Molecular phylogeny and morphology reveal two novel entomopathogenic species of Hypocreales (Polycephalomycetaceae and Cordycipitaceae), from China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
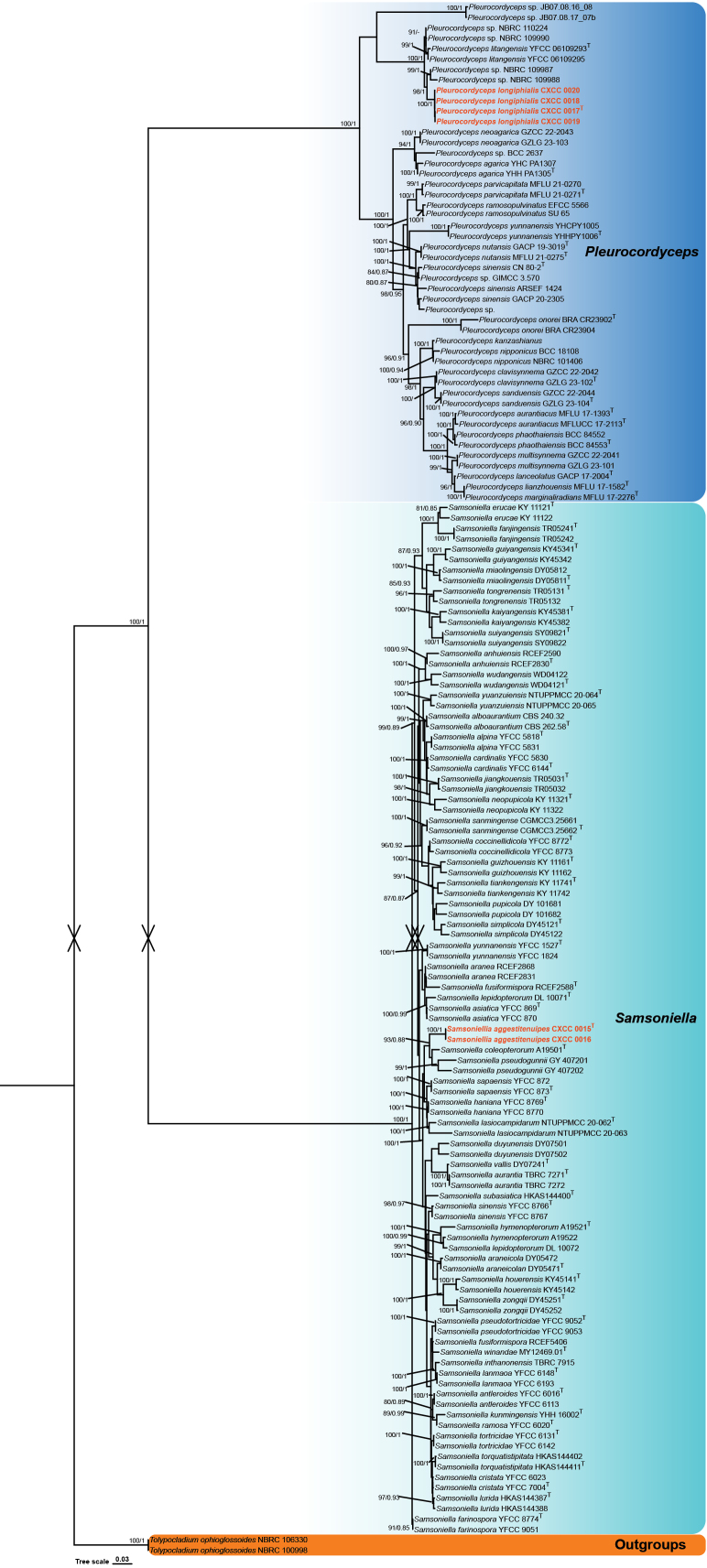 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3| Gene | Primer (forward/reverse) | 5’-Sequence-3’ | References |
| nr SSU | NS1 | GTAGTCATATGCTTGTCTC |
|
| NS4 | CTTCCGTCAATTCCTTTAAG | ||
| nr LSU | LR5 | ATCCTGAGGGAAACTTC | |
| LR0R | GTACCCGCTGAACTTAAGC | ||
|
| ITS5 | GGAAGTAAAAGTCGTAACAAGG |
|
| ITS4 | TCCTCCGCTTATTGATATGC | ||
|
| EF1α-EF | GCTCCYGGHCAYCGTGAYTTYAT | |
| EF1α-ER | ATGACACCRACRGCRACRGTYTG | ||
|
| RPB1-5’F | CAYCCWGGYTTYATCAAGAA | |
| RPB1-5’R | CCNGCDATNTCRTTRTCCATRTA | ||
|
| RPB2-5′F | CCCATRGCTTGTYYRCCCAT | |
| RPB2-5′R | GAYGAYMGWGATCAYTTYGG |
| Current name | Voucher | GenBank accession number | References | |||||
|---|---|---|---|---|---|---|---|---|
|
| nr SSU | nr LSU |
|
|
| |||
|
| YHH PA1305T |
|
| – |
|
|
|
|
|
| YHC PA1307 |
|
| – |
|
|
|
|
|
| MFLUCC 17-2113T |
|
|
|
|
|
|
|
|
| MFLU 17-1393T |
|
|
|
|
|
|
|
|
| GZLG 23-102T |
| – |
|
| – | – |
|
|
| GZCC 22-2042 |
|
|
|
|
|
|
|
|
|
|
|
| – | – | – |
| |
|
| GACP 17-2004T |
|
|
|
|
|
|
|
|
| MFLU 17-1582T |
|
|
|
|
|
|
|
|
| YFCC 06109293T |
|
|
|
|
|
|
|
|
| YFCC 06109295 |
|
|
|
|
|
|
|
|
|
|
|
|
|
| – |
|
|
|
|
|
|
|
|
| – |
|
|
|
|
|
|
|
|
| – |
|
|
|
|
|
|
|
|
| – |
|
|
|
| MFLU 17-2276T |
|
|
|
| – |
|
|
|
| GZLG 23-101 |
|
|
| – | – |
|
|
|
| GZCC 22-2041 |
|
|
|
|
|
|
|
|
| GZLG 23-103 |
| – |
| – | – | – |
|
|
| GZCC 22-2043 |
|
|
|
|
|
|
|
|
| BCC 18108 |
|
|
|
|
|
|
|
|
| NBRC 101406 |
|
|
| – |
| – |
|
|
| GACP 19-3019T |
|
|
|
|
|
|
|
|
| MFLU 21-0275T |
|
|
|
|
|
|
|
|
| BRA CR23902T |
| – | – | – | – | – |
|
|
| BRA CR23904 |
| – | – | – | – | – |
|
|
| MFLU 21-0270 |
|
|
|
|
|
|
|
|
| MFLU 21-0271T |
|
|
|
|
|
|
|
|
| BCC 84553T |
| – |
|
|
| – |
|
|
| BCC 84552 |
| – |
|
|
| – |
|
|
| EFCC 5566 |
| – |
|
|
| – |
|
|
| SU 65 | – | – |
|
|
| – |
|
|
| GZLG 23-104T |
| – |
|
| – |
|
|
|
| GZCC 22-2044 |
|
|
|
|
|
|
|
|
| ARSEF 1424 |
|
|
|
|
|
|
|
|
| CN 80-2T |
|
|
|
|
|
|
|
|
| GACP 20-2305 |
|
|
|
|
|
|
|
| BCC 2637 |
|
|
|
| – |
|
| |
| JB07.08.16_08 |
|
|
|
|
|
|
| |
| JB07.08.17_07b | – |
|
|
|
|
| ||
| NBRC 109987 |
| – |
| – | – | – | Unpublished | |
| NBRC 109988 |
| – |
| – | – | – | Unpublished | |
| NBRC 109990 |
| – |
| – | – | – | Unpublished | |
| NBRC 110224 |
| – |
| – | – | – | Unpublished | |
| GIMCC 3.570 | – |
|
|
|
| – | Unpublished | |
| – |
|
|
| – | – | – | Unpublished | |
|
| YHCPY1005 |
|
|
|
|
|
|
|
|
| YHHPY1006T |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| CBS 262.58T |
| – |
|
| – | – |
|
|
| CBS 240.32 |
| – |
|
|
|
|
|
|
| YFCC 5818T | – |
|
|
|
|
|
|
|
| YFCC 5831 | – |
|
|
|
|
|
|
|
| RCEF2830T | – |
|
|
|
| – |
|
|
| RCEF2590 | – |
|
|
|
| – |
|
|
| YFCC 6113 | – |
|
|
|
|
|
|
|
| YFCC 6016T | – |
|
|
|
|
|
|
|
| RCEF2831 | – |
|
|
|
|
|
|
|
| RCEF2868 | – |
|
|
|
|
|
|
|
| DY05471T |
| – |
|
| – |
|
|
|
| DY05472 |
| – |
|
| – |
|
|
|
| YFCC 869T |
| – | – |
|
|
|
|
|
| YFCC 870 |
| – | – |
|
|
|
|
|
| TBRC 7271T |
| – |
|
|
|
|
|
|
| TBRC 7272 |
| – |
|
| – |
|
|
|
| YFCC 5830 | – |
|
|
|
|
|
|
|
| YFCC 6144T | – |
|
|
|
|
|
|
|
| YFCC 8772T | – |
|
|
|
|
|
|
|
| YFCC 8773 | – |
|
|
|
|
|
|
|
| A19501T |
| – | – |
|
|
|
|
|
| YFCC 6023 |
|
|
|
|
|
|
|
|
| YFCC 7004T |
|
|
|
|
|
|
|
|
| DY07501 |
| – |
|
|
|
|
|
|
| DY07502 |
| – |
|
| – |
|
|
|
| KY 11121T |
| – |
|
| – |
|
|
|
| KY 11122 |
| – |
|
| – |
|
|
|
| TR05241T |
| – |
|
| – |
|
|
|
| TR05242 |
| – |
|
| – |
|
|
|
| YFCC 8774T | – |
|
|
|
|
|
|
|
| YFCC 9051 | – |
|
|
|
|
|
|
|
| RCEF5406 | – |
|
| – |
| – |
|
|
| RCEF2588T | – |
|
| – | – | – |
|
|
| KY45341T |
| – |
|
| – |
|
|
|
| KY45342 |
| – |
|
| – |
|
|
|
| KY 11161T |
| – |
|
| – |
|
|
|
| KY 11162 |
| – |
|
| – |
|
|
|
| YFCC 8769T | – |
|
|
|
|
|
|
|
| YFCC 8770 | – |
|
|
|
|
|
|
|
| KY45141T |
| – |
|
| – |
|
|
|
| KY45142 |
| – |
|
| – |
|
|
|
| A19521 |
| – | – |
|
| – |
|
|
| A19522T |
| – | – |
|
| – |
|
|
| TBRC 7915 |
| – |
|
|
|
|
|
|
| TR05031T |
| – |
|
|
|
|
|
|
| TR05032 |
| – |
|
|
|
|
|
|
| KY45381T |
| – |
|
|
|
|
|
|
| KY45382 |
| – |
|
|
|
|
|
|
| YHH 16002T | – |
|
|
|
|
|
|
|
| YFCC 6193 | – |
|
|
|
|
|
|
|
| YFCC 6148T | – |
|
|
|
|
|
|
|
| NTUPPMCC 20-062T |
| – |
|
|
|
|
|
|
| NTUPPMCC 20-063 |
| – |
|
| – |
|
|
|
| DL 10071T |
| – | – | – |
| – |
|
|
| DL 10072 |
| – | – | – | – | – |
|
|
| HKAS144387T |
|
|
|
|
|
| |
|
| HKAS144388 |
|
|
|
|
|
|
|
|
| DY05811T |
| – |
|
|
|
|
|
|
| DY05812 |
| – |
|
|
|
|
|
|
| KY 11322 |
| – |
|
| – |
|
|
|
| KY 11321T |
| – |
|
| – |
|
|
|
| GY 407202 |
| – |
|
| – |
|
|
|
| GY 407201 |
| – |
|
| – |
|
|
|
| YFCC 9052T | – |
|
|
|
|
|
|
|
| YFCC 9053 | – |
|
|
|
|
|
|
|
| DY 101682 |
| – |
|
| – |
|
|
|
| DY 101681 |
| – |
|
| – |
|
|
|
| YFCC 6020T | – |
|
|
|
|
|
|
|
| CGMCC3.25661 | – |
|
|
|
|
|
|
|
| CGMCC3.25662 T | – |
|
|
|
|
|
|
|
| YFCC 873T |
| – | – |
|
|
|
|
|
| YFCC 872 |
| – | – |
|
|
|
|
|
| DY45121T |
| – |
|
|
| – |
|
|
| DY45122 |
| – |
|
|
| – |
|
|
| YFCC 8766T | – |
|
|
|
|
|
|
|
| YFCC 8767 | – |
|
|
|
|
|
|
|
| HKAS144400T |
|
|
|
|
|
|
|
|
| SY09821T |
| – |
|
|
|
|
|
|
| SY09822 |
| – |
|
|
|
|
|
|
| KY 11741T |
| – |
|
| – |
|
|
|
| KY 11742 |
| – |
|
| – |
|
|
|
| TR05131 T |
| – |
|
|
|
|
|
|
| TR05132 |
| – |
|
|
|
|
|
|
| HKAS144411T |
|
|
|
|
|
|
|
|
| HKAS144402 |
|
|
|
|
|
|
|
|
| YFCC 6142 | – |
|
|
|
|
|
|
|
| YFCC 6131T | – |
|
|
|
|
|
|
|
| DY07241T |
| – |
|
|
|
|
|
|
| MY12469.01T |
| – |
|
|
|
|
|
|
| WD04121T |
| – |
|
| – | – |
|
|
| WD04122 |
| – |
|
| – | – |
|
|
| NTUPPMCC 20-064T |
| – |
| – |
|
|
|
|
| NTUPPMCC 20-065 |
| – |
|
|
|
|
|
|
| YFCC 1527T | – |
|
|
|
|
|
|
|
| YFCC 1824 | – |
|
|
|
|
|
|
|
| DY45251T |
| – |
|
|
|
|
|
|
| DY45252 |
| – |
|
|
|
|
|
|
| NBRC 106330 |
|
|
|
|
|
|
|
|
| NBRC 100998 |
|
|
|
|
|
|
|
| Species | Host/Substrate | Countries found | References |
|---|---|---|---|
|
| China |
| |
|
| Thailand |
| |
|
|
| China |
|
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
|
| China |
|
|
|
|
|
|
|
| Thailand |
| |
|
| China |
| |
|
|
| China |
|
|
| Japan |
| |
|
| China |
| |
|
| Ecuado |
| |
|
|
| Thailand |
|
|
| China | ||
|
| Thailand |
| |
|
| China |
| |
|
|
| Japan |
|
|
|
| China |
|
|
| China | ||
|
|
| China |
|
|
| China |
| Species | Host/Substrate | Countries found | References |
|---|---|---|---|
|
|
|
|
|
|
| Soil, | England |
|
|
| China |
| |
|
| Spider | China |
|
|
| China |
| |
|
| Spider | China |
|
|
| Spider | China |
|
|
| China, Laos, Vietnam |
| |
|
| China, Thailand, Vietnam | ||
|
| China |
| |
|
| China |
| |
|
| Snout beetle ( | China |
|
|
| Saturniida pupae ( | China |
|
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| Spider, | Vietnam |
|
|
| Ant ( | China |
|
|
| Spider | China |
|
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China, Vietnam, Argentina |
| |
|
| China |
| |
|
|
| China |
|
|
| Lepidopterous larvae | Thailand, China |
|
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| Vietnam |
| |
|
|
| UK, Ireland |
|
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
|
| China |
|
|
| China |
| |
|
| China |
| |
|
| Ant | China |
|
|
| China |
| |
|
| China |
| |
|
| Thailand |
| |
|
| China |
| |
|
| China |
| |
|
| China |
| |
|
| China |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEntomopathogenic Microorganisms in Pest Control · Plant Pathogens and Fungal Diseases · Infectious Diseases and Mycology
Introduction
The taxonomic classification of entomopathogenic and mycoparasitic fungi has undergone significant revisions in recent years, driven by advances in molecular phylogenetics (Sung et al. 2007; Mongkolsamrit et al. 2020; Wang et al. 2020; Dong et al. 2022; Kuephadungphan et al. 2022; Tanaka et al. 2023; Tang et al. 2023; Xiao et al. 2023; Chuang et al. 2024). A significant milestone was the establishment of the family Polycephalomycetaceae by Xiao et al. (2023), which was identified as a phylogenetically distinct lineage sister to Ophiocordycipitaceae. This family includes the type genus Polycephalomyces and three other genera, Dingleyomyces, Perennicordyceps, and Pleurocordyceps, which together form a well-supported clade. Most recently, Wang et al. (2024b) described an additional genus, Paradingleyomyces, parasitizing stromata of Ophiocordyceps cf. cochlidiicola in Yunnan Province, China, further expanding the taxonomic breadth of the family. Taxa within Polycephalomycetaceae display notable ecological versatility, infecting a broad spectrum of hosts that include both insects and fungi (Mongkolsamrit et al. 2019; Xiao et al. 2023; Liu et al. 2024b).
Among these genera, Pleurocordyceps was established by Wang et al. (2021) based on integrative morphological and phylogenetic evidence. The type species, Pleurocordyceps sinensis, was originally described as Paecilomyces sinensis by Chen et al. (1984). Xiao et al. (2023) confirmed the delineation of its generic boundaries and familial placement, recognizing 22 accepted species (Index Fungorum, accessed December 2025; https://www.indexfungorum.org/Names/Names.asp). Pleurocordyceps species are obligate parasites of insects or fungi. The sexual morph is characterized by fleshy, stipitate stromata ranging from reddish-brown to yellow, bearing capitate fertile heads with immersed, pyriform to ovoid perithecia. Asci are cylindrical, thick-walled, and feature a distinct apical cap, producing filiform ascospores that disarticulate into uniform cylindrical secondary spores (Wang et al. 2021; Xiao et al. 2023). The asexual morph is hyphomycetous and highly polymorphic, occasionally forming synnemata with conidial masses. Two types of phialides (α and β) produce globose to ellipsoidal α-conidia aggregated in slimy heads, and fusiform β-conidia occurring singly or in chains (Matočec et al. 2014; Wang et al. 2021; Xiao et al. 2023).
A major taxonomic revision by Wang et al. (2021) addressed persistent nomenclatural inconsistencies, such as the misapplication of the name Polycephalomyces formosus due to the erroneous strain ARSEF 1424. Phylogenetic analyses supported the transfer of ten species to Pleurocordyceps: P. agarica, P. aurantiacus, P. lianzhouensis, P. marginaliradians, P. nipponica, P. onorei, P. phaothaiensis, P. ramosopulvinatus, P. sinensis, and P. yunnanensis (Wang et al. 2021). Although Polycephalomyces ramosus represents the earliest described name within the clade and groups phylogenetically with Po. tomentosus and P. sinensis, the latter was designated as the type species due to the more comprehensive morphological and ecological data available (Wang et al. 2021).
Within Cordycipitaceae, the genus Samsoniella was recently erected to accommodate a phylogenetically distinct clade of entomopathogenic fungi, named in honor of Professor Robert A. Samson for his significant contributions to mycological research (Mongkolsamrit et al. 2018). This genus is distinguished by its vivid red-orange stromata in the sexual morph and similarly pigmented, irregularly branched synnemata, and an Isaria-like asexual morph, with oval to fusiform conidia serving as a key diagnostic characteristic (Mongkolsamrit et al. 2018; Dong 2023). Recent taxonomic studies, particularly in China, have rapidly expanded the genus, which currently comprises 56 species, 38 of which have been described in the last 3 years (Index Fungorum, accessed 15 December 2025; https://www.indexfungorum.org/Names/Names.asp). The recognition and circumscription of this genus not only refine the taxonomy of insect-associated fungi but also underscore the continuing expansion of fungal diversity, with S. inthanonensis designated as the type species. Despite these advances, a synthetic overview integrating host range, morphological variation, and geographic distribution for these rapidly diversifying genera is still lacking, and numerous recently collected specimens await study.
Materials and methods
Specimen collection and fungal isolation
Fungal specimens were obtained from Guizhou and Sichuan Provinces, China. Each specimen was photographed, documenting the collection site, GPS coordinates, and altitude. Following surface debris removal, samples were placed in sterile containers and transported under refrigeration to the laboratory. Specimens were subsequently cleaned, assigned accession numbers, and air-dried before processing. Pleurocordyceps was isolated using two methods: (1) tissue isolation, involving surface sterilization of internal sclerotia with 75% ethanol, aseptic dissection into 2–3 mm segments, and transfer to PDA plates; and (2) ascospore isolation, in which stromata were sequentially sterilized with 75% ethanol and 30% H_2_O_2_, rinsed five times with sterile water, dried, and plated as 2–3 mm fragments. The PDA medium (200 g/L potato extract, 20 g/L dextrose, 20 g/L agar) was supplemented with streptomycin (0.1 g/L) and tetracycline (0.05 g/L) after autoclaving to inhibit bacterial growth. For Samsoniella, conidia were directly collected from wild synnemata and inoculated onto PDA. All cultures were incubated at 25 °C, and purified fungal isolates were maintained at 25 °C or stored on PDA slants at 4 °C for preservation.
Voucher specimens (accession nos. CXAC 0015–0020) and corresponding axenic cultures (accession nos. CXCC 0015–0020) are deposited at the College of Agronomy Herbarium (CXAC) and the Chuxiong Fungal Culture Collection (CXCC) at Chuxiong Normal University, China, and are available for taxonomic identification, molecular analysis, and future studies.
Morphological characterization
Ecological data, including host/substrate associations and geographical origin, were documented. Fungal isolates from agar slants were inoculated onto potato dextrose agar (PDA) plates and incubated at 25 °C for several days. Colony morphology was assessed with emphasis on conidial arrangement, phialide structure, and pigment production. For microscopic examination, mycelial fragments from 14-day-old cultures were placed on 5 mm agar blocks, mounted on glass slides, and incubated in moist chambers to promote sporulation. Morphological characteristics of asexual structures, including conidiophores, phialides, and conidia, were observed and measured using an Olympus BX53 compound microscope (Olympus Corporation, Tokyo, Japan).
Extraction of DNA, polymerase chain reaction (PCR), and molecular sequencing
Genomic DNA was extracted from fungal mycelia using a commercial plant DNA isolation kit (FORE GENE, China) and amplified via polymerase chain reaction (PCR) for six genetic regions: ITS, nr SSU, nr LSU, tef1-α, rpb1, and rpb2, with primers listed in Table 1. The PCR was performed in a 25 µL reaction mixture containing 2.5 µL of 10× PCR Buffer (2 mM Mg^2+^; Transgen Biotech), 0.25 µL of Taq DNA Polymerase (Transgen Biotech), 2 µL of dNTPs (2.5 mM), 1 µL of DNA template (~500 ng/μL), 1 µL of each primer (10 μM), and 17.25 µL of deionized water. Amplification was conducted in a BIO-RAD T100™ Thermal Cycler under the thermal cycling conditions described by Dong et al. (2025). The PCR products were electrophoretically verified, purified, and subjected to bidirectional Sanger sequencing at the Beijing Genomics Institute (Shenzhen, China) using the original amplification primers.
Phylogenetic analyses
A six-locus dataset comprising the internal transcribed spacer (ITS), nuclear small and large ribosomal subunits (nr SSU and nr LSU), translation elongation factor 1-α (tef1-α), and the largest and second-largest subunits of RNA polymerase II (rpb1 and rpb2) was assembled for phylogenetic reconstruction. Newly generated sequences were verified through BLAST searches against the GenBank database and combined with reference sequences from related taxa obtained from GenBank (Table 2). Each gene region was independently aligned using MAFFT v7.526 with the L-INS-i strategy (http://mafft.cbrc.jp/alignment/server/, accessed 1 August 2025), followed by manual refinement in BioEdit v7.7.1. The individual alignments were concatenated into a supermatrix using FASconCAT-G v1.06 (Kück and Longo 2014). Phylogenetic conflict among loci was assessed using the partition homogeneity test in PAUP v5.0 with 1000 replicates (p > 0.01). The optimal partitioning scheme and corresponding nucleotide substitution models were selected under the Bayesian Information Criterion (BIC) using PartitionFinder2 v2.0.0 with the greedy algorithm (Lanfear et al. 2016). Maximum likelihood (ML) analysis was performed with IQ-TREE v3.0.1 under the optimal partition scheme with the GTR+G+I model, and branch support was evaluated using 1000 ultrafast bootstrap replicates (Nguyen et al. 2015). Bayesian inference (BI) was conducted in MrBayes v3.2.7. The best-fit substitution model (GTR+G+I) was selected with MrModeltest v2.2 (Nylander 2004). Markov chain Monte Carlo (MCMC) simulations were performed for 5,000,000 generations, sampling every 1,000 generations (Ronquist and Huelsenbeck 2003). Convergence was assessed in Tracer v1.7.2 to ensure effective sample sizes (ESS) exceeded 200, with the first 25% of trees discarded as burn-in (Rambaut et al. 2018). The resulting phylogenetic trees were visualized and annotated in FigTree v1.4.4 (https://tree.bio.ed.ac.uk/software/figtree/), with final layout and editing performed in Adobe Illustrator CS6 following the graphical standards of Xie et al. (2023).
Results
Phylogenetic analyses
The phylogenetic analysis of Pleurocordyceps and Samsoniella species was performed using a data matrix comprising sequences from 149 samples (Table 2). Two strains of Tolypocladium ophioglossoides (NBRC 106330 and NBRC 100998) were used as the outgroups. The final aligned dataset had a total length of 6,034 bp (including gaps), with the following partitions: ITS 768 bp, nr SSU 1,449 bp, nr LSU 913 bp, tef1-α 1,022 bp, rpb1 732 bp, and rpb2 1,150 bp. Both BI and ML analyses generated trees with congruent topologies, in which Pleurocordyceps and Samsoniella formed distinct and well-supported clades (Fig. 1). The overall tree topologies were consistent with those reported in previous studies. The two newly discovered species were placed within well-supported clades in their respective genera: P. longiphialis clustered with P. litangensis and an undescribed Pleurocordyceps sp. (strains NBRC 109987, 109988, 109990, and 110224), while S. aggestitenuipes grouped with S. coleopterorum and S. pseudogunnii. Each of the new species formed a separate, distinct branch within its respective clade, clearly distinguishing them from their closely related species (Fig. 1).
Molecular phylogenetic analyses using the ML and BI based on combined ITS, nr SSU, nr LSU, tef1-α, rpb1, and rpb2 sequence data. Two strains of Tolypocladium ophioglossoides (NBRC 106330 and NBRC 100998) were used as outgroup taxa. Statistical support values (BS ≥ 80% and PP ≥ 0.80) are shown at the nodes for ML bootstrap support (BS) and BI posterior probabilities (PP). Isolates in red type are those analyzed in this study. The scale bar represents the expected number of changes per site.
Morphological features
The morphological characteristics and photomicrographs of the two newly described species, Pleurocordyceps longiphialis (Polycephalomycetaceae) and Samsoniella aggestitenuipes (Cordycipitaceae), are presented in Figs 2, 3, with comprehensive descriptions provided in the Taxonomy section.
Morphology of Pleurocordyceps longiphialis (holotype CXAC 0017; ex-type culture CXCC 0017). A–C. Overview of Pleurocordyceps; D. Colonies on PDA after one month; E. Synnemata on the culture; F, G, I, J, M. α-phialides; H, L. β-phialides and β-conidia; K. β-conidia; N. α-conidia. Scale bars: 20 mm (A); 10 mm (B, C); 10 mm (D); 5 mm (E); 20 µm (F–L); 10 µm (M, N).
Morphological features of Samsoniella aggestitenuipes (holotype CXAC 0015; ex-type culture CXCC 0015). A. Synnemata growing on a lepidopteran pupa; B. Conidia aggregating at the apex of synnemata; C, D, G–N. Phialides with conidia on PDA; E, F. Colonies on PDA after 21 days (E. Obverse; F. Reverse). Scale bars: 10 mm (A, E, F); 3 mm (B); 10 µm (C, D, G–N).
Taxonomy
Two new species are described in this study.
Table 3 lists all hosts, substrates, and geographical locations of Pleurocordyceps species.
Pleurocordyceps
longiphialis
Taxon classificationFungiHypocrealesOphiocordycipitaceae
Q.Y. Dong & C.D. Xu sp. nov.
0F58F123-6EF7-5709-8EF6-982EDFD2034D
860988
Etymology.
The specific epithet longiphialis refers to the characteristically elongated phialides, which distinguish this species from its closely related congeners.
Holotype.
China, • Guizhou Province, Qiandongnan Miao and Dong Autonomous Prefecture, Shibing County, Chengguan Town, on the larva of Scarabaeoidea, 25 March 2025, Quanying Dong (holotype CXAC 0017; ex-holotype living culture, CXCC 0017).
Stromata morph.
Host a Scarabaeoid larva, 47–32.5 × 3–5 mm, swollen, cylindrical, black. Stromata arise from the head and thoracic region of the host, solitary or paired, simple or branched, clavate, yellow to brown, 47–142.5 × 1.5–3.5 mm. In some specimens, the stromatal apex expands into a fist to globose-shaped head, occasionally fissuring into two or three lobes; in others, it divides into two segments, one longer and thicker, the other shorter and narrower. Certain stromata bear a black stipe, 6.5–17 mm long. Sterile.
Culture characters.
Colony on PDA attaining a diameter of 42–44 mm after one month at 25 °C. Colony cottony, fluffy, with high mycelial density, central pale brownish-white umbonate, surrounded by successive concentric rings: an inner dark brown band, a broader light to dark brown zone, a distinct yellow ring, and an off-white periphery with a yellowish tint; reverse dark brown center and creamy-white margin. Hyphae smooth walled, branched, septate, hyaline, 1–2.5 µm wide. Synnemata emerging after two months, clavate or with a mucronate apex, solitary, unbranched, and 2–5 mm long, 0.5–2.5 mm wide, yellowish to yellow, emerging on the middle part of the synnemata or on the top, with conidial masses on the surface (Fig. 2E). Terminal portion of a synnemata covered by a viscous mass. Cultures readily produced phialides and conidia on potato dextrose agar after 14 days at 25 °C. Conidiophores are cylindrical, hyaline, smooth-walled, simple to verticilate form 1–3. Phialides have two types, α-phialides solitary, cylindrical or lanceolate, tapering gradually from base to apex, 9.5–101 × 2–3.5 µm, 2–3.5 µm wide (base), 1–2 µm wide (apex). β-phialides terminal on solitary on hyphae; lageniform or subulate, tapering abruptly from the base to the apex, 7.5–15.5 × 1–2.5 µm, 1–2.5 µm wide (base), 0.5–1.5 µm wide (apex). α-conidia one-celled, hyaline, smooth, subglobose, ovoid to ellipsoidal, 2–3 × 2–3 µm. β-conidia one-celled, hyaline, smooth, fusiform, oblong-elliptical to ellipsoidal, 4.5–8.5 × 1.5–3.5 µm, solitary or aggregated in long chains.
Host.
Scarabaeoidea larva
Known distribution.
Guizhou Province, China.
Additional specimens examined.
China, • Guizhou Province, Qiandongnan Miao and Dong Autonomous Prefecture, Shibing County, Chengguan Town, on the larva of Scarabaeoidea, 25 March 2025, Quanying Dong (paratype CXAC 0018; ex-paratype living culture, CXCC 0018). China, • Guizhou Province, Qiandongnan Miao and Dong Autonomous Prefecture, Shibing County, Baiduo Township, isolated from the larva of Scarabaeoidea, 1 April 2025, Quanying Dong (paratype CXAC 0019, CXAC 0020; ex-paratype living culture, CXCC 0019, CXCC 0020).
Commentary.
The diagnostic morphology of Pleurocordyceps longiphialis conforms to the generic concept of Pleurocordyceps. The species produces dimorphic, often verticillately branched phialides, comprising two types: α-phialides, verticillate with a cylindrical to subulate base tapering into a long neck, and β-phialides, solitary, narrow, lageniform to subulate, and tapering abruptly. Conidia are also dimorphic: α-conidia are globose to subglobose or ellipsoidal, while β-conidia are fusiform and occur solitary or in chains. The species can be distinguished by the following combination of morphological characteristics: α-phialides cylindrical to lanceolate, 9.5–101 × 2–3.5 μm; β-phialides lageniform to subulate, 7.5–15.5 × 1–2.5 μm; α-conidia subglobose, ovoid to ellipsoidal, 2–3 × 2–3 μm; and β-conidia fusiform, oblong-elliptical to ellipsoidal, 4.5–8.5 × 1.5–3.5 μm, occurring solitary or aggregated in long chains. Furthermore, this species was isolated from a Scarabaeoidea larva.
Phylogenetically, Pleurocordyceps longiphialis is strongly supported (BS = 100%, PP = 1) and clusters with P. litangensis (strains YFCC 06109293 and YFCC 06109295) and an undescribed Pleurocordyceps sp. (strains NBRC 109987, 109988, 109990, and 110224). However, it forms a distinct clade separate from these taxa (Fig. 1). Morphologically, P. longiphialis differs from related species in several aspects. P. litangensis, described from Litang County, Sichuan Province, China, possesses aculeate and relatively shorter α-phialides (11.2–12.8 μm vs. 9.5–101 μm in P. longiphialis), longer β-phialides (9.9–27.8 μm vs. 7.5–15.5 μm), larger α-conidia (3.2–6.1 × 1.8–3.9 μm vs. 2–3 × 2–3 μm), and smaller β-conidia (3.5–6.1 × 1.4–2.5 μm vs. 4.5–8.5 × 1.5–3.5 μm). Ecologically, P. litangensis is reported from the host Ophiocordyceps sinensis (Ophiocordycipitaceae), whereas P. longiphialis is associated with a Scarabaeoidea larva (Liu et al. 2024b).
Table 4 lists all hosts, substrates, and geographical locations of Samsoniella.
Samsoniella
aggestitenuipes
Taxon classificationFungiHypocrealesCordycipitaceae
Q.Y. Dong & S.Y. Gao sp. nov.
9A1037E9-FFB2-5DCD-BEDA-6980B52D124D
860989
Etymology.
The epithet aggestitenuipes combines aggesti-, from Latin aggestus, referring to the conidial accumulation, with tenuipes, from Cordyceps tenuipes, reflecting a morphology similar to C. tenuipes but distinguished by synnematal branches that bear massive conidial aggregates.
Holotype.
China, • Sichuan Province, Chengdu City, Qionglai City, Tiantaishan County, isolated from the pupa of Lepidoptera, 21 August 2024, Quanying Dong (holotype CXAC 0015; ex-holotype living culture, CXCC 0015).
Sexual morph.
Undetermined.
Culture characters.
Conidial arrangement Isaria-like. Synnemata two or several, on the pupae of Lepidoptera buried in soil, white, up to 1.8–2.7 cm long. Stipes cylindrical, 0.6 mm wide, producing a mass of conidia at the branches of synnemata, powdery.
Colonies on PDA attaining a diameter of 49–52 mm in 21 days at 25 °C, white to cream-colored, soft cottony aerial mycelium, reverse pale yellow. Hyphae smooth-walled, branched, septate, hyaline, 1–2 µm wide. Synnemata arising from the middle body of pupae were irregularly branched, 1.8–2.7 cm long, 0.3–0.6 mm wide; cylindrical or clavate stipes with powdery white heads. Cultures readily produced phialides and conidia after 1 week on potato dextrose agar at 25 °C showing a granular appearance due to profuse conidiation. Conidiophores cylindrical or clavate, hyaline, smooth-walled, simple to verticilate form 1–3, 5–10.5 × 2–2.5 µm, 1–2 µm wide (base), 2–2.5 µm wide (apex). Phialides from aerial mycelium straight to slightly flexuose, solitary or in whorls of two to four on each branch, cylindrical to flask-shaped, usually with a slightly swollen basal part, 6–28 × 1–2.5 µm, tapering gradually or abruptly from 1–2 µm (base) to 0.5–1.5 µm (apex). Conidia hyaline, subglobose, ovoid, or fusiform, smooth, one-celled, 2–4 ×1.5–2.5 µm, usually in chains.
Other material examined.
China, • Sichuan Province, Chengdu City, Qionglai City, Tiantaishan County, on the pupa of Lepidoptera, 21 August 2024, Quanying Dong (paratype CXAC 0016; ex-paratype living culture CXCC 0016).
Host.
Lepidoptera pupa.
Known distribution.
Sichuan Province, China.
Commentary.
Samsoniella aggestitenuipes displays typical genus-level characteristics such as solitary or whorled phialides and conidia varying from subglobose to ovoid or ellipsoid. It is distinguished by the following unique combination of morphological characteristics: phialides solitary or in whorls of 2–4, cylindrical, 6–28 × 1–2.5 µm; conidia mostly subglobose, ovoid, or fusiform, 2–4 × 1.5–2.5 µm, usually aggregated in chains. Furthermore, the species was isolated from a Lepidoptera pupa. Phylogenetically, S. aggestitenuipes is strongly supported (BS = 93%, PP = 0.88) and clusters with S. coleopterorum and S. pseudogunnii, but it is distinguished from these two species by forming a separate clade within this group (Fig. 1). Morphologically, these two species differ from S. aggestitenuipes in the following ways. Samsoniella coleopterorum, a species described from China, has aculeate and relatively shorter phialides measuring 5.4–9.7 × 1.2–1.8 μm and smaller conidia, 1.7–2.5 × 1.2–1.8 μm vs. 2–4 × 1.5–2.5 µm (Chen et al. 2020). Samsoniella pseudogunnii, a species described from Guiyang, Guizhou Province, and also similar to S. aggestitenuipes in appearance, has relatively shorter phialides (6.8–11 × 2.2–2.4 µm vs. 6–28 × 1–2.5 µm) and relatively smaller conidia (2.8–3.2 × 1.7–2.1 µm vs. 2–4 × 1.5–2.5 µm) (Chen et al. 2021).
Discussion
Employing an integrative taxonomic framework of molecular phylogenetics and morphology, we describe two new entomopathogenic fungi from China: Pleurocordyceps longiphialis and Samsoniella aggestitenuipes. This discovery deepens our understanding of hypocrealean diversity while providing fresh perspectives on host specificity and biogeography. The strong phylogenetic distinction between the species thereby affirms the value of a multilocus methodology for defining species boundaries in these morphologically complex groups.
Host diversity and economic value
Both Pleurocordyceps and Samsoniella exhibit broad host ranges and ecological versatility. Pleurocordyceps infects insects across multiple orders (Coleoptera, Hemiptera, and Lepidoptera) and also parasitizes fungi such as Elaphomyces, Ophiocordyceps, Paraisaria, and Perennicordyceps (Table 3). This dual ecological strategy highlights remarkable adaptability. The discovery of P. longiphialis on a Scarabaeoidea larva extends the genus’ host range to this coleopteran group, revealing a previously unreported niche. Notably, its closest relative, P. litangensis, parasitizes Ophiocordyceps sinensis (Liu et al. 2024b). The observed variation in host use among related Pleurocordyceps species suggests notable ecological plasticity. This pattern could reflect a potential link between host switching from fungi to insects and diversification within the genus, a hypothesis that requires further phylogenetic and genomic investigation.
Samsoniella shows similar host plasticity, infecting Lepidoptera, Coleoptera, Hymenoptera, and Arachnida (Table 4). Although S. aggestitenuipes was isolated from a lepidopteran pupa, a common host among related species, its phylogenetic distinction and morphological traits, such as elongated phialides and unique synnemata, indicate niche specialization. The coexistence of generalist and specialist lineages underscores evolutionary flexibility in host–pathogen interactions within these fungi.
Beyond their ecological significance, the genera Pleurocordyceps and Ophiocordyceps are of considerable economic importance. Pleurocordyceps species produce valuable bioactive metabolites such as antioxidants and antimicrobials from Pl. nipponica and anti-inflammatory agents from P. phaothaiensis, highlighting their potential for pharmaceutical and industrial use (Sangdee et al. 2017; Somsila et al. 2018; Sonyot et al. 2020).
Ophiocordyceps sinensis is a renowned medicinal fungus in traditional Chinese medicine. However, its natural scarcity and high cost limit sustainable utilization. Recent discoveries of novel sterols and immunomodulatory polysaccharides in O. sinensis continue to unveil its complex bioactive foundation (Yao et al. 2024; Qian et al. 2025; Wang et al. 2025). A viable alternative, Samsoniella hepiali, was isolated in 1982 from O. sinensis collected in Yunnan Province. It mirrors the chemical and pharmacological profile of natural O. sinensis, exhibiting analgesic, anti-aging, immunomodulatory, hypoglycemic, nephroprotective, anti-inflammatory, and antidepressant activities. Approved as a health food ingredient in 2001, it is used in the commercial product “Jinshuibao Capsule,” with current annual sales of related products reaching 3 billion CNY (Dai et al. 1989; Wang et al. 2017; Ge et al. 2018; Zhao et al. 2020; Li et al. 2022). Furthermore, S. hepiali serves as a key component in over 260 health products globally, contributing to an estimated yearly market value of CNY 10 billion (Wang et al. 2020). Continued exploration of these genera therefore offers significant opportunities for discovering new bioactive compounds and expanding their economic applications.
Geographic distribution and endemism
Both genera exhibit wide geographic distributions consistent with their broad host ranges. Pleurocordyceps occurs in Asia (China, Japan, and Thailand) and South America (Ecuador) (Table 3), while Samsoniella is found across East Asia (China, Laos, Thailand, and Vietnam), Europe (UK and Ireland), and South America (Argentina) (Table 4). A striking concentration of species discoveries, particularly of Samsoniella, centers in southwestern China, most notably Yunnan Province. This biodiversity hotspot appears to act as a diversification center for these fungi, likely promoted by complex topography, diverse ecosystems, and abundant insect hosts.
The recent discoveries of P. longiphialis in Guizhou and S. aggestitenuipes in Sichuan, both bordering Yunnan, underscore the incomplete documentation of regional fungal diversity, indicating its likely extension beyond currently known ranges. These biogeographic patterns, characterized by widely distributed genera with multiple presumed endemic species, reflect a complex evolutionary history involving both dispersal and isolation. However, sampling bias across regions hinders clear differentiation between true endemism and sampling artifacts.
This study demonstrates the efficacy of integrative taxonomy in revealing cryptic fungal diversity. The newly described P. longiphialis and S. aggestitenuipes offer novel insights into the evolution of Hypocreales, illustrating how host shifts and geographic isolation serve as key drivers of speciation. The extensive host ranges and distinctive biogeographic distributions of genera such as Pleurocordyceps and Samsoniella further reflect their high ecological adaptability. Finally, the biosynthetic potential of these and related taxa warrants further investigation, as they may represent promising sources of novel bioactive compounds for agricultural and pharmaceutical applications.
Supplementary Material
XML Treatment for Pleurocordyceps longiphialis
XML Treatment for Samsoniella aggestitenuipes
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ban S Sakane T Nakagiri A (2015) Three new species of Ophiocordyceps and overview of anamorph types in the genus and the family Ophiocordyceptaceae.Mycological Progress 14: 1–12. 10.1007/s 11557-014-1017-8 · doi ↗
- 2Bischoff JF Rehner SA Humber RA (2006) Metarhizium frigidum sp. nov.: A cryptic species of M. anisopliae and a member of the M. flavoviride complex.Mycologia 98: 737–745. 10.3852/mycologia.98.5.73717256577 · doi ↗ · pubmed ↗
- 3Bu J Wei DP Liu ZH Yang Y Liu ZL Kang JC Peng XC Xie SW Zhang HG He ZJ Huang SK Zhang X Hyde KD Wijayawardene NN Wen TC (2025) Molecular phylogeny and morphology reveal four novel species in Cordycipitaceae in China.Myco Keys 116: 91–124. 10.3897/mycokeys.116.14700640248652 PMC 12004076 · doi ↗ · pubmed ↗
- 4Cao B Phurbu D Ralaiveloarisoa A Liimatainen K Niskanen T Ramírez-Cruz V Bradshaw AJ Dentinger BTM Ramírez-Guillén F Cortés-Pérez A Guzmán-Dávalos L Villalobos-Arámbula AR Castro-Jauregui O Da Silva PS Day R Davoodian N Lebel T Castellano M May TW Toome M Vasey J Hofer K Thangavel R Braithwaite M Braithwaite L Dutta AK Chattopadhyay P Roy N Tanti B Biswas PR Arumugam E Kezo K Kaliyaperumal M Murugadoss R Ji JX Kakishima M Cooper J Nuytinck J Luo ZL Wang WP Zhang X Wen TC Jia AL Fan XL Jiang N Jiao N Zhang Y Manawasinghe IS Li H Xiong YR Zhou T Tan QL Hoshino T Lagaet P De Lange R Verbeken A Tondeleir L Manz · doi ↗
- 5Chaverri P Bischoff JF Evans HC Hodge KT (2005) Regiocrella, a new entomopathogenic genus with a pycnidial anamorph and its phylogenetic placement in the Clavicipitaceae. Mycologia 97: 1225–1237. ttps://10.1080/15572536.2006.1183273216722216 · doi ↗ · pubmed ↗
- 6Chen QT Xiao SR Shi ZY (1984) Paecilomyces sinensis sp. nov. and its connection with Cordyceps sinensis.Mycosystema 3: 24–28.
- 7Chen WH Han YF Liang JD Tian WY Liang ZQ (2020) Morphological and phylogenetic characterisations reveal three new species of Samsoniella (Cordycipitaceae, Hypocreales) from Guizhou, China.Myco Keys 74: 1–15. 10.3897/mycokeys.74.5665533149720 PMC 7588496 · doi ↗ · pubmed ↗
- 8Chen WH Liang JD Ren XX Zhao JH Han YF Liang ZQ (2021) Cryptic diversity of Isaria-like species in Guizhou.Life 11: 1–14. 10.3390/life 11101093 PMC 853993034685462 · doi ↗ · pubmed ↗