Dissection of amino acid acquisition pathways demonstrates that amino acid starvation of Borrelia burgdorferi results in a (p)ppGpp-independent maladaptive response
Arti Kataria, Eric Bohrnsen, Benjamin Schwarz, Dan Drecktrah, D. Scott Samuels, Aaron B. Carmody, Lara M. Myers, Ashley M. Groshong

TL;DR
This study shows that Borrelia burgdorferi, the Lyme disease bacterium, cannot adapt to amino acid starvation using a common bacterial stress response mechanism.
Contribution
The paper reveals that B. burgdorferi lacks a (p)ppGpp-dependent response to amino acid starvation, leading to maladaptive outcomes.
Findings
Peptide transport and the BB0401 transporter are essential for B. burgdorferi viability during infection.
Amino acid starvation does not trigger the canonical stringent response in B. burgdorferi.
The bacterium shows maladaptive phenotypes under severe amino acid stress.
Abstract
Borrelia burgdorferi, the causative agent of Lyme disease, is well known for its unique physiology and enzootic cycle. Building on previous work showing peptide transport is essential for viability, we endeavored to clearly define the impact of peptide starvation on the spirochete and directly compare peptide starvation to targeted free amino acid starvation. Herein, we confirm the ability of a putative glutamate transporter, BB0401, to transport glutamate and aspartate as well as demonstrate its requirement for viability. Using conditional mutants for both peptide transport and BB0401, we characterize these systems throughout the enzootic cycle, confirming their essential role during murine infection and revealing that they are dispensable during prolonged colonization of the tick midgut. We broadly define the metabolic perturbations resulting from these starvation models and show,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
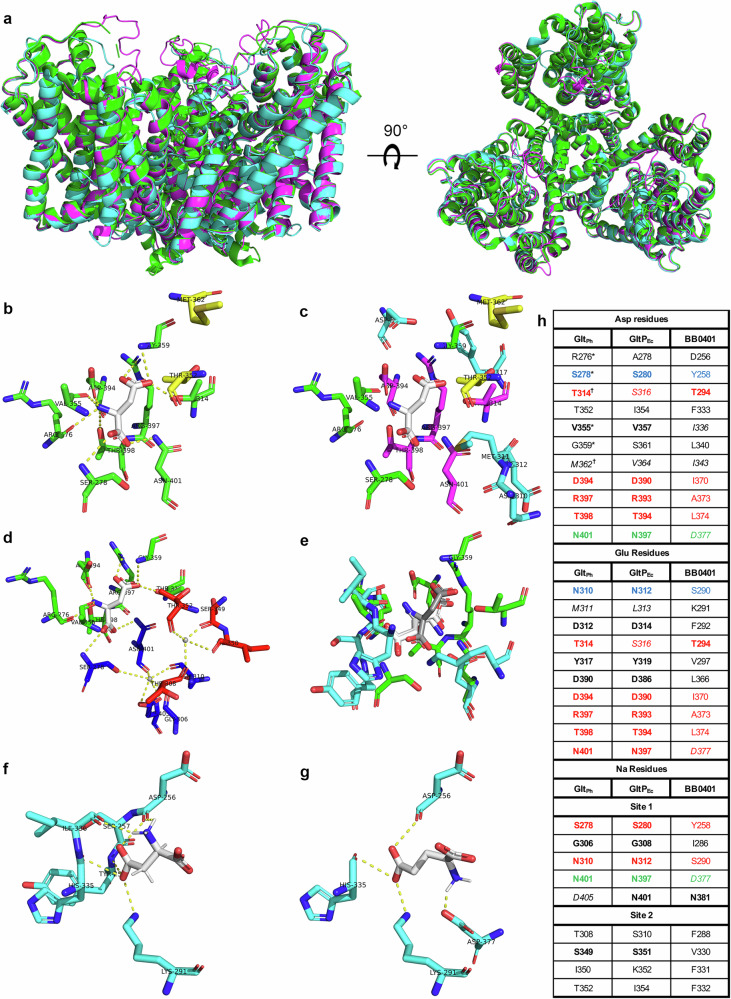 Figure 1
Figure 1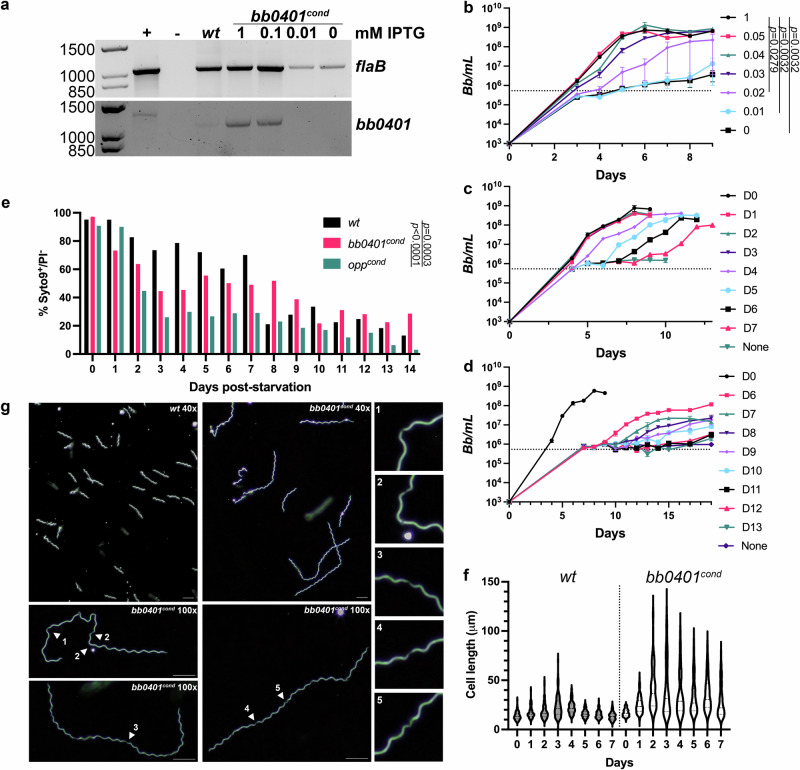 Figure 2
Figure 2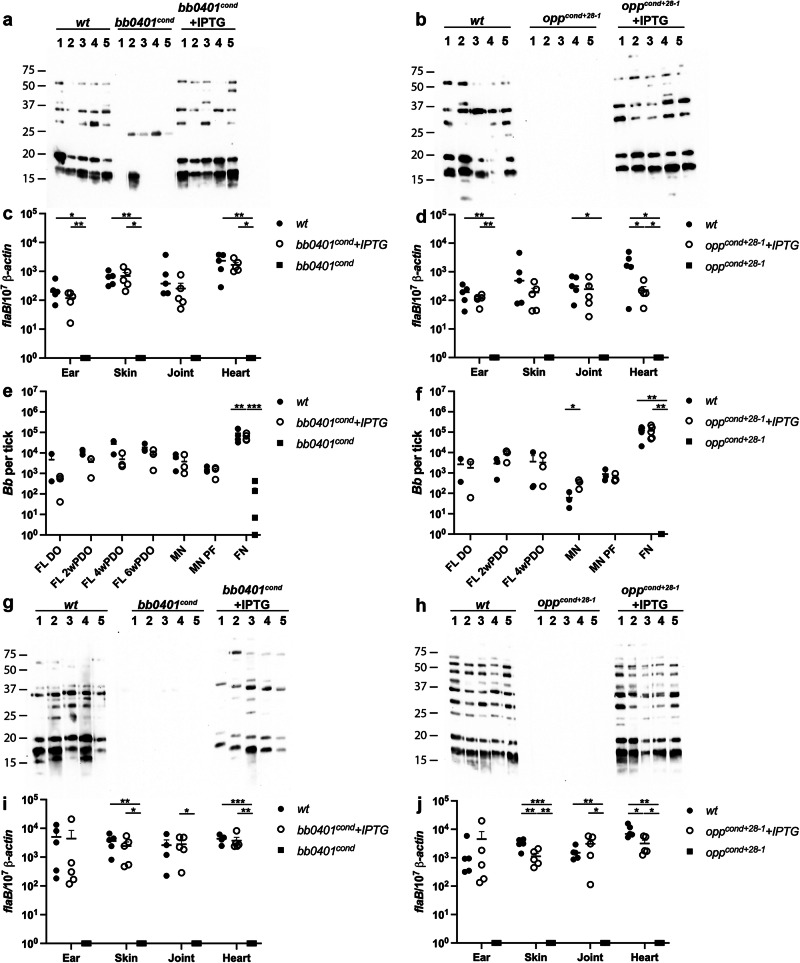 Figure 3
Figure 3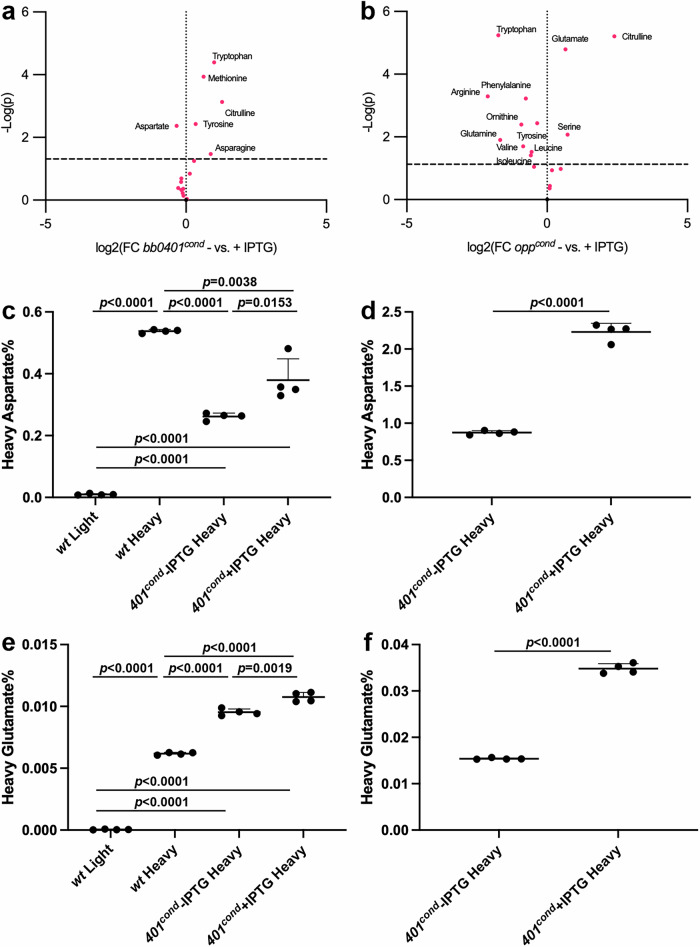 Figure 4
Figure 4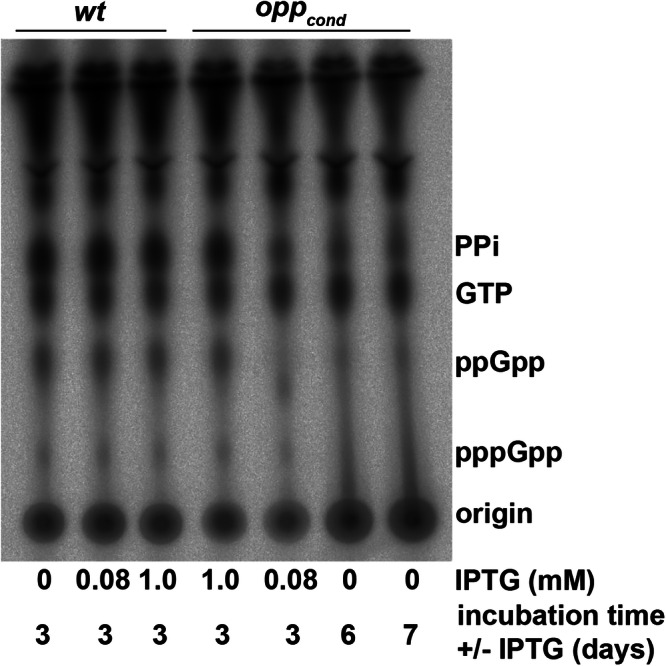 Figure 5
Figure 5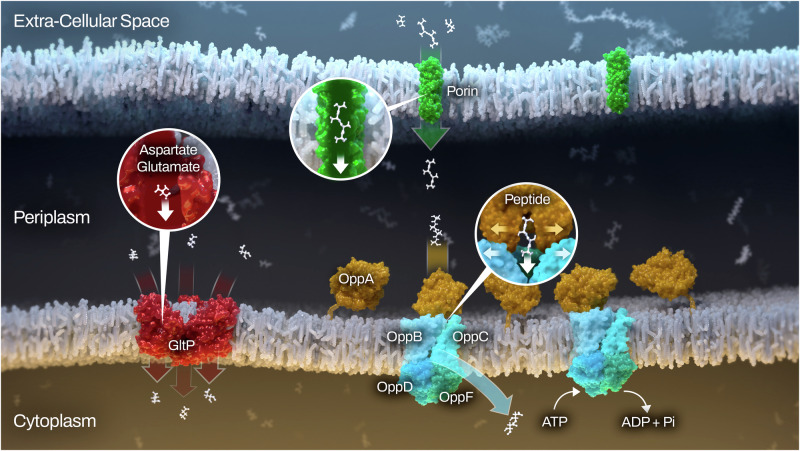 Figure 6
Figure 6- —https://doi.org/10.13039/100006492Division of Intramural Research, National Institute of Allergy and Infectious Diseases (Division of Intramural Research of the NIAID)
- —https://doi.org/10.13039/100000060U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Yersinia bacterium, plague, ectoparasites research · Parasites and Host Interactions
Introduction
Bacterial genomes remain in an evolutionary flux, adjusting to provide physiological support for bacteria to survive within their established niches or to adapt to new ones^1^. Non-fastidious bacteria often retain much of their biosynthetic capabilities, allowing them to survive diverse nutritional environments. Alternatively, fastidious bacteria may relinquish synthetic pathways to allow for pathogenic adaptations or to improve overall cellular economy. The Borrelia burgdorferi genome is well-tailored for its enzootic lifestyle. Several metabolic synthesis pathways have been abandoned given its predictable microenvironments, and genomic real estate has been allocated to its multitude of endogenous plasmids, which contain numerous paralogous surface proteins that define and engage with its narrow vector and broad host ranges^2,3^. The Lyme disease spirochete encodes very few regulatory systems^4–8^, primarily relying on the cyclic di-nucleotide second messenger c-di-guanosine monophosphate (c-di-GMP) controlling tick-phase gene regulation and the RNA polymerase sigma N/RNA polymerase sigma S (RpoN/RpoS; BB0450/BB0771) sigma factor cascade for vertebrate-phase gene regulation. B. burgdorferi has been shown to utilize the alarmone 3’,5’-bis(pyrophosphate)- and guanosine pentaphosphate- ([p]ppGpp) to induced the stringent response under starvation conditions^9–11^. The spirochete encodes a RelA/SpoT homolog (Rel_Bbu_; BB0198) with dual functionality to synthesize and hydrolyze (p)ppGpp^9^. Additionally, B. burgdorferi encodes a DnaK suppressor protein (DksA; BB0168), which is a transcription factor through which (p)ppGpp affects transcription in response to nutrient flux^12,13^. The stringent response in the spirochete has been shown to be critical for survival during the transition from the vegetative cell state in the nutrient-replete molted tick and to the subsequent blood meal^9,13^.
Amino acids are crucial building blocks, needed not only for peptide synthesis but also nitrogen and carbon metabolism, cell-cell communication, and peptidoglycan synthesis; they also serve as intermediates for many metabolic pathways^14^. Canonical bacterial responses to amino acid starvation are hardwired into their transcriptional response via (p)ppGpp and the stringent response^12^. Upon sensing amino acid limitations, bacteria are able to reduce cell growth and/or depend on biosynthetic pathways to support essential amino acid requirements. In the absence of amino acid synthesis pathways, B. burgdorferi depends on transport of peptides or free amino acids to maintain its intracellular free amino acid pools^15–19^. We had previously characterized a conditional mutant of the oligopeptide transport system (Opp) and demonstrated that B. burgdorferi depends on peptide transport for viability^19^. Additionally, we were able to demonstrate that there was only a small window (~48 h) in which starved spirochetes could recover and prolonged starvation resulted in cell elongation, which was unexpected given the limitation of core building blocks. To refine our understanding of amino acid starvation in B. burgdorferi, we sought to compare our peptide starvation model with that of a targeted amino acid deficiency. The partially defined and complex nature of B. burgdorferi cultivation media prevents direct media modifications for something as ubiquitous as amino acids^20^. Therefore, as with peptide transport, we sought to target a specific amino acid transporter.
The Glt transporters are part of a larger family of dicarboxylate/amino acid:cation symporters (DAACS)^21^. These transporters are known to symport sodium ions and/or protons along with their designated substrate(s). The Glt transporters are a subgroup of DAACS that symport L-glutamate, although many have been shown to transport aspartate either preferentially or in tandem. E. coli encodes both a GltP (GltP_Ec_) and GltS (GltS_Ec_), and these transporters demonstrate differences in ion and substrate selectivity^22,23^. GltP_Ec_ facilitates proton-dependent symport of L-glutamate, L-aspartate, and D-glutamate while GltS_Ec_ is sodium-dependent and is primarily an L-glutamate symporter with some tolerance for D-glutamate and L-glutamine^22–24^. The two GltP homologs with solved crystal structures are from archaea, Pyrococcus horikoshii (Glt_Ph_) and Thermococcus kodakarensis (Glt_Tk_)^25–27^. Unlike GltP_Ec_, Glt_Ph_ has been shown to utilize sodium during symport; however, there are conflicting reports about its utilization of protons^26,28,29^; Glt_Tk_ appears to exclusively use sodium ions^27^. Additionally, both transporters have been shown to exclusively transport L- and D-aspartate instead of glutamate^26,27,29^. Other Glt homologs have been characterized with respect to transport selectivity, with Bacillus stearothermophilus GltT (GltT_Bs_) similar to GltP_Ec_ in transport of L-glutamate, L-aspartate, and D-glutamate while Bacillus subtilis GltP (GltP_Bs_) was shown to primarily transport L-glutamate and L-aspartate with minimal transport of D-glutamate, L-glutamine, and L-asparagine^23,30^.
Herein, we characterize the putative GltP (BB0401) in B. burgdorferi^3,31^. Structurally, Swiss-Model^32^ generated BB0401 models using Glt_Ph_ (PDB ID 2nwl)^26^ as a template fit well with other GltP homologs, though binding residues are not highly conserved. We compare a conditional mutant for bb0401 to our peptide starvation model using the conditional Opp transport mutant, allowing us to evaluate the impact of a targeted, limited amino acid starvation to that of a broad, severe amino acid starvation. The phenotypes of the two mutants appear somewhat similar, with the loss of bb0401 affecting growth and morphology as with the Opp system; however, we show that the impacts to cellular processes are distinct. We characterize both the bb0401 and opp conditional mutants in the laboratory model of the enzootic cycle using in vivo IPTG-supplementation, which represents the first use of the IPTG-inducible system for studies of B. burgdorferi in ticks and exposes an unexpected capacity for these mutants to survive within the tick vector given their growth phenotypes in vivo. Metabolomics comparisons define the breadth of the metabolic lesions incurred by these starvation models and heavy labeling experiments confirm that BB0401 is indeed a functional GltP. Finally, we use our peptide starvation model to demonstrate that B. burgdorferi has uncoupled its amino acid starvation response from the (p)ppGpp-driven stringent response, which accounts for the maladaptive phenotype observed in the opp conditional mutant.
Results
BB0401 is the only encoded GltP homolog but demonstrates minimal binding site conservation
Prior annotations of the B. burgdorferi genome originally categorized BB0729 as a GltP homolog, with later analyses uncovering BB0401 as an additional homolog^31^. However, Eggers et al. posited that BB0729 was likely a Na + /cystine symporter, TcyP, based on high homology to the B. subtilis TcyP; this inference was supported by its co-transcription with bb0728, the B. burgdorferi cdr gene encoding CoA-disulfide reductase, as L-cystine is required to generate CoA^33^. The human cystine transporter SLC7A11 is a cationic amino acid transporter (CAT) and was included as an outgroup. To evaluate the relationships between BB0401, BB0729, and other DAACs homologs, we generated a phylogenetic tree with homologs from representative human and bacterial species (Fig. S1). Notably, BB0401 and other spirochete homologs (labeled as GltP) diverged early from other DAACs and are separate from bacterial and archaeal GltPs. Meanwhile, BB0729 and other Borrelia homologs (labeled TcyP) make a distinct clade with other TcyP homologs. While BB0401 is more distantly related to other bacterial DAACs, BB0729 specifically clusters with the TcyP homologs, strengthening the evidence that it may not serve as B. burgdoferi’s GltP homolog.
Glt_Ph_ (PDB ID 2nwl)^26^ has been shown to form a homotrimer (Fig. 1a) consistent with the closely related eukaryotic excitatory amino acid transporters (EAATs) operative in the neuronal glutamate circuit^24–27,34^. Each protomer can transport cargo and trimerization is believed to provide stability within the membrane. The Glt_Ph_ binding pocket for aspartate consists of nine liganding amino acids^24^ (Figs. 1b-d and h, S2b). Of note, four of the nine aspartate-binding residues interact via main-chain carbonyl groups instead of side groups. Binding of sodium ions in Glt_Ph_ (PDB ID 2nwx) have been described for two sites adjacent to the aspartate binding pocket and consists of five amino acids for site 1 and four amino acids for site 2^26^ (Figs. 1d and h, S2d). In a previous study, homology modeling of GltP_Ec_ against the structure of Glt_Ph_ demonstrated a high degree of conservation among the aspartate binding site, though docking studies suggested the glutamate binding site was slightly shifted from the predicted aspartate binding site^24^ (Fig. 1c). Rahman et al. also demonstrated that accessory residues T314 and M362 in Glt_Ph_ influence cargo specificity as mutagenesis of these residues increased glutamate binding^24^. The same study demonstrated that an L313M mutation in GltP_Ec_ ablates glutamate transport, suggesting this substitution in Glt_Ph_ (M311) may contribute to loss of L-glutamate transport^24^.Fig. 1. Residues lining the predicted BB0401 binding pocket are not highly conserved.a Side and top view of Glt_Ph_ (2nwl; green), GltP_Ec_ homology model (magenta), and BB0401 homology model (cyan) alignment. b Residues involved in Asp binding in crystal structure of Glt_Ph_ complexed with Asp (PDB ID 2nwl), Asp is shown in white, accessory residues determined by Rahman et al., in yellow, bonds denoted with yellow dashed lines. c Comparison of Asp binding site shown in the crystal structure of Glt_Ph_ complexed with Asp (PDB ID 2nwl) and corresponding residues as predicted Glu binding site as determined by AutoDock in Rahman et al. Asp is shown in white, residues unique to Asp binding in green, residues unique to Glu binding in cyan, and residues which overlap the two binding sites in magenta. Accessory residues identified by Rahman et al. are shown in yellow. d Residues involved in Na binding in crystal structure of GltPh complexed with Asp and Na (PDB ID 2nwx). Asp is shown in white, Asp liganding residues in green, Na binding site 1in blue, Na binding site 2 in red, and bonds denoted with yellow dashed lines. e Overlay of Asp binding site from crystal structure of Glt_Ph_ complexed with Asp (PDB ID 2nwl) and predicted Asp and Glu binding sites in the BB0401 homology model as determined by SwissDock. Asp from the GltPh crystal structure is shown in grey, Asp and Glu from BB0401 SwissDock both shown in white, residues for Glt_Ph_ binding in green, and putative residues for BB0401 binding as determined by SwissDock in cyan. Individual models of BB0401 SwissDock binding shown for (f) Asp and (g) Glu. Ligands shown in white, bonds denoted as yellow dashed lines. Structures were visualized in PyMol. h) Residue table showing corresponding amino acids based on the Asp liganding residues in Glt_Ph_ crystal structure complexed with Asp (PDB ID 2nwl), the Glu binding residues for GltP_Ec_ as determined by AutoDock in Rahman et al., and the two Na binding site residues as defined in Glt_Ph_ crystal structure complexed with Asp and Na (PDB ID 2nwx). Identical amino acids are in bold, similar amino acids as defined by BLOSUM62 are in italics. Residues that are shared between amino acid binding sites are in red, residues shared with Na binding sites are in blue, and residues shared among all three sites are in green. *Glt_Ph_ aspartate binding residues with main-chain carbonyl group interactions. †Residues identified by Rahman et al. which can increase binding of Glu when mutated in Glt_Ph_.
We generated BB0401 and GltP_Ec_ homology models using SwissModel^32^ based on the crystal structure of Glt_Ph_ (PDB ID 2nwl). The GMQE value for the BB0401 model was 0.57 and QMEANDisCo value was 0.54 ± 0.05. The GMQE value for the GltP_Ec_ model was 0.64 and QMEANDisCo value was 0.64 ± 0.05. Given the low resolution of 2nwl and the low homology between BB0401 and Glt_Ph_, we also modeled BB0401 as a homotrimer using AlphaFold2-Multimer^35^; the AlphaFold model aligned well with the model generated by SwissModel (Fig. S2a, RMSD 1.431 Å). Given the similarity between the two models, we proceeded with the BB0401 model based on the 2nwl crystal structure for simplicity. Using the model of BB0401 we compared overall structure (Fig. 1a) and binding sites (Fig. S2b-d) with that of Glt_Ph_^26^ and the predicted sites for GltP_Ec_^24^. Both models generated by SwissModel model for BB0401 and GltP_Ec_ aligned well with the Glt_Ph_ structure and the RMSD value of Glt_Ph_ and BB0401 was 0.587 Å and Glt_Ph_ and GltP_Ec_ was 0.303 Å. (Fig. 1a). While liganding residues showed consistent positional alignments within the different binding sites (Fig. S2b-d), unlike GltP_Ec_, BB0401 residues lining the different binding pockets showed limited conservation, specifically with respect to amino acid binding sites (Fig. 1h). However, of the six non-conserved residues in the aspartate binding site of BB0401, three are residues that interact via the main-chain carbonyl group (Figs. 1a and h and S2b). Of note, key residue D405 in sodium binding site 1 of Glt_Ph_ correlates with N401 in GltP_Ec_ and N381 in BB0401. A D405N Glt_Ph_ mutant resulted in loss of sodium binding at Site 1^26^, consistent with reports that GltP_Ec_ is not sodium dependent, and suggests that BB0401 may behave similarly. Given the lack of binding site conservation, we used SwissDock to simulate binding of aspartate and glutamate in our model of BB0401. We found that both amino acids were predicted to bind within the same aspartate binding pocket identified in Glt_Ph_ (Fig. 1e). Hydrogen bond predictions in PyMOL suggested only partial conservation with the Glt_Ph_ aspartate liganding residues (BB0401 residues D256/I336 for Asp and D256/D377 for Glu) and showed no conservation with the glutamate binding site predicted for GltP_Ec_^24^ with the exception of D377 which is shared with Glt_Ph_^26^ aspartate binding (Fig. 1f-g). The disparity between binding pocket residues for BB0401 and GltP_Ec_/Glt_Ph_ suggest differences in substrate specificity or transport mechanisms for BB0401.
bb0401 is important for growth in vitro
As previously shown, peptide transport is required for growth as demonstrated via an opp^cond^ mutant^19^. The opp^cond^ mutant was generated by targeting the nucleotide binding domain containing proteins (OppDF; BB0334-5), thus allowing abrogation of peptide transport from all five binding proteins in a single mutant. To characterize the contributions of BB0401 to B. burgdorferi growth and the enzootic cycle, we set out to construct a bb0401 mutant. We generated a Δbb0401 suicide vector (pΔbb0401, Table S1, Fig. S3a) that replaces the bb0401 open reading frame (ORF) with a gentamicin antibiotic selection marker (P_flgB_-aacC1) via homologous recombination. Repeated attempts to transform our wild-type (wt) strain (Table S1) with pΔbb0401 failed, suggesting the gene may be essential for in vitro cultivation. Therefore, we utilized the conditional mutagenesis approach that had previously been used to generate opp^cond 19^. A shuttle vector containing an IPTG-inducible bb0401, constitutive lacI, and P_flgB_-aadA streptomycin antibiotic selection marker (pibb0401, Table S1, Fig. S3b) was generated. pibb0401 was transformed into wt, and a single streptomycin resistant clone was selected for subsequent transformation with pΔbb0401 in the presence of 1 mM IPTG. A single clone (bb0401^cond^) was confirmed to have the bb0401 ORF replaced with the P_flgB_-aacC1 selectable marker and carry the inducible shuttle vector as well as all parental plasmids except cp32-3, cp9, and lp5, which are plasmids known to be dispensable for the enzootic cycle^36^ (Fig. S3c).
We sought to quantify the changes in expression of bb0401 due to IPTG titration within this system. While analysis of protein expression is ideal in such a system, and has indeed been used to confirm expression for previous conditional mutants^37–44^, attempts to express either full length or truncated BB0401 failed. Additionally, generation of serum against a synthesized peptide from BB0401 failed to detect the protein in B. burgdorferi whole cell lysates. Instead, we turned to a semi-quantitative analysis of full-length transcripts using RT-PCR (Fig. 2a). We tested 10-fold reductions in IPTG concentrations to assess the changes in transcription levels of bb0401 compared to the housekeeping gene flaB. We performed first strand cDNA reactions on equivalent amounts of RNA and performed subsequent PCR with our full-length primers to visualize the relative quantities of transcript for our targets. We found a significant reduction of intact flaB transcripts in cells treated with 0.01 mM or no IPTG, suggesting RNA degradation in starved cultures. 1 mM and 0.1 mM IPTG concentrations appeared to have higher levels of transcript compared with the wt strain. Alternatively, 0.01 mM IPTG and absence of IPTG showed to detectable bb0401 transcript as expected under starvation conditions. We noticed a difference in size from our genomic DNA positive control and our RT-PCR samples for bb0401, though the wt and bb0401^cond^ transcripts were of the same size. To confirm this size difference was not due to some modification of the transcript, we sequenced the PCR products and found no changes to the transcript sequence. We believe the size difference is an artifact of the large increase in the loading volumes required to visualize the RT-PCR bands for bb0401 compared to the minimal load of the diluted sample of gDNA for the positive control.Fig. 2. Starvation of bb0401^cond^ significantly impacts growth.a RT-PCR amplification of full transcripts for bb0401 and flaB using equivalent amounts of RNA. “+” represents 20 ng gDNA for flaB and 0.2 ng gDNA for bb0401. Loading volumes for bb0401 RET-PCR samples were 15x greater than flaB to allow for transcript visualization. b Growth curve demonstrating the effects of IPTG titration (0-1 mM) on bb0401^cond^ at an inoculation of 10^3^ spirochetes/ml. Error bars represent ±SEM, n = 3, p values were determined by two-way ANOVA. Graph is representative of three independent growth curves. c, d Recovery growth curves of bb0401^cond^ demonstrating the ability of the starved culture to grow with the addition of 1 mM IPTG on the days designated in the key. Dotted lines denote the limit of detection by darkfield microscopy. Error bars represent ±SEM, n = 3. Graphs are representative of three independent recovery curves. e Percent viability as determined by flow cytometry of wt, bb0401^cond^, and opp^cond^ without IPTG at 37 °C. p values were determined by two-way ANOVA. f Violin plot showing spirochete length for wt and bb0401^cond^ without IPTG as measured by ImageJ. D0 represents normal cell length before starvation. Legnth measurements for wt and bb0401^cond^ (n > 100) are compared by two-way ANOVA, p < 0.0001. g Darkfield microscopy showing wt and bb0401^cond^ without IPTG for 3 days with a 40x or 100x objective. Scale bars represent 10 μm. Numbered arrows show to septation points in cell body and correlate with numbered panels below.
To evaluate the loss of bb0401 expression on growth we performed a growth curve using titrations of inducer for bb0401^cond^ (Fig. 2b) and found that without inducer, bb0401^cond^ demonstrated a significant defect in growth. With respect to IPTG concentrations, there was a more narrow window for intermediate growth (Fig. 2b, 0.02–0.03 mM) compared with opp^cond^ (0.04–0.08 mM)^19^ as well as a lower overall threshold for optimal growth (Fig. 2b, 0.04 mM), as opp^cond^ required at least 0.1 mM IPTG to attain normal growth^19^. We also noted that bb0401^cond^ was able to proliferate during the first three days of cultivation without IPTG ( ~ 100–1000-fold increase overall; Fig. 2b) in contrast to opp^cond^, which demonstrated no growth once inducer was removed from culture^19^.
bb0401cond in vitro phenotype is distinct from oppcond
Abrogation of the Opp system and subsequent peptide starvation in B. burgdorferi resulted in unexpected phenotypes related to growth and morphology^19^. Therefore, we sought to compare the broad amino acid starvation phenotype of the Opp system with the more focused starvation of bb0401^cond^. Previously, we had shown that opp^cond^ was unable to recover growth after approximately 24-48 h of “starvation” (e.g., without inducer)^19^. Surprisingly, bb0401^cond^ was able to recover from lack of inducer over a significant period of starvation (Fig. 2c, d). Initially, we performed a recovery curve where aliquots of bb0401^cond^ with no inducer were spiked with 1 mM IPTG over d 0-7; however, we only noted a small defect in recovery after seven days of starvation (Fig. 2c). Therefore, we performed an additional recovery curve over d 6-13 (Fig. 2d). While we observed similar recovery curves as seen with opp^cond^, where peak densities started to drop with successive recovery curves after a critical period of starvation^19^, bb0401^cond^ was able to starve significantly longer than opp^cond^ without impacting recovery (d 6-7 vs d 1-2, respectively). Overall, growth curves and recovery curves between the two mutants demonstrated similar trends with differences in initial growth before stalling and ability to recovery indicative of either a severe or modest starvation in opp^cond^ and bb0401^cond^, respectively.
Given the drastic difference in growth phenotypes between the two mutants, we sought to quantify cell viability during starvation. For a global assessment of cell viability, we utilized live/dead staining coupled with flow cytometry, comparing opp^cond^ and bb0401^cond^ survival without IPTG to wt grown at 37 °C (Fig. 2e). Gating strategies to identify live/dead populations are shown in Fig. S4a-b. To minimize mutant perturbations during centrifugations, flow was performed on samples in BSK-II media. Syto9^+^/PI^-^ and Syto9^+^/PI^+^ defined our live and dead cells, respectively. Wt was able to maintain ~80% viability up to seven days, approximately 2 d after reaching stationary phase, then dropped to ~20%. Consistent with the published recovery curve^19^, opp^cond^ viability dropped to ~30% by d 3, the point at which normal growth could not be recovered. A low level of viability was maintained by opp^cond^ until d 13-14 where viability is near zero. In contrast, and consistent with recovery curves in Fig. 2c, d, bb0401^cond^ viability drops to ~50% over d 3-8 and ultimately hovers near ~30% by d 14. B. burgdorferi is frequently grown at room temperature to simulate incubation within an unfed tick, a state in which spirochetes undergo little proliferation^9,45^. To determine how reduced pressure to proliferate impacts these mutants, we conducted flow cytometry on cultures incubated at room temperature (Fig. S4c). All cultures maintained >60% viability over a four-week period, the approximate time required for fed larvae to molt.
An unexpected phenotype observed during starvation of opp^cond^ was cell elongation^19^. When grown without IPTG, opp^cond^ was observed to produce a heterogenous population of short and elongated cells, and cell elongation progressed up to about tenfold normal cell length over time. We also observed heterogenous cell length populations when bb0401^cond^ was grown without IPTG (Fig. 2f, g). As with opp^cond^, many of the longest cells were unable to be measured as they create tangled masses in which individual cells cannot be distinguished; however, we were able to clearly show cell elongation for bb0401^cond^ under starved conditions with similar cell length heterogeneity as seen with opp^cond^. Interestingly, there are some clear differences in elongated cell morphology compared with opp^cond^. Elongated opp^cond^ often had extensive regions of cell flattening at the cell center that was shown to be due to loss of flagellar overlap, and cell septation could not be visualized in elongated spirochetes^19^. In contrast, bb0401^cond^ elongated cells maintain planar waveform, and cell septation can be seen at regular intervals (Fig. 2g). Additionally, opp^cond^ maintained motility in elongated cells, with cells that demonstrated cell flattening still motile at cell poles where flagella were intact^19^. Unexpectedly, bb0401^cond^ showed little to no motility after 2 d without inducer, with only occasional twitching despite preservation of planar waveform. These data suggest that while the starvation conditions may be superficially similar, there are distinct phenotypes elicited by targeted free amino acid starvation and peptide starvation.
bb0401 and opp are crucial for murine infection
We sought to determine whether one or both transport systems are critical for the murine model of infection. Previously, we attempted to utilize an IPTG supplementation in vivo model to test infectivity of opp^cond^, which resulted in serological conversion when mice were needle-inoculated and fed IPTG-water but no culture-positive tissues were recovered^19^. We subsequently found that the original opp^cond^ mutant used in the study had spontaneously lost lp28-1 during cultivation, a plasmid that is essential for infection^36^ (Fig. S3c). Therefore, for this infection study we selected a different opp^cond+28-1^ clone what was missing lp21, a plasmid not known to have an impact on the enzootic cycle^36^, but contained lp28-1 (Fig. S3c). We intradermally needle-inoculated mice with 10^4^ spirochetes of strains wt, bb0401^cond^, or opp^cond+28-1^; mutants were inoculated into two different cohorts that were either given water or water supplemented with IPTG. To evaluate infections, we collected sera for western blotting (Fig. 3a, b) and collected tissues for culture (Table 1) and qPCR (Fig. 3c, d) at 2 wk post-inoculation. In contrast to wt, we found that all mice inoculated with opp^cond+28-1^ without IPTG-supplemented water were seronegative (Fig. 3a-b) and culture negative (Table 1) at 2wk post-inoculation. While bb0401^cond^ showed some seroreactivity in mice without IPTG, the antibody response was less robust and likely indicative of a short-term survival before being cleared, consistent with the mutant’s ability to survive in vitro starvation for longer periods of time. Alternatively, all mice that received IPTG-water were culture positive for all tissues from mice infected with either bb0401^cond^ or opp^cond+28-1^ (Table 1). qPCR of tissues at 2 wk post-inoculation (Fig. 3c-d) demonstrated burdens consistent with wt for mice that received IPTG; the only statistically significant difference was an approximate tenfold reduction in heart tissue for opp^cond+28-1^ (Fig. 3d). Alternatively, mice that did not receive IPTG showed no detectable burdens by qPCR. IPTG-supplementation was maintained until 4 wk post-inoculation in a subset of mice used to perform larval acquisition studies and mice maintained positive tissue cultures even at this later timepoint (Table 1). We were able to detect transcripts of bb0401 in hearts infected with wt or bb0401^cond^ where mice received IPTG (Fig. S5c) after 4 wk post-inoculation; transcripts of the mutant were consistent with wt and confirm that IPTG levels in the tissue were sufficient to induce normal levels of transcript. Overall, these data confirm that both bb0401 and opp are essential for murine infectivity. Gilbert et al.^46^ used an in vivo inducible gene system to control OspC expression, a critical gene for early colonization, in mice. This study demonstrated complementation via IPTG-induction of OspC in an OspC mutant at 1 wk post-inoculation, though not all tissues were culture positive^46^. Here, we see the ability of the IPTG-complementation model to maintain infection for up to 4 weeks post-inoculation with viable spirochetes recovered from all tissues. Because IPTG-inducible systems have been shown to accrue mutations in the lac operator when expression is essential for growth^41^, we confirmed that the mutant’s ability to survive in tissues was not due to loss of IPTG sensitivity by passaging cultures from each tissue into media with or without IPTG to test IPTG sensitivity.Fig. 3bb0401 and opp transporters are required for infection and transmission.Western blots of sera against spirochete whole cell lysates for mice needle-inoculated with (a) bb0401^cond^ and (b) opp^cond^. DNA burdens of tissues (n = 5) for mice 2 wk post-inoculation as determined by qPCR for (c) bb0401^cond^ and (d) opp^cond^. Viability plating of fed larvae at drop-off (FL DO, n = 3), fed larvae at timepoints post-drop-off (FL #wPDO, n = 3), post-molt nymphs (MN, n = 3), molted nymphs prior to feeding (MN PF, n = 3), and fed nymphs (FN, n = 5) for (e) bb0401^cond^ and (f) opp^cond^. Western blots of sera against spirochete whole cell lysates for mice nymph-inoculated with (g) bb0401^cond^ and (h) opp^cond^. DNA burdens of tissues (n = 5) for mice 2 wk post-infestation as determined by qPCR for (i) bb0401^cond^ and (j) opp^cond^. Mouse cohorts that received IPTG are represented with a “+IPTG.” All p-values were determined for pairwise comparisons using a two-tailed unpaired t test. p value <0.05, ** <0.01, *** <0.001, **** <0.0001. Exact p values can be found in Supplementary data 3.Table 1. Tissue Culture for in vivo studies2wk post-inoculationwtbb0401*^cond^bb0401^cond^ + IPTG*wtopp*^cond+lp28-1^opp^cond+lp28-1^ + IPTGEar5/50/55/55/50/55/5Inoculation site5/50/55/53/50/55/5Tibiotarsal joint5/50/55/55/50/55/5Bladder5/50/55/54/50/55/5Heart5/50/55/54/50/55/5Total tissues25/250/2525/2521/250/2525/25Total mice5/50/55/55/50/55/54wk post-inoculationwt**bb0401^cond^bb0401^cond^ + IPTGwt**opp^cond+lp28-1^opp^cond+lp28-1^ + IPTGEar4/4ND4/42/2ND3/3Inoculation site4/4ND4/42/2ND3/3Tibiotarsal joint4/4ND4/42/2ND3/3Bladder4/4ND4/42/2ND3/3Heart4/4ND4/42/2ND3/3Total tissues20/20ND20/2010/10ND15/15Total mice4/4ND4/42/2ND3/32wk post-infestationwt**bb0401^cond^bb0401^cond^ + IPTGwt**opp^cond+lp28-1^opp^cond+lp28-1^ + IPTGEar5/50/55/55/50/54/5Inoculation site5/50/55/55/50/50/5Tibiotarsal joint5/50/55/55/50/55/5Bladder5/50/55/55/50/55/5Heart5/50/55/55/50/55/5Total tissues25/250/2525/2525/250/2519/25Total mice5/50/55/55/50/55/5
bb0401 and opp are not required for persistence within the molting tick
To determine how important these transport systems are for tick colonization, we fed naïve Ixodes scapularis larvae on mice infected with wt and mice infected with bb0401^cond^ or opp^cond+28-1^ that were receiving IPTG-supplementation at 2 wk post-inoculation. At drop-off, fed larvae were pooled and evaluated for burdens via semi-solid plating and qPCR (Figs. 3e, f and S5a-b; FL DO). Given that IPTG should be present in the bloodmeal of larvae fed on mutant-infected mice supplemented with IPTG, we expected that the larvae would be successfully colonized. Indeed, both bb0401^cond^ and opp^cond+28-1^ colonized larvae similar to wt as shown by plating and qPCR. To determine whether subsequent digestion of the bloodmeal and tick molting, a process expected to deplete residual IPTG, would impact colonization with the mutant strains, pools of larvae were collected every 2 wk post-drop off until larvae molted (Figs. 3e, f and S5a-b; 2-6wk PDO for bb0401^cond^ and 2-4 wk PDO for opp^cond+28-1^) and after molt (Figs. 3e, f and S5a-b; MN) for evaluation by semi-solid plating and qPCR. Surprisingly, bb0401^cond^ and opp^cond+28-1^ survived as well as wt during the transition from fed larvae to flat nymph. Ten colonies were isolated from each pool of ticks used in plating and confirmed to be IPTG responsive during in vitro culture.
bb0401 and opp are required for survival within the feeding nymph and transmission
Finally, we evaluated the ability of bb0401^cond^ and opp^cond+28-1^ to replicate within the feeding nymph and transmit to naïve mice. We confirmed colonization of molted nymphs 1 d prior to transmission experiments (Figs. 3e, f and S5a-b; MN PF), which was approximately one month after molt. Ten nymphs colonized with wt, bb0401^cond^, or opp^cond+28-1^ were placed on each mouse, and the mutant-infected nymphs were allowed to feed on two different cohorts of mice (with or without IPTG). Pools of nymphs from each mouse were evaluated by semi-solid plating and qPCR for burdens (Figs. 3e, f and S5a-b; FN). Nymphs infected with bb0401^cond^ and fed on mice without IPTG contained DNA burdens equivalent to that of the pre-fed nymphs but failed to show proliferation of spirochetes similar to wt; plating also showed significantly lower viable burdens compared to DNA burdens demonstrating not just the inability to proliferate during feeding, but a significant loss of viable spirochetes. Alternatively, only trace amounts of DNA were detected for opp^cond+28-1^ fed on mice without IPTG and no viable colonies were found by plating. Wt and mutant-infected nymphs fed on mice supplemented with IPTG contained equivalent burdens by plating and qPCR. Again, colonies were recovered from plating, and all isolates were shown to be IPTG responsive confirming there were not spontaneous mutations. To evaluate transmission, we collected sera for western blotting (Fig. 3g, h) and tissues for culture (Table 1) and qPCR (Fig. 3i, j) at 2 wk post-infestation. We found that all mice infested with ticks colonized by bb0401^cond^ or opp^cond+28-1^ without IPTG-supplemented water displayed no seroreactivity, and tissue cultures were all negative at 2 wk post-inoculation. Mice that received IPTG-water had positive tissue cultures for both bb0401^cond^ and opp^cond+28-1^, and tissues at 2 wk post-inoculation demonstrated qPCR burdens consistent with wt for all tissues, with only a small reduction in opp^cond+28-1^ skin and heart. Given the use of IPTG induction during the tick feeding cycle has not been utilized to our knowledge, we sought to detect IPTG in mouse plasma and ticks (Fig. S5d-e). IPTG was only able to be detected above the limit of quantification in mouse plasma and fed nymphs, though samples showed consistent amounts of IPTG within these groups. While we can extrapolate from the mouse plasma IPTG levels that larvae would have obtained some inducer during the bloodmeal, we cannot confirm whether residual IPTG was present in the molting nymph. Overall, these data demonstrate that both bb0401 and opp are essential for replication within the tick and transmission to the host and confirms that the inducible system can be utilized throughout the enzootic cycle.
BB0401 is a GltP
To understand how the loss of these amino acid transporters impacts the spirochete metabolically, we assayed a broad panel of metabolites for +/- IPTG cultures of bb0401^cond^ and opp^cond^ after 48 h of starvation (Figs. 4a, b, S6b-c, Supplementary data 1). As a control for any background metabolic shifts that arise from the use of the inducible system in B. burgdorferi, we included analysis of an inducible GFP (igfp) strain (Fig. S6a, Supplementary data 1). The only metabolite that resulted in a value above the 10% false discovery rate cutoff (FDR; -log(p)=2.7) was citrulline (logFC=0.99) which was higher in the uninduced condition. Intracellular concentrations of 38 and 58 metabolites changed above the 10% FDR threshold for bb0401^cond^ and opp^cond^, respectively (Fig. S6b-c, Supplementary data 1). Of these affected metabolites, 31 showed overlap between the two strains but only 27 of these overlapping metabolites were shifted in the same direction; notably, citrulline was higher in the uninduced sample as seen in the igfp control. Acetyl-CoA was increased during starvation of both mutants with an 8.0-fold increase in bb0401^cond^ and a 9.7-fold increase in opp^cond^. Nucleotides were affected in both mutants in a consistent manner that reflected large scale energy depletion with nucleotide monophosphates increasing and nucleotide triphosphates decreasing under starvation conditions (Fig. S6b-c, Supplementary data 1). In addition to the conserved energy-associated pattern, strain-specific patterns in specialized nucleotide derivatives, including an increase in c-di-GMP (cdGMP), were observed in opp^cond^ but not bb0401^cond^ (Supplementary data 1). Glycolytic and pentose phosphate pathway (PPP) intermediates were largely reduced in both strains with a much stronger phenotype in these pathways for opp^cond^. An exception was observed for fructose 1,6-bisphosphate (FBP) and dihydroxyacetone phosphate (DHAP) which were elevated specifically in bb0401^cond^ starvation. Of the amino acids, only aspartate (logFC = −0.33) was reduced in bb0401^cond^ in the starved condition, while a number of other amino acids were found to be more abundant (e.g. tryptophan). In contrast, opp^cond^ showed reduced levels of multiple amino acids in the starved state (tryptophan, tyrosine, phenylalanine, arginine, glutamine, isoleucine, ornithine, valine, and leucine; logFC between -0.35 and -2.11); only glutamate and serine were more abundant.Fig. 4bb0401 is a GltP.Volcano plots showing amino acid changes in (a) bb0401^cond^, and (b) opp^cond^ when grown without and with 1 mM IPTG. Dotted line represents 10% FDR, n = 4. Tabulated results can be found in Supplementary data 1. Heavy amino acid transport assays for aspartate after (c) 48 h starvation and (d) 7 d starvation and glutamate after (e) 48 h starvation and (f) 7 d starvation of bb0401^cond^. p-values were determined for pairwise comparisons using a two-tailed unpaired t test, n = 4.
While the metabolomics data suggested that BB0401 may transport aspartate, compensation by the peptide transport system, as suggested by the contrasting pattern of tryptophan between the bb0401^cond^ and opp^cond^ mutants, complicates the direct metabolic readout using unlabeled substrates. Therefore, we sought to directly test the transport of glutamate and aspartate by BB0401. Using a heavy isotope labeling assay, we evaluated transport of the two amino acids in bb0401^cond^ under induced and uninduced conditions. To confirm we could detect and differentiate light and heavy amino acids, we tested uptake in wt after incubation with heavy or light amino acids for 2 h and were able to discriminate and detect transport of heavy amino acids (Fig. 4c, e). When we tested transport of these amino acids on bb0401^cond^ with IPTG or 48 h after removal of inducer (Fig. 4c, e), there was a small but statistically significant decrease in the intracellular abundance of these amino acids under the starved condition (1.4- and 1.1-fold decrease for aspartate and glutamate, respectively). Given bb0401^cond^ starvation timepoints were observed to only impact recovery of the bacteria starting at 7 d without inducer (Fig. 2c, d), we repeated the assay by testing transport of amino acids after 7 d of starvation, and we increased the labeling time to 6 h to prolong the time for amino acid flux (Fig. 4d, f). Under these conditions, we saw an increase in levels of heavy labeled amino acids as a percentage of the total amino acid signal, due to the longer exposure time, as well as a more dramatic reduction in the amount of heavy labeled aspartate and glutamate imported in the starved culture (2.5- and 2.3-fold for aspartate and glutamate, respectively). Heavy glutamate was found to be imported at significantly lower percentages of the total glutamate signal than heavy aspartate in all samples, consistent with reports for Glt_Ph_ and Glt_Tk_ preferences for aspartate^26,27,29^.
Amino acid starvation does not stimulate a stringent response
E. coli utilizes the production of (p)ppGpp to initiate a stringent response under amino acid starvation conditions^10,47^. B. burgdorferi can synthesize and degrade (p)ppGpp using the homolog Rel_Bbu_, which is required for the response to nutrient starvation in vitro, for survival in ticks between blood meals, and has been shown to modulate the spirochete’s transcriptome^9,48^. To determine whether this response circuit for amino acid starvation was intact in B. burgdorferi we evaluated the transcriptome using RNAseq. When we first looked at RNA quality from opp^cond^ cultured without inducer for either two or five days, we found that prolonged starvation resulted in significant RNA degradation (Fig. S7). RNA degradation also appeared to be problematic in quantification of bb0401 transcript during starvation, though it is unclear whether this phenotype is unique to amino acid starvation models. Given these results we chose to incubate both mutants without IPTG for two days to standardize the comparison between the two strains and to utilize a timepoint at which both mutants began to show effects of starvation but minimally impacted RNA quality. RNAseq was performed on cultures with and without IPTG for each strain in quadruplicate (Supplementary data 2). Transcriptional changes for bb0401^cond^ +/- IPTG were unremarkable, with bb0401 being the only gene with a |logFC | >1 and a padj < 0.05 (logFC = −3.325). opp^cond^ +/- IPTG demonstrated a larger transcriptional response of 102 genes with a |logFC | >1 and padj < 0.05; 53 genes were up-regulated and 49 were down-regulated under starvation with the greatest fold change for BBD24 (logFC=3.08). The targets of the opp^cond^ mutagenesis, oppD and oppF (logFC = −2.59 and −1.42, respectively), were down-regulated during starvation. Interestingly, some of the substrate binding proteins for the opp system were differentially regulated with oppA1 and oppA3 down-regulated (logFC = -1.55 and -1.22, respectively) and oppA4 was up-regulated (logFC=1.17).
Given that B. burgdorferi encodes three major regulatory systems—Histidine kinase-1/Response regulatory protein-1 (Hk1/Rrp1; BB0420/BB0419) and its product c-di-GMP, RpoN/RpoS, Rel_Bbu_ and its product (p)ppGpp—we compared the opp^cond^ transcriptome to known regulons to determine which regulatory schemes may be amino acid responsive. The only significant overlap was found with the Rel_Bbu_ regulon as defined by Drecktrah et al.^9^. The Rel_Bbu_ regulon was evaluated under several conditions (stationary growth, media starvation, and starvation recovery) with transcriptional variations for each condition; therefore, to simplify the comparison, we tabulated the Rel_Bbu_ regulon as any gene under the three conditions that demonstrated a |logFC | >1. Using these criteria, the Rel_Bbu_ regulon consists of 511 genes with only 42 genes overlapping differential expression in opp^cond^. Of these overlapping genes, only 20 genes were regulated in the same direction due to loss of (p)ppGpp synthesis or amino acid starvation while the remainder were inversely regulated between the two conditions. DksA is also known to work cooperatively with (p)ppGpp or independently to facilitate interactions with RNAP for transcriptional control^12^. Of the 454 genes that define the DksA regulon^13^, only 43 genes overlap with the differentially regulated genes for opp^cond^ and only 3 genes were regulated in the same direction due to loss of DksA or amino acid starvation. Twenty-one genes in this overlap were unique to the DksA regulon and not found in Rel_Bbu_.
The inability to transcriptionally respond to severe amino acid starvation strongly suggests that the (p)ppGpp response in B. burgdorferi is uncoupled from amino acid sensing mechanisms. However, with the small amount of transcriptional overlap, we sought to quantify the production of (p)ppGpp during amino acid starvation using ^32^P-labeling and thin layer chromatography (Fig. 5). We first confirmed that IPTG treatment of wt cultures did not result in changes to (p)ppGpp levels. opp^cond^ was grown under limiting starvation conditions (0.08 mM IPTG, 3 d) and severe starvation conditions (0 mM, 6 d and 7 d)^19^ to evaluate the (p)ppGpp response. Consistent with our transcriptomic data, (p)ppGpp levels did not increase during starvation, instead they appeared to decrease under limiting starvation and could no longer be resolved under severe starvation, suggesting the stringent response was not activated under these conditions.Fig. 5. Abrogation of peptide transport does not result in a (p)ppGpp response.Thin layer chromatography analysis of ^32^P-orthophosphate labeled opp^cond^ cultures with or without IPTG. wt controls with and without IPTG were included to establish baseline (p)ppGpp production.
Discussion
As an extreme auxotroph, B. burgdorferi likely contains many transport systems that are critical for survival^3^. Investigation of these systems are often stymied by limitations in manipulating the complex, partially defined in vitro growth medium and system redundancies encoded in the genome. Herein, we have characterized the contributions of a multipartite oligopeptide transport system and an aspartate/glutamate transporter to meeting the spirochete’s amino acid needs. We previously demonstrated that B. burgdorferi relies on peptide transport for viability^19^, a seemingly expedient approach to amino acid acquisition in the absence of de novo synthesis pathways. However, we hypothesized that, of the handful of free amino acid transporters encoded in the B. burgdorferi genome, we would likely find other essential amino acid acquisition pathways, either to complement amino acid biases related to available peptide composition or for amino acids required for non-proteinogenic roles in the spirochete’s physiology.
There has been a growing list of genes required for B. burgdorferi viability that include the response regulator Rrp2, BamA porin, DedA, telomere resolvase ResT, FtsH protease, RNaseY, Ldh, and BBD18^37–44^. Herein, we find that both Opp and BB0401 are critical transporters for the spirochete both in vitro and in vivo. While GltPs have been shown to provide ancillary amino acid support for bacteria^49–51^, B. burgdorferi is the only bacteria to date that has been shown to require it for viability, similar to the Opp system. Superficially, both mutants display growth and morphology phenotypes that appear similar. It is likely that the subtle differences in IPTG concentrations required for normal growth are reflective of small differences in expression levels needed to meet the spirochetes’ acquisition needs. However, the ability of bb0401^cond^ to survive starvation conditions for significantly longer than opp^cond^ could be indicative of a technical limitation such as RNA or protein stability or the severity of the metabolic lesion such as narrow versus broad amino acid starvations. Interestingly, opp^cond^ spirochetes retain motility during starvation, though the motility is highly dysregulated due to aberrant morphologies^19^. In contrast, bb0401^cond^ quickly became non-motile despite maintenance of the cell’s planar waveform. Indeed, the “elongation” noted for bb0401^cond^ is likely a result of this lack of motility. As seen with the flaB mutant^52^, without the ability to swim in opposing direction during replication, B. burgdorferi cells will often grow in chains where septa can be distinguished at regular intervals as seen with bb0401^cond^. If these chains represent individual cell division events as expected, then our growth curves are likely underestimating bb0401^cond^ growth potential, as long spirochetes were enumerated as single cells. Spirochete morphology and motility is dependent on their endoplasmic flagella and peptidoglycan layer^52^, the latter of which requires glutamate. While we have shown that BB0401 transports both glutamate, one would expect significant perturbation of the cell’s peptidoglycan would result in other morphological anomalies and the lack thereof may suggest a more thorough evaluation of the peptidoglycan composition may be warranted.
Our study of these transporters in vivo required the use of a supplemental IPTG infection model. We were able to show that ad libitum supplementation of IPTG via water was sufficient to complement expression of both bb0401 and opp in the conditional mutants in all tissues collected up to 4 wks during murine infection. We were also able to demonstrate IPTG-induction of bb0401 in vivo resulted in expression levels similar to wt. We were unsure whether these mutants would survive within the tick through the molting phase, as we had no way to supplement IPTG outside of the ticks’ bloodmeal; however, we found that both mutants survived all stages of digestion and molting. While we could not rule out residual IPTG (or other lactose analogs) in the tick due to a high threshold of detection, our room temperature culture experiment (Fig. S4c) strongly suggests that these systems are not critical in the absence of proliferation stimuli. Therefore, the two mutants likely survived within the tick for ~2-2.5 months post-repletion due to bacterial quiescence in the molting nymph and these data shed more insight on the metabolic state of the bacteria in these understudied conditions. Regardless, upon taking the nymphal bloodmeal, these mutants were unable to transition to peak replication phases and resulted in little to no viable bacteria post-feeding. Notably, not only did the mutant spirochetes fail to proliferate during nymphal feeding, but they also failed to survive within the midgut; opp^cond^ tick plating resulted in no viable spirochetes and bb0401^cond^ showed fewer viable spirochetes than the unfed nymph. While there were some viable spirochetes in bb0401^cond^ nymphs, it is likely that the motility defect would have prevented migration out if the midgut and into the skin. Notably, IPTG-supplementation was sufficient to complement both strains in the tick and during transmission and infection. These data demonstrate that IPTG-supplementation during the enzootic cycle is a viable model for evaluating conditional mutants at each stage of the cycle.
We were able to experimentally confirm that BB0401 functions as a GltP. Given the detrimental phenotype of bb0401^cond^ and the homology of BB0729 to TcyP transporters, we believe BB0401 to be the only GltP homolog in B. burgdorferi and propose designating BB0401 as GltP_Bb_; Fig. 6 summarizes our current knowledge of amino acid transport in the spirochete. Of note, GltP_Bb_ preferentially transported aspartate though it appears to retain low affinity for glutamate, which is in contrast to the other characterized homologs which either exclusively transport aspartate (Glt_Ph_ and Glt_Tk_) or display equivalent transport of both glutamate and aspartate (GltP_Ec_) and may contribute to the overall lack of binding residue conservation or the predicted consolidation of the binding site for both amino acids in contrast to GltP_Ec_. Metabolomics also demonstrated the extent of amino acid starvation for opp^cond^, allowing us to extrapolate which amino acids are not being transported via the remaining free amino acid transporters encoded by B. burgdorferi. BB0401 is one of the few transporters that have highly conserved substrates; therefore, these data will be helpful to screen for function related to the remaining transporters. While we saw other metabolic shifts due to these targeted starvations, the metabolic perturbations were surprisingly narrow in both shared and unique changes between the two starvation types. Interestingly, norepinephrine, which is synthesized from tyrosine, was lower in starved opp^cond^ as was tyrosine, suggesting that opp could have some role in transporting amino acids derivatives, though our limited metabolomics approach would likely have missed a number of these targets. Norepinephrine had previously been demonstrated to play a role in up-regulation of OspA^53^, a tick phase surface protein, and has been implicated in virulence of other pathogens^54^. It is unclear which component of BSK-II would provide norepinephrine during in vitro culture, though the medium does contain rabbit serum. It is also possible that we detected a norepinephrine analog that could not be resolved using this metabolomics approach and warrants further studies. Nucleoside mono-, di-, and triphosphates were broadly affected in both strains, with many triphosphates lower under starvation conditions and many mono-/diphosphates higher, indicating a loss of energetic potential. Of note, opp is an ATP-driven transporter where GltP_Bb_ is a symporter, but ATP was lower with starvation of both mutants, suggesting that ablation of NBD-binding in opp^cond^ was not generally responsible for these shifts. Potentially contributing to the drop in nucleoside triphosphates is perturbation of the glycolysis pathway in both strains, which serves as the primary energy generation pathway for B. burgdorferi^3^.Fig. 6. Graphical summary of confirmed amino acid transport mechanisms for *B. burgdorferi.*Non-specific porins (green) in the outer membrane facilitate peptide and amino acid flux into the periplasm and may provide a limitation in peptide size during passive diffusion. The five OppAs (OppA1-5; orange) are lipoproteins tethered to the inner membrane where upon binding peptides in the periplasm they dock to the Opp permease (OppBC; light blue) and transport their cargo to the cytoplasm. The change in permease conformation is facilitated by hydrolysis of ATP by the nucleotide binding domain (OppDF; blue and green). Alternatively, GltP (red) can symport free glutamate or aspartate along with protons and/or sodium ions.
As had been inferred from the significant maladaptive responses in the opp^cond^ mutant^19^, we found that there is no significant stringent response initiated in B. burgdorferi upon amino acid starvation, despite having the machinery to generate and utilize the alarmone (p)ppGpp. While the lack of de novo amino acid synthesis would prevent the spirochete from up-regulating enzyme pathways to supplement amino acids via synthesis, control over aspects such as cell growth is also lost. As a result, we find opp^cond^ suffers from uncontrolled cell elongation under starvation conditions and, though cells may retain viability for a short period of time, these cells lose the ability to recover from starvation quickly. The inability to mitigate amino acid injury by the bacteria is likely unimportant in the context of the enzootic cycle, as the predictable nutritional niches occupied by the spirochete are not specifically deficient in amino acids; therefore, one of the other limited nutrients likely initiates the stringent response. However, in the context of bacterial inhibitors, the failure to respond to amino acid stress presents a novel opportunity to develop targeted and unique approaches to bacterial inhibition. Following this logic, we published a proof-of-concept study that showed the opp system could be targeted by small molecules that resulted in bacterial growth inhibition^55^. The GltP_Bb_ could be targeted in concert with Opp to increase bacterial inhibition. While EAATs are critical to the host^24^ and could complicate off-target effects of inhibiting GltP_Bb_, the lack of conservation among predicted binding pocket residues may help discriminate between the two. The development of more targeted treatment approaches could shift the clinical landscape for Lyme disease treatment, specifically in relation to Lyme-related complications such as persistent infection or post-treatment Lyme disease.
Materials and methods
Sequence alignments and phylogeny
Multiple sequence alignments of proteins were generated using Multiple Sequence Comparison by Log-Expectation (MUSCLE)^56^. The output file was submitted to Phylogenetic Maximum Likelihood (PhyML)^57^ for phylogenetic analysis and the output phylogenetic tree was displayed using the Interactive Tree of Life (iTOL)^58^. The list of proteins and their UniProt IDs can be found in Table S3.
Modeling and docking
BB0401 and GltP_Ec_ was modeled after the Glt_Ph_ crystal structure (PDB ID 2nwl)^26^ using SwissModel^32^ or de novo using AlphaFold2-Multimer^35^. ChimeraX^59^ was used to calculate Root Mean Square Deviation (RMSD) values. SwissDock^60^ was used to dock glutamate and aspartate in the homology model of BB0401. Structures were visualized in either PyMol^61^ or ChimeraX. The crystal structure of Glt_Ph_ bound to sodium ions (PDB ID 2nwx)^26^ was used for the overlay imaging examining conservation of sodium binding sites. The GltP_Ec_ binding residues were identified in a previous study by Rahman et al.^24^.
Bacterial strains and culture conditions
Escherichia coli strains Top10 and Stellar (Table S1) were grown in Luria-Bertani (LB) broth or on LB plates with appropriate antibiotics (ampicillin [Amp; 100 μg/ml], spectinomycin [Spec; 100 μg/ml], or gentamicin [Gent; 5 μg/ml]) at 37 °C. All strains of Borrelia burgdorferi used in this study are derived from B31 5A18 NP1^62^ (Table S1), which has a kanamycin selection cassette inserted in lp25 and is missing lp56 and lp28-4 (Fig. S3). B. burgdorferi strains were cultured in modified Barbour-Stoenner-Kelly II (BSK-II) medium^63^ supplemented with 6% rabbit serum and relevant antibiotics (kanamycin [Kan; 400 μg/ml], streptomycin [Strep; 50 μg/ml], erythromycin [Erm; 0.06 μg/ml] or gentamicin [Gent; 50 μg/ml]) at 37 °C in a CO_2_ incubator (5%) unless otherwise noted. Conditional lethal mutants were grown with the addition of 1 mM isopropyl-β-d-thiogalactoside (IPTG; GoldBio) unless otherwise noted. Conditional lethal mutants without IPTG were maintained in minimal volumes and low densities to prevent spontaneous mutations in the lac operator sequence and the outgrowth of constitutively active transcription of target genes as has been previously seen in conditional lethal Borrelia mutants^41^.
Generation of plasmids and mutant strains
B. burgdorferi B31 chromosomal sequence AE000873.1 was used as reference for all cloning. All plasmids and strains used in this study are referenced in Table S1. All oligonucleotides used in this study are referenced in Table S2. pibb0401 Shuttle Vector. The ORF of bb0401 was amplified using 5’/3’ ibb0401 (1232 bp), which included 15 bp overlap with the 5’ and 3’ restriction sites of the targeted cloning vector. The IPTG-inducible, cp9-based shuttle vector containing a Spec/Strep selection cassette (P_flgB_-aadA), pJSB275^39^, was digested with NdeI/HindIII to release the luciferase ORF and linearize the vector backbone. The bb0401 ORF and pJSB275 vector backbone were joined using InFusion EcoDry (Clonetech), transformed into chemically competent Stellar E. coli (Clonetech), and positive clones were selected with Spec. A positive clone was identified by PCR using 5’/3’ pJSB275 seq and sequence was confirmed by Sanger sequencing. The resulting plasmid was designated pibb0401 (pEcAG286). pΔbb0401 Suicide Vector. The regions upstream (F1; 1073 bp) and downstream (F2; 1073 bp) of bb401 were amplified using 5’/3’ bb0401 null F1 and 5’/3’ bb0401 null F2, respectively. The P_flgB_-aacC1 Gent cassette (966 bp) was amplified from pBRV2 using 5’/3’ bb0401 null gent. Primers for all three fragments included 15 bp overlaps with adjacent fragments and were assembled into BamHI-digested pUC19 using InFusion EcoDry, transformed into chemically competent Stellar E. coli, and positive clones were selected with Gent. A positive clone was identified by PCR using pless Gent F/R and sequence was confirmed by Sanger sequencing. The resulting plasmid was designated pΔbb0401 (pEcAG259) ibb0401 Mutant. Competent B. burgdorferi B31 5A18 NP1^62^ was electroporated with pibb0401 as previously described^64^ and transformants were selected in BSK-II with Kan and Strep. Clones were screened for the presence of aadA with pless Strep F/R and DNA from transformants were subsequently transformed into E. coli Top10 to confirm recovery of shuttle vector. Selected clones were screened for endogenous plasmid content using a modification of the plasmid multiplex PCR described in Bunikis et al.^65,66^. A single clone was selected for subsequent cloning and designated ibb0401. bb0401^cond^ Mutant. The conditional bb0401 mutant was generated by electroporating competent ibb0401 with linearized pΔbb0401 and transformants were selected in BSK-II with Kan, Strep, Gent, and 1 mM IPTG. Single clones were screened for the presence of aacC1 with pless Gent F/R. Selected clones were screened for endogenous plasmid content using a modification of the plasmid multiplex PCR^65,66^. A clone with all parental plasmids except cp32-3, cp9, and lp5, plasmids known to be dispensable for the enzootic cycle^36^, was selected for characterization and designated bb0401^cond^ (Fig. S3). pigfp Shuttle Vector. The ORF of gfp was amplified using 5’/3’ iGFP (750 bp), which included 15 bp overlap with the 5’ and 3’ restriction sites of the targeted cloning vector and was cloned as described above for pibb0401. A positive clone was identified by PCR using 5’/3’ pJSB275 seq and sequence was confirmed by Sanger sequencing. The resulting plasmid was designated pibb0401 (pEcAG304). igfp Strain. Competent B. burgdorferi B31 5A4^36^ was electroporated with pigfp as previously described^64^ and transformants were selected in BSK-II with Strep. Clones were screened for the presence of aadA with pless Strep F/R and DNA from transformants were subsequently transformed into E. coli Top10 to confirm recovery of shuttle vector. qPCR standards. 5’/3’ primers for flaB, bb0401, and mouse β-actin were used to amplify target coding regions for each gene and fragments were cloned into pCR2.1 using the TOPO Cloning Kit (Thermo). Individual clones were confirmed via Sanger sequencing.
Growth kinetics
bb0401^cond^ growth was evaluated similar to that of opp^cond 19^. In brief, cultures were grown in BSK-II medium with 1 mM IPTG and appropriate antibiotics to mid-logarithmic growth. Cultures were washed with PBS to remove IPTG and resuspended in BSK-II at the density of 1 × 10^6^ or 1 × 10^3^ cells/ml respectively with varying IPTG concentrations (0–1 mM). In case of recovery analysis, cultures were washed as described above and resuspended in BSK-II media without IPTG and incubated at 37^o^C; each day, 1 mM IPTG was added to aliquots of cultures to determine their ability to recover from lack of induction. All growth experiments were performed in triplicate and cell density was enumerated daily by dark-field microscopy. Growth curves were plotted using Prism software (v9.5.1; GraphPad Software, Inc.) and are representative of three individual experiments.
Microscopy and measurement of spirochete length
Wt and bb0401^cond^ cell lengths were evaluated similar to that of opp^cond 19^. Strains were grown in BSK-II medium (bb0401^cond^ with 1 mM IPTG) with appropriate antibiotics to mid-logarithmic growth. Cultures were washed with PBS and resuspended in BSK-II at the density of 1 × 10^6^ spirochetes/ml in the absence of IPTG for wt and bb0401^cond^ or with 1 mM IPTG for bb0401^cond^. Images were acquired daily using a 40x objective and a minimum of 100 spirochete lengths were measured for each sample using ImageJ 1.54 g. Images to inspect individual cells were collected using a 100x objective and processed using ImageJ 1.54 g. These data were graphed as whisker plots using Prism software. Images were collected with a Zeiss Axiocam 208 color camera (Zeiss).
RT-PCR
For quantification of bb0401 transcript by qRT-PCR in vitro, wt and bb0401^cond^ strains were grown in BSK-II medium with 1 mM IPTG for the mutant strain and appropriate antibiotics to mid-logarithmic growth. Cultures were washed with PBS to remove IPTG and resuspended in BSK-II at the density of 1×10^6^ spirochetes/ml in triplicate and growth at 37 °C for 2 d. bb0401^cond^ was growth with 1 mM, 0.1 mM, 0.01 mM or without IPTG. Samples were collected and RNA was purified using the Zymo Direct-zol RNA miniprep kit (Zymo Research) according to manufacturer’s protocols. Contaminating DNA was removed using TURBO DNA-free Kit (Thermo). Equivalent amounts of RNA were converted to cDNA using the SuperScript First-Strand Synthesis System (Thermo) and ‘3 primers for the flaB and bb0401 ORFs (Table S2). Equivalent amounts of cDNA were subjected to PCR using 5’/3’ primers for the flab and bb0401 ORFs (Table S2) and Apex Red Master Mix (Genesee Scientific).
Flow cytometry
wt, opp^cond^, and bb0401^cond^ were grown in BSK-II medium with 1 mM IPTG and appropriate antibiotics to mid-logarithmic growth. Cultures were washed with PBS to remove IPTG and resuspended in BSK-II at the density of 1 × 10^4^ spirochetes/ml for wt and bb0401^cond^ and 1 × 10^6^ spirochetes/ml for opp^cond^ and incubated at 37 °C or 1 × 10^7^ spirochetes/ml for all strains at room temperature incubation. The cell viability was tested using BDFacs Symphony cytometer using standard live/dead stains, Syto 9 (1 μM) (Thermo Fisher Scientific) propidium iodide (5 μM) (Thermo Fisher Scientific). Samples were stained and analyzed directly in BSK-II using the gating strategy described in Fig. S4a-b. Particulate matter was identified and excluded using forward- and side-scatter then bacterial cells were gated using Syto9 (FITC channel) and propidium iodide (TxRed-mCh channel).
Ethics statement
All animal work was conducted according to the guidelines of the National Institutes of Health, Public Health Service Policy on Humane Care and Use of Laboratory Animals, and the United States Institute of Laboratory Animal Resources and National Research Council, Guide for the Care and Use of Laboratory Animals^67^. All protocols were approved by the Rocky Mountain Laboratories, NIAID, NIH Animal Care and Use Committee (2021-041E). The Rocky Mountain Laboratories are accredited by the International Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC). We have complied with all relevant ethical regulations for animal use.
Infection studies
All infections were performed on female C3H/HeJ mice aged five to eight weeks (Jackson Laboratories). Cohorts of 5 mice were utilized for 2wk infection timepoints for both needle- and nymph-inoculation based on a Fisher’s Exact Test of determining infection (99%) or no-infection (1%) probability with an alpha of 0.01 and a power of 90. Our 4wk mouse infection cohorts were sized to generate enough ticks for subsequent analysis. No exclusion criteria, randomization, or blinding strategies were used during the experiment. B. burgdorferi strains were used to needle-inoculate cohorts of mice intradermally or infected nymphs (n = 10) were placed on mice as described below. For opp^cond^ and bb0401^cond^, cohorts of mice were either given IPTG water (2% sucrose, 80 mM IPTG) or water alone starting a week before infection, and this was maintained throughout the duration of the study. Tissues (ear, inoculation site skin, tibiotarsal joint, bladder, and heart) were collected for qPCR and cultured in BSK-II supplemented with BAM (Borrelia antibiotic mixture; 0.05 mg/ml sulfamethoxazole, 0.02 mg/ml phosphomycin, 0.05 mg/ml rifampicin, 0.01 mg/ml trimethoprim, and 2.5 μg/ml amphotericin B) and 1 mM IPTG (for mutants only). DNA was isolated from qPCR tissue samples using DNeasy Blood and Tissue Kit (Qiagen) modified by pre-digestion with collagenase IV (Sigma) and proteinase K (Sigma). qPCR assays were conducted using TaqMan Fast Advanced Master Mix (Thermo) on a QuantStudio 5 (Thermo). At two- and four-weeks post-infection mouse sera was collected and blotted at 1:1000 against B. burgdorferi whole cell lysates on nitrocellulose, followed by goat anti-mouse HRP at 1:15000 (Southern Biotech). Blots were developed with SuperSignal West Pico PLUS (Thermo) and imaged on iBright 1500 (Invitrogen).
qRT-PCR
For quantification of bb0401 transcript by qRT-PCR in vivo, infected tissues were disrupted via the Qiagen TissueLyser LT and RNA was purified using the Zymo Direct-zol RNA miniprep kit (Zymo Research) according to manufacturer’s protocols. Contaminating DNA was removed using TURBO DNA-free Kit (Thermo). Plus and minus RT cDNA libraries were made using SuperScript IV VILO Master Mix (Thermo) and qPCR was performed using TaqMan Fast Advanced Master Mix (Thermo) and primers/probes listed in Table S2 on a QuantStudio 5 (Thermo) with plasmids containing the target ORF serving as quantification standards.
Tick studies
Pathogen-free Ixodes scapularis larvae (Oklahoma State University) were allowed to naturally acquire a bloodmeal from infected mice at two-weeks post-infection via whole body infestation. Fed larvae were allowed to molt into flat nymphs and were subsequently fed on naïve mice via capsule feeding^68^ to measure transmission. Pooled ticks at all stages were evaluated for spirochete burdens via semi-solid plating^64^ and qPCR^69^ as previously described. Semi-solid plating media for all strains included BAM and conditional mutants were also grown with 1 mM IPTG. qPCR assays were conducted using TaqMan Fast Advanced Master Mix on a QuantStudio 5.
RNAseq
To test RNA quality, opp^cond^, the most impaired mutant, was grown BSK-II medium with 1 mM IPTG and appropriate antibiotics to mid-logarithmic growth. Cultures were washed with PBS to remove IPTG and resuspended in BSK-II at the density of 1 × 10^7^ spirochetes/ml with or without 1 mM IPTG and grown at 37 °C for 2 d or 5 d in quadruplicate. Samples were analyzed on the BioRad Experion according to manufacturer’s protocols. For RNAseq analysis opp^cond^ and bb0401^cond^ were grown as described but incubated for only 2 d in quadruplicate. Cell pellets were collected, and samples were flash frozen in liquid nitrogen and sent to SeqCenter for RNAseq analysis according to their standard prokaryotic protocols and genes on plasmids that were not present in all samples were excluded from comparison. The full data table is available as Supplementary data 2.
Metabolomics
opp^cond^ and bb0401^cond^ were grown in BSK-II medium with 1 mM IPTG and appropriate antibiotics to mid-logarithmic growth. Cultures were washed with PBS to remove IPTG and resuspended in BSK-II at the density of 1 × 10^7^ spirochetes/ml with or without 1 mM IPTG and grown at 37 °C for 48 h in quadruplicate. Sample were then washed 0.9% sodium chloride solution (Teknova) and treated with LCMS grade methanol (Fisher Scientific), incubated on ice, and an equal volume of LCMS grade water (Fisher Scientific) was added. Samples were agitated to ensure layer mixing for 30 m under refrigeration and centrifuged at 16,000 × g for 20 min. 400 μl of the top (aqueous) layer was collected for LCMS analysis of central polar metabolites.
Aqueous metabolites were analyzed via liquid chromatography tandem mass spectrometry (LC-MS/MS) using a combination of two analytical methods with opposing ionization polarities^70,71^. All methods used a LD40 XR UHPLC (Shimadzu Co.) system for separation and a 6500+ QTrap mass spectrometer (AB Sciex Pte. Ltd.) for detection and quantification. Negative mode samples were eluted from a Waters™ Atlantis T3 column (100 Å, 3 μm, 3 mm × 100 mm) using a binary gradient from 5 mM tributylamine, 5 mM acetic acid in 2% isopropanol, 5% methanol, 93% water (v/v) to 100% isopropanol over 5 m. Two distinct multiple reaction monitoring (MRM) pairs in negative mode were used for each metabolite. Positive mode samples were eluted from a Phenomenex Kinetex F5 column (100 Å, 2.6 μm, 100 × 2.1 mm) with a gradient from 0.1% formic acid in water to 0.1% formic acid in acetonitrile over 5 m. Tributylamine and all synthetic molecular references were purchased from Millipore Sigma. LCMS grade water, methanol, isopropanol and acetic acid were purchased through Fisher Scientific.
All signals were integrated using SciexOS 3.1 (AB Sciex Pte. Ltd.). Signals were confirmed visually. Signals with greater than 50% missing values were discarded and remaining missing values were replaced with the lowest registered signal value. Filtered datasets of the negative mode aqueous metabolites total sum normalized after initial filtering. The positive mode aqueous metabolomics dataset was scaled and combined with the negative mode aqueous metabolite dataset using common signals for tyrosine and phenylalanine to generate scalars between the datasets. A Benjamini-Hochberg method for correction for multiple comparisons was imposed to establish false discovery rate (FDR) cut-offs for significance.
Amino acid uptake assay
opp^cond^ and bb0401^cond^ were grown in BSK-II medium with 1 mM IPTG and appropriate antibiotics to mid-logarithmic growth. Cultures were washed with PBS to remove IPTG and resuspended in BSK-II at the density of 1 × 10^7^ spirochetes/ml with or without 1 mM IPTG and grown at 37 °C for 48 h or 7 d in quadruplicate. An equal density of wt cells were prepared for light and heavy controls. To compete with unlabeled amino acids, we added 1:1 heavy amino acids as defined by the CMRL content. Therefore, when cultures were ready, 30 mg/l of heavy labelled aspartic acid and 86.2 mg/l of heavy labelled glutamic acid and cultures were incubated for 2 h or 6 h as noted. Samples were then prepared and extracted as detailed above.
Novel MRMs were developed via direct injection of authentic standards. Heavy glutamate (¹³C5,¹⁵N1) and heavy aspartate (¹³C4,¹⁵N1) were purchased from Cambridge Isotope Laboratories, Inc. Homologous signals were confirmed for heavy isotopes to maintain comparisons to normal isotope metabolites. All metabolites were detected in negative mode according to the same LC-MS/MS parameters detains above for negative mode metabolomics profiling. Aspartate was detected via 132- > 88 and 137- > 92 for normal and heavy isotope respectively. Glutamate was detected via 146- > 102 and 152- > 107 for normal and heavy isotope respectively. Glucose was detected via 179- > 89 and 185- > 92 for normal and heavy isotope respectively.
IPTG detection assay
Samples of 20 μl pooled sera (1 wk after starting IPTG-supplementation), 5 fed larvae, 10 unfed nymphs, or 3 fed nymphs were prepared and extracted as detailed above. IPTG standard was purchased from Gold Bio and detected in negative mode according to the LC-MS/MS parameters described above for negative mode metabolomics profiling. IPTG was detected with dual MRMs (237- > 161, 237- > 59) and signal retention times were compared to standard injections. Limit of quantitation (LOQ) was set at 3x baseline detection.
(p)ppGpp detection and thin layer chromatography
B. burgdorferi cultures were grown at 35 °C in BSK + RS to late log phase, washed three times in cold dPBS and resuspended in BSK + RS at 5 × 10^6^ Bb/ml containing 20 μCi/ml ^32^P either with or without IPTG as described and grown at 35 °C. At the times indicated, cells were collected by centrifugation at 6000 x g for 15 m at 4 °C, the supernatant discarded, and the cell pellets weighed. Nucleic acids were extracted by adding 20 μl of ice-cold 6.4 M formic acid and samples stored at -80 °C. Load volumes were standardized by pellet weight before separation on polyethylenimine (PEI) cellulose plates (EMD) by TLC as previously described^9^.
Statistics and reproducibility
All statistical analyses were performed using Prism software (v9.5.1; GraphPad Software, Inc.). All data is represented as mean ± SEM. Statistical analysis of growth curves and flow cytometry data was performed using two-way ANOVA. p values for cell length, qPCR DNA burdens, viability plating, IPTG detection and heavy amino acid labeling were determined for pairwise comparisons using two-tailed unpaired t tests. Exact p-values are provided when available, p-value < 0.05 was considered statistically significant.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Supplementary information
Transparent Peer Review file Supplementary Information Description of Additional Supplementary Files Supplementary data 1 Supplementary data 2 Supplementary data 3 Supplementary data 4 Reporting Summary
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Boyle, W. K. et al. Dks A Controls the response of the Lyme disease spirochete Borrelia burgdorferi to starvation. J. Bacteriol.20110.1128/JB.00582-18.(2019).10.1128/JB.00582-18PMC 635174430478087 · doi ↗ · pubmed ↗
- 2Groshong, A. M., Dey, A., Bezsonova, I., Caimano, M. J. & Radolf, J. D. Peptide uptake is essential for Borrelia burgdorferi viability and involves structural and regulatory complexity of its oligopeptide transporter. m Bio 810.1128/m Bio.02047-17.(2017).10.1128/m Bio.02047-17PMC 573691429259089 · doi ↗ · pubmed ↗
- 3Sze, C. W. et al. Lactate dehydrogenase is the Achilles’ heel of Lyme disease bacterium Borreliella burgdorferi. bio Rxiv 10.1101/2025.02.07.637162. (2025).10.1128/mbio.03728-24PMC 1198037640111021 · doi ↗ · pubmed ↗
- 4Boujnane, M. B., Boukerb, A. M., Connil, N. Bacterial gene expression in response to catecholamine stress hormones. Curr. Opin. Endocr. Metab. Res.36 (2024).
- 5Letunic, I. & Bork, P. Interactive Tree of Life (i TOL) v 6: recent updates to the phylogenetic tree display and annotation tool. Nucleic Acids Res.10.1093/nar/gkae 268. (2024).10.1093/nar/gkae 268PMC 1122383838613393 · doi ↗ · pubmed ↗