Construction and Expression Analysis of the bin 3xOllas Tool Line
Mengyuan Yi, Hannah Sonnenberg, Vimala Anthonydhason, Ruth H. Palmer

TL;DR
Scientists created a new fruit fly line to track a protein called Bin, which helps study its role in development and reproduction.
Contribution
A new Drosophila knock-in line with a 3×Ollas tag at the biniou locus for precise visualization of the Bin transcription factor.
Findings
The bin 3xOllas allele shows native expression in visceral mesoderm without disrupting Bin function.
Bin is expressed in adult reproductive tissues like ovarian escort cells and testis hub and cyst cells.
Abstract
Precise detection of endogenous protein expression is essential for understanding gene function in vivo . We generated a Drosophila knock-in line inserting a 3×Ollas tag at the C-terminus of the endogenous biniou ( bin ) locus, enabling specific visualization of the Bin transcription factor. The bin 3xOllas allele faithfully recapitulates native expression in the visceral mesoderm and does not disrupt Bin function. Integrating single-cell RNA-sequencing data, we further identified Bin expression in adult reproductive tissues, including ovarian escort cells and testis hub and cyst cells. This allele provides a robust tool for studying Bin function and dynamics.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
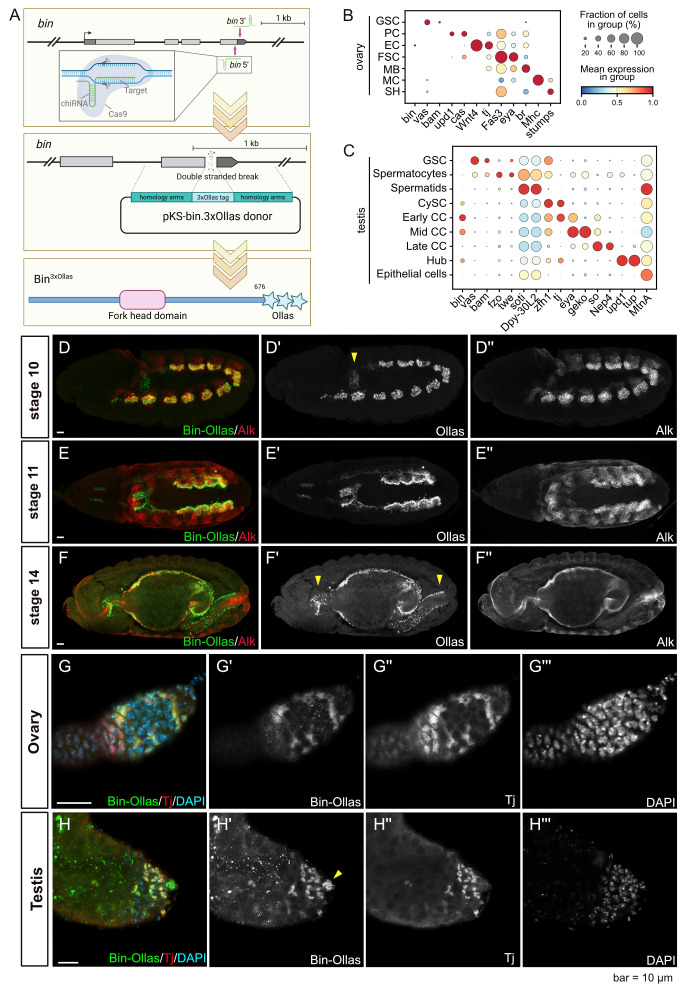 Figure 1
Figure 1|
|
|
|
|
|
Bloomington |
BDSC: 51323 |
|
|
This study |
|
|
|
|
|
|
pU6-BbsI-chiRNA |
Addgene |
45946 |
|
pBlueScript-II-KS(-)-bin.3xOllas donor |
This study |
|
|
|
|
|
|
rat anti-Ollas |
Novus |
MAb L2 |
|
rabbit anti-Alk |
Palmer lab (Lorén et al., 2003) |
Boy |
|
guinea pig anti-Tj |
Gift from D. Godt (Li et al., 2003; Gunawan et al., 2013) |
|
|
Fluorophore coupled secondary antibodies |
Jackson Immunoresearch |
706-606-148 Gp647 111-546-144 Rb488 715-166-151 MCy3 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDevelopmental Biology and Gene Regulation · Genetic and Clinical Aspects of Sex Determination and Chromosomal Abnormalities · Genomics and Chromatin Dynamics
Description
The FoxF family transcription factor biniou (bin) is essential for visceral mesoderm (VM) specification and midgut morphogenesis during Drosophila embryogenesis. Together with Bagpipe (Bap) (Azpiazu and Frasch 1993) and Alk (Lorén et al., 2001), Bin expression is an early marker of the embryonic VM and is employed as a VM marker (Zaffran et al., 2001; Jakobsen et al., 2007). Downstream of Bin lies a broad transcriptional program required for VM development and myogenesis, including bap (which participates in a positive feedback loop), dpp , βTub60D , hand , and many other VM- and muscle-related genes (Lorén et al., 2001; Zaffran et al., 2001; Zaffran and Frasch 2002; Jakobsen et al., 2007; Popichenko et al., 2007; Anllo and DiNardo 2022).
To enable direct detection of endogenous Bin protein, we developed a CRISPR/Cas9-mediated * bin ^3xOllas^ * knock-in allele, which introduces a C-terminal 3xOllas epitope tag ( Figure 1A ). The Ollas tag (14 amino acids) is a synthetic, highly specific epitope with minimal background in Drosophila , thus providing a clean and reliable tool for in vivo immune detection (Park et al., 2008). Importantly, while bin mutants are embryonic lethal (Zaffran et al., 2001), homozygous * bin ^3xOllas ^ * flies were viable and fertile, and no developmental abnormalities were observed, indicating that the tag does not disrupt Bin function. Using this line, we first confirmed that Bin ^3×Ollas ^ faithfully reflects endogenous Bin distribution in the embryonic VM. At stage 10, discrete clusters of Bin * ^3×Ollas^ * –positive cells corresponding to early VM precursors were observed ( Figure 1D ). By stage 11, these clusters extended both anteriorly and posteriorly to form a continuous Bin-positive VM domain, Bin ^3xOllas^ expression appears enriched in founder cells ( Figure 1E ). These founder cells subsequently fuse with neighboring fusion-competent myoblasts to form multinucleate visceral muscle. By stage 14, Bin ^3×Ollas^ -positive cells were observed in the entire muscle layer surrounding the midgut. A strong Bin ^3xOllas^ signal was also consistently observed in the foregut and hindgut mesoderm ( Figure 1F ). These results demonstrate that the * bin ^3×Ollas^ * line faithfully reports endogenous spatial and temporal expression dynamics of bin during embryonic VM development without impacting its transcription factor function, providing a reliable tool for assessing potential expression in additional tissues.
The availability of this line, combined with publicly available single-cell RNA-seq datasets (Li et al., 2022), prompted a re-examination of bin expression beyond embryonic tissues. scRNA-seq profiles revealed unexpected bin expression in adult gonads. In the ovary, bin transcripts were specifically enriched in escort cells (EC), a somatic population surrounding early germline cells ( Figure 1B ). In the testis, bin expression was detected in hub cells and cyst cell lineages, with the strongest signal in early cyst cells ( Figure 1C ). These transcriptomic data suggested previously unrecognized roles for Bin in somatic support cell populations.
Guided by this transcriptomic analysis, we examined Bin ^3xOllas^ expression in adult gonads. In the ovary, Bin ^3xOllas^ localized specifically to EC within the germarium ( Figure 1G ), consistent with both the scRNA-seq results and a recent study showing that Bin maintains EC identity and shapes the GSC-supportive niche by modulating BMP signaling (Tu et al., 2021). Similarly, in the testis, Bin ^3xOllas^ was detected in hub cells and in cyst stem cell (CySC) and early cyst cell populations, with weaker expression in late cyst cells ( Figure 1H ), closely matching the transcriptomic anaylses. Although little is known about Bin function in cyst cells, a recent embryonic study has shown that Bin activity in the VM generates signals such as Slit and FGF to maintain testis precursor niche polarization (Anllo and DiNardo 2022). The presence of Bin protein in adult somatic gonadal cells therefore raises the possibility of direct roles for Bin within the testis itself, a question that remains open for future investigation.
Together, these findings establish the * bin ^3×Ollas^ * line as a reliable and versatile tool for investigating Bin function in diverse developmental contexts and uncovers a previously unappreciated adult gonadal expression of Bin with potential implications for somatic control of germ cell development.
Methods
** Single-cell RNA-seq analysis of Drosophila gonads **
Single-cell RNA-seq data from Drosophila melanogaster testis and ovary were obtained from the Fly Cell Atlas (https://cloud.flycellatlas.org) and analyzed using Scanpy in Python. Cell annotations were harmonized to broader biological categories using published references (Slaidina et al., 2020 for ovary; Witt et al., 2019 for testis). In ovary, cells were grouped into: Germline Stem Cells (GSC), Polar Cells (PC), Escort Cells (EC), Follicle Stem Cells (FSC), Main Body Follicle cells (MB), Muscle Cells (MC), and Sheath Cells (SH). In testis, cells were grouped into: Germline Stem Cells (GSC), spermatocytes, spermatids, Cyst Stem Cells (CySC), Early Cyst Cells (Early CC), Mid Cyst Cells (Mid CC), Late Cyst Cells (Late CC), Hub Cells (Hub), and Epithelial Cells. Marker genes for each population were curated from the literature, and dot-plot visualizations were generated to show scaled expression of lineage-specific markers across gonadal cell types.
** Generation of * bin ^3xOllas^ * Knock-in Fly Allele **
The * bin ^3×Ollas^ * allele was generated following the CRISPR/Cas9 protocol previously used to generate * bap ^HA^ * (Wolfstetter et al., 2025). Briefly, a bin.3×Ollas pBluescript *II KS (-) * (GenScript) donor was constructed containing 782 bp upstream and 776 bp downstream homology arms flanking a sequence encoding three tandem 14 amino acid Ollas tags (SGFANELGPRLMGK–SGFANELGPRLMGK–SGFANELGPRLMGK). The pU6-BbsI-chiRNA gRNA expression vector (Addgene) carrying two CRISPR target sites (5′-TAGGCCGGCTTGCGATCAATGGG-3′ and 5′-ATGCACGCCATCCCAAGTTGAGG-3′) was injected together with the bin.3xOllas donor into * y ^1^ M{vas-Cas9}ZH-2A * embryos (Bloomington 51323) by BestGene Inc.. The * bin ^3×Ollas^ * allele was confirmed by Sanger sequencing (GATC services, Eurofins).
Sample Preparation, Immunofluorescence, and Imaging
All washes were performed with PBS containing 0.3% NP-40 (wash solution). Embryos were collected over a 6–12 h period and dechorionated in 50% bleach (2.5% NaClO) for 3 minutes, then rinsed thoroughly with water. Fixation was carried out in glass vials containing 2 mL of heptane and 2 mL of 4% formaldehyde in PBS, shaken at room temperature (200 rpm) for 40 minutes. After removing the lower formaldehyde phase, 2 mL of methanol was added, and the vial was vigorously shaken for 1 minute to remove the vitelline membrane. Embryos were washed in 100% methanol and stored at –20 °C. Before staining, embryos were rehydrated through 50% methanol/50% wash solution followed by 100% wash solution.
Testes and ovaries were dissected from adult flies anesthetized with CO 2 in cold PBS. Ovarioles were carefully separated using fine needles. Tissues were fixed in 4% formaldehyde for 15 minutes, followed by three washes (3 × 15 minutes) in wash solution.
For immune staining, samples were blocked in 5% normal goat serum (NGS) in wash solution for 30 minutes at room temperature. Primary antibodies were applied overnight at 4 °C. After three washes, samples were incubated with Alexa Fluor-conjugated secondary antibodies (Thermo Fisher) for 2 hours at room temperature in the dark. DAPI was used to label nuclei. After three washes, all samples were mounted in Fluoromount-G (SouthernBiotech). Primary antibodies employed were rat anti-Ollas (MAb L2, Novus) at 1:20, rabbit anti-Alk (Boy) at 1:750 (Lorén et al., 2003), and guinea pig anti-Traffic jam (Tj; gift from D. Godt) at 1:5000 (Li et al., 2003; Gunawan et al., 2013). Secondary antibodies employed were from Jackson Immunoresearch and were employed at 1:500. All samples were imaged using a Zeiss Axio Imager Z2 with LSM 800.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Anllo L Di Nardo S 202246 Visceral mesoderm signaling regulates assembly position and function of the Drosophila testis niche.Dev Cell 5781534-580710091023.e 510.1016/j.devcel.2022.03.00935390292 PMC 9050945 · doi ↗ · pubmed ↗
- 2Azpiazu N Frasch M 199371 tinman and bagpipe: two homeo box genes that determine cell fates in the dorsal mesoderm of Drosophila.Genes Dev 77B 0890-93691325134010.1101/gad.7.7b.13258101173 · doi ↗ · pubmed ↗
- 3Gunawan F Arandjelovic M Godt D 2013529 The Maf factor Traffic jam both enables and inhibits collective cell migration in Drosophila oogenesis.Development 140130950-19912808281710.1242/dev.08989623720044 · doi ↗ · pubmed ↗
- 4Jakobsen JS Braun M Astorga J Gustafson EH Sandmann T Karzynski M Carlsson P Furlong EE 2007101 Temporal Ch IP-on-chip reveals Biniou as a universal regulator of the visceral muscle transcriptional network.Genes Dev 21190890-93692448246010.1101/gad.43760717908931 PMC 1993875 · doi ↗ · pubmed ↗
- 5Li H Janssens J De Waegeneer M Kolluru SS Davie K Gardeux V Saelens W David FPA BrbićM Spanier K Leskovec J Mc Laughlin CN Xie Q Jones RC Brueckner K Shim J Tattikota SG Schnorrer F Rust K Nystul TG Carvalho-Santos Z Ribeiro C Pal S Mahadevaraju S Przytycka TM Allen AM Goodwin SF Berry CW Fuller MT White-Cooper H Matunis EL Di Nardo S Galenza A O'Brien LE Dow JAT FCA Consortium§ Jasper H Oliver B Perrimon N Deplancke B Quake SR Luo L Aerts S Agarwal D Ahmed-Braimah Y Arbeitman M Ariss MM Augsburger J Ayush K Baker CC Banisch T Birker K Bodmer R Bo · doi ↗ · pubmed ↗
- 6Li MA Alls JD Avancini RM Koo K Godt D 20031026 The large Maf factor Traffic Jam controls gonad morphogenesis in Drosophila.Nat Cell Biol 5111465-7392994100010.1038/ncb 105814578908 · doi ↗ · pubmed ↗
- 7Lorén CE Englund C Grabbe C Hallberg B Hunter T Palmer RH 200374 A crucial role for the Anaplastic lymphoma kinase receptor tyrosine kinase in gut development in Drosophila melanogaster.EMBO Rep 481469-221X 78178610.1038/sj.embor.embor 89712855999 PMC 1326337 · doi ↗ · pubmed ↗
- 8Lorén CE Scully A Grabbe C Edeen PT Thomas J Mc Keown M Hunter T Palmer RH 200161 Identification and characterization of D Alk: a novel Drosophila melanogaster RTK which drives ERK activation in vivo.Genes Cells 661356-959753154410.1046/j.1365-2443.2001.00440.x 11442633 PMC 1975818 · doi ↗ · pubmed ↗