3 (5)0.2 (3)0 (0)0.9 (17)13.1 (23/176)0.6 (1/176)P-value < 0.0010.47 < 0.0010.1060.750.9280.830.240.67–0.3860.1840.015SeasonWinter (n = 1095)10.4 (114)^a^2.8 (5/179)4.3 (47)^b^5.5 (60)^b^0.8 (9)^b^0.6 (7)0.2 (2)0.2 (2)0 (0)^b^0.2 (2)0.5 (5)23.9 (11/46)0 (0/46)Spring (n = 1157)4.0 (46)1.2 (1/83)8.8 (102)7.0 (81)0.1 (1)0.4 (5)0 (0)0.2 (2)0.2 (2)0 (0)0.3 (4)15.4 (12/78)1.3 (1/78)Summer (n = 1438)5.6 (81)^b^2.7 (1/37)8.9 (128)8.7 (125)0.1 (2)0.2 (3)0.1 (1)0.2 (3)0.3 (4)0.1 (1)0.1 (2)17.2 (10/58)1.7 (1/58)Autumn (n = 1396)6.5 (91)6.2 (8/129)8.5 (118)8.7 (122)0.1 (1)0.6 (8)0.1 (1)0.1 (1)0.7 (10)^a^0 (0)0.7 (10)^a^7.4 (9/121)0 (0/121)Total (n = 5086)4.3 (218)2.3 (10/428)6.8 (348)6.4 (328)0.1 (4)0.3 (16)0 (1)0.1 (6)0.3 (16)0 (1)0.3 (16)10.2 (31/303)0.7 (2/303)P-value < 0.0010.03 < 0.001**** < 0.001**** < 0.0010.0530.070.60.009 < 0.0010.0250.001**0.46Bold indicates statistical significance^^the total number of samples analysed for Cryptosporidium spp. analyses differs from that for the other parasites. The number of animals analysed is indicated as n in the format (+/n) for each variable+: number of positive animalsn*: number of animalsFaecal consistency: scale from 1 to 7, where 1–2 indicates very firm stools, 3–4 normal stools and 5–7 soft to liquid stools^a^adjusted residual > 2: statistically significantly higher than expected^b^adjusted residual ≤ 2: statistically significantly lower than expected
Dogs
Young dogs (< 1 year) showed a significantly higher risk of infection by Cystoisospora spp. (χ^2^ = 108,19; *df * = 1; P = < 0.001), G. duodenalis (χ^2^ = 37.96; df = 1; P = < 0.001), Cryptosporidium spp. (χ^2^ = 25.78; df = 1; P = < 0.001), T. canis (χ^2^ = 40.35; df = 1; P = < 0.001) and S. stercoralis (χ^2^ = 11.19; df = 1; P = < 0.001). The risk of G. duodenalis infection was higher in dogs from breeding (χ^2^ = 114.07; df = 2; P = < 0.001), while Cryptosporidium spp. (χ^2^ = 13.83; df = 2; P = < 0.001) and D. caninum (χ^2^ = 11.61; df = 2; P = 0.003) were more frequently detected in owned dogs. Among shelter dogs, a higher positivity rate was recorded for T. canis (χ^2^ = 48.90; df = 2; P = < 0.001), T. leonina (χ^2^ = 69.93; df = 2; P = < 0.001), ancylostomids (χ^2^ = 49.07; df = 2; P = < 0.001) and T. vulpis (χ^2^ = 64.60; df = 2; P = < 0.001). Cystoisospora spp. (χ^2^ = 60.25; *df *= 2; P = < 0.001), G. duodenalis (χ^2^ = 36.91; df = 2; P = < 0.001), T. canis (χ^2^ = 22.95; df = 2; P = < 0.001) and T. leonina (χ^2^ = 10.27; df = 2; P = 0.006) were more frequently detected in samples from dogs with soft stools (consistency scores of 5–7) than normal stools (scores of 1–4). Regarding seasonality, G. duodenalis was more prevalent in the winter (χ^2^ = 238.80; df = 3; P = < 0.001), whereas Cystoisospora (χ^2^ = 47.95; df = 3; P < 0.001), T. leonina (χ^2^ = 22.34; df = 3; P = < 0.001) and ancylostomids (χ^2^ = 29.24; df = 3; P = < 0.001) were more prevalent in the autumn.
Cats
A significantly higher-than-expected prevalence of T. cati infection was observed in male cats (χ^2^ = 4.48; df = 1; P = 0.03). Kittens (< 1 year) had a higher risk of infection by Cystoisospora spp. (χ^2^ = 80.90; df = 1; P = < 0.001) and T. cati (χ^2^ = 18.99; *df *= 1; P < 0.001). Cats from feline colonies were found to be more frequently infected by G. duodenalis (χ^2^ = 57.60; df = 2; P < 0.001) and T. leonina (χ^2^ = 6.16; df = 2; P = 0.046), while Cystoisospora spp. (χ^2^ = 67.42; df = 2; P = < 0.001) and T. cati (χ^2^ = 35.47; df = 2; P = < 0.001) infections were more prevalent in shelter cats. In cats with soft stools (score 5–7), infection by G. duodenalis was more prevalent (χ^2^ = 31.15; df = 2; P < 0.001), whereas in cats with hard dry stools (score 1–2), infections by Cystoisospora spp. (χ^2^ = 15.65; df = 2; P = < 0.001) were more prevalent. As in the case of dogs, the prevalence of G. duodenalis infection was higher in winter (χ^2^ = 122.04; df = 3; P = < 0.001) and that of Joyeuxiella spp. (χ^2^ = 9.36; df = 3; P = 0.025) was higher in autumn.
Trends in parasite infection prevalences over the years
Figures 3 and 4 show the prevalence data of protozoan and helminth infections detected in canine samples throughout the study period, while Figs. 5 and 6 present the corresponding prevalence data obtained from feline samples.Fig. 3. Trends in the percentages of protozoan parasites detected in dogs over time. ^^The number of samples analysed for the diagnosis of Cryptosporidium spp. is not the same as for the other parasites. Details are presented in Table 3Fig. 4. Trends in the percentages of helminth parasites detected in dogs over timeFig. 5Trends in the percentages of protozoan parasites detected in cats over time. ^^The number of samples analysed for the diagnosis of Cryptosporidium spp. is not the same as for the other parasites. Details are presented in Table 3Fig. 6. Trends in the percentages of helminth parasites detected in cats over time
In dogs, the prevalence of G. duodenalis infection was higher than expected in 2021, 2022 and 2023 (χ^2^ = 137.08; df = 10; P = < 0.001), whereas the prevalence of Cystoisospora spp. showed a decreasing trend in 2021 and 2022 (χ^2^ = 86.14; df = 10; P = < 0.001). In the case of helminth infections, only those caused by ancylostomids showed a higher-than-expected prevalence in 2023 in both dogs (χ^2^ = 39.25; *df *= 10; P = < 0.001) and cats (χ^2^ = 42.47; df = 10; P = < 0.001).
Joyeuxiella infection in cats showed a clear increasing trend in the recent years (from 2020 to the present).
Discussion
This is the largest epidemiological survey of endoparasites in dogs and cats based on coprological data conducted in Spain. Our results provide valuable insight into the prevalence and epidemiological trends of intestinal parasites in dogs and cats living in Madrid over a 10-year period. The significant variations in parasite prevalences, co-infections and associated epidemiological factors identified here contribute to a better understanding of intestinal parasite infections in companion animals.
The overall prevalence of parasites in dogs (26%) was higher than in cats (21.4%), consistent with findings from other large-scale studies [30–32]. These differences may be attributed to factors such as study area, diagnostic techniques and population characteristics (e.g. age and housing conditions) [33]. However, it is important to consider the limitations of coprological methods, particularly their sensitivity for detecting parasites with intermittent shedding patterns which may lead to underdiagnoses [34].
In dogs, the most prevalent parasite detected was G. duodenalis (16.0%), whereas in cats, Cystoisospora spp. (7.8%) and T. cati (7.6%) were the most frequently detected. These findings are in line with data from previous studies indicating that G. duodenalis is a widespread protozoan parasite in dogs, particularly in environments with high animal densities such as shelters and breeding facilities [35]. Similarly, Cystoisospora spp. is known to be a prevalent parasite in feline populations, especially among kittens, cats with gastrointestinal disorders and shelter animals [32, 36, 37]. The lower detection rate of helminth parasites may be related to deworming protocols. These are routinely used in pets but do not typically include chemicals that prevent infection by protozoa. Veterinarians must therefore interpret the results of a coprological analysis and decide whether treatment against protozoal infections is necessary in otherwise healthy animals. Emphasis should be placed on preventing protozoan infections through other strategies such as proper hygiene and faecal removal from the environment [38].
Interestingly, the less prevalent parasites detected in dogs and cats were E. aerophilus (0.1% and 0.2%, respectively), in agreement with reports of studies in dogs samples [39, 40]. In cats, the least prevalent parasite was D. caninum (0.1%). This suggests that these parasites have low transmission rates in this species. In the case of D. caninum in cats, this could be related to effective flea control measures. Alternatively, D. caninum prevalence may be underestimated because of detection difficulties arising from the intermittent shedding of proglottids in faeces and the rupture of the ovigerous capsule, as the eggs may be indistinguishable from those of other cestode species [41].
The most common co-infections observed in dogs were G. duodenalis/T. canis, while in cats, they were Cystoisospora spp./T. cati and Cystoisospora spp./G. duodenalis. These results highlight G. duodenalis as one of the most prevalent parasites in companion animals in the last decade, with significance implications not only in mono-infections but also in co-infections with other intestinal parasites. Controlling this parasite is crucial, as a positive result for another parasite could mask the presence of this potentially zoonotic pathogen, posing public health risks, especially in households where dogs and cats live with people at risk (e.g. children, pregnant women, immunocompromised individuals or elderly people). Our data indicate that specific parasites co-exist in the same hosts, likely owing to similar transmission routes or overlapping environmental conditions. The relatively low prevalence of co-infections, however, suggests that mono-infections were common in our study sample, in line with the findings of previous studies in the same area [42].
Prevalence data for lung worms detected by the Baermann–Wetzel method should be interpreted with caution, as the total sample size differs from that of other analyses. This is because the Baermann–Wetzel method is not a routine procedure and was only requested when there was specific suspicion of infection. In addition, the absence of T. gondii in all feline faecal samples should be underscored although expected, given the results of other authors [43]. This could be owing to the limited shedding period of T. gondii oocysts in cats or a low environmental contamination level during the sampling period. Further work is needed with a larger sample size and different diagnostic methods for a more comprehensive analysis of T. gondii prevalence in cats and the importance of its transmission to humans [44].
The analysis of the epidemiological variables studied served to identify significant correlations between host characteristics and the parasite detected. Age played a key role in infection risk, such that puppies and kittens under 10 year of age were more susceptible to infections, such as Cystoisospora spp., G. duodenalis, Cryptosporidium spp., T. canis and S. stercoralis infection in dogs and Cystoisospora spp. and T. cati infection in cats. This higher susceptibility is most likely due to an immature immune system and increased exposure to parasites in crowded environments [45, 46]. We should also consider the role of transplacental transmission in T. canis and lactogenic transmission in both T. canis and T. cati, as these are critical transmission routes in young animals [11].
Housing conditions also influenced infection risk. Dogs from breeders had a higher prevalence of G. duodenalis infection, but in this case, owned animals showed higher rates than those from the shelter. This could be because owned puppies were often brought in for deworming prior to vaccination and frequently originated from breeding facilities where prevalence is higher. In contrast, the shelter submitting samples implemented very strict control measures, both for the animals and the environment, which likely limited parasite transmission. Shelter dogs were more frequently infected by T. canis, T. leonina, ancylostomids, T. vulpis and E. aerophilus. In contrast, owned dogs showed a higher prevalence of Cryptosporidium spp. and D. caninum infection, most likely because of ease of direct transmission due to closer contact between animals and even humans. As for D. caninum infection, improved household conditions help maintain the life cycle of fleas, facilitating the persistence of eggs, larval and pupal stages of the fleas in the environment and, although most pet owners deworm their pets, they do so on the basis of very different criteria and often irregularly [47, 48]. Similarly, cats from colonies featured higher infection rates of G. duodenalis and T. leonina, whereas shelter cats had a greater prevalence of Cystoisospora spp. and T. cati. This underscores the impact of environmental factors, population density and the lack of awareness of pet owners of parasite transmission [49] and highlights the importance of parasite control in pets owing to zoonotic risks [50, 51].
Faecal consistency (according to the Bristol Stool Chart) was also found to correlate with infection prevalence. Hence, dogs with soft stools (scores 5–7) had higher detection rates of Cystoisospora spp., G. duodenalis, T. canis and T. leonina, while in cats, G. duodenalis was more common in soft stools and Cystoisospora spp. and T. cati were more frequent in hard dry faeces samples (scores 1–2). This suggests that certain parasites contribute to gastrointestinal disturbances, while others may cause subclinical disease [52]. Accordingly, we would recommend routine coprological tests on ‘clinically healthy’ animals with faeces with normal characteristics, especially in cats, as they usually defecate in litter boxes and depending on the composition of the litter the organoleptic properties of the faeces may be modified making their consistency classification more difficult.
Seasonality in prevalence was observed, G. duodenalis infection being more prevalent in winter and T. leonina and ancylostomids in autumn in both dogs and cats. The higher winter prevalence of G. duodenalis may be explained by increased indoor housing, leading to higher transmission rates, whereas autumn peaks in helminths could be related to increased environmental contamination in this season because the climate conditions are more favourable for the survival of infective larval stages [53].
Longitudinal analysis of data collected from 2013 to 2023 revealed fluctuations in parasite prevalences. In dogs, G. duodenalis showed a higher-than-expected increase in 2021–2023, suggesting a rising trend, possibly attributable to increased awareness and improved diagnostic techniques compared with early years [54]. Conversely, Cystoisospora spp. in dogs showed a decreasing trend in 2021–2022. This may be attributed to improved hygiene practices and management strategies in shelter animals [55]. Helminth infections, particularly ancylostomids infections, experienced a significant increase in 2023 in both dogs and cats. This could suggest their re-emergence, possibly due to environmental or climate factors [56]. Notably, Joyeuxiella spp. infection in cats showed a consistent increase from 2020 to the present, warranting further investigation into their epidemiology and potential risk factors for transmission.
Conclusions
This study provides updates and comprehensive data regarding the prevalence of endoparasites and their co-infections and epidemiological trends in dogs and cats over a decade. G. duodenalis emerged as the most prevalent parasite detected in both dogs and cats living in Madrid. These findings highlight the importance of regular parasitological surveillance, targeted deworming protocols and improved hygiene measures to control parasite transmission and reduce associated zoonotic risks.
The increasing trend in pet ownership highlights the need for increased awareness and preventive strategies to address the public health challenges posed by parasite infections in companion animals. Future research should focus on identifying emerging trends, assessing the impact of climate change on parasite epidemiology and exploring host–parasite interactions to enhance prevention and control strategies for companion animals. By adopting a One Health approach that collectively considers animal, human and environmental health, we can protect both companion animals and their owners, reducing the impacts of parasite diseases on public health.
These findings highlight the importance of regular coprological analysis for the detection of intestinal parasites, either in veterinary practices or via reference laboratories. Such monitoring is valuable for quality control of established deworming protocols, allowing confirmation of their effectiveness or the need for protocol adjustments.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ministerio de Derechos Sociales, Consumo y Agenda 2030-ANFAAC. https://www.mdsocialesa 2030.gob.es/index.htm. Accessed 4 Dec 2024.
- 2American Animal Hospital Association-American Veterinary Medical Association Preventive Healthcare Guidelines Task Force. Development of new canine and feline preventive healthcare guidelines designed to improve pet health. J Am Anim Hosp Assoc. 2011;47:306–11.10.5326/JAAHA-MS-400721896837 · doi ↗ · pubmed ↗
- 3Mc Grath K, Caldwell P. Diagnostic approach to constipation in children. In: Di Palma J, editor. Constipation–Causes, Diagnosis and Treatment. London: Intech Open; 2012. 10.5772/29180.
- 4ESCCAP UK. http://www.esccapuk.org.uk. Accessed on 2025 Jun 10.