Road traffic noise exposure and blood DNA methylation at birth and in childhood: An epigenome-wide meta-analysis
Zhebin Yu, Irene Fontes Marques, Simon Kebede Merid, Kimberley Burrows, Ana Goncalves Soares, Andrei Pyko, Mikael Ögren, Göran Pershagen, Johanna Lepeule, Norun Hjertager Krog, Gunn Marit Aasvang, Michelle S.W. Kusters, Maria Foraster, Mariona Bustamante, Miriam Leskien

TL;DR
This study explores how road traffic noise might affect DNA methylation in children, suggesting possible biological mechanisms linking noise exposure to health effects.
Contribution
The study provides new evidence linking road traffic noise exposure to DNA methylation changes in children's blood.
Findings
46 DNA methylation sites showed suggestive associations with road traffic noise exposure.
One DNA methylation site (cg09400092) was significantly associated with recent noise exposure in an independent cohort.
93 differentially methylated regions were identified, with 14 showing nominal significance in an external validation cohort.
Abstract
Road traffic noise exposure has been associated with multiple adverse outcomes in epidemiological studies. However, the underlying biological mechanisms remain unclear. The aim of this study was to investigate the association between road traffic noise exposure and cord blood and child blood DNA methylation (DNAm). Data from six European studies (BAMSE, Generation R, HELIX, INMA, LISA, PIAMA) were used to perform the discovery epigenome-wide meta-analysis. Prenatal, infancy, and recent road traffic noise exposure was assessed at the residential addresses. Blood DNAm was measured using the Illumina 450 K or EPIC arrays. To identify differentially methylated positions (DMPs), we fitted robust linear regression models for each cohort, and the results were subsequently meta-analyzed. Differentially methylated regions (DMRs) were identified using Comb-p and DMRcate. Findings were then…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1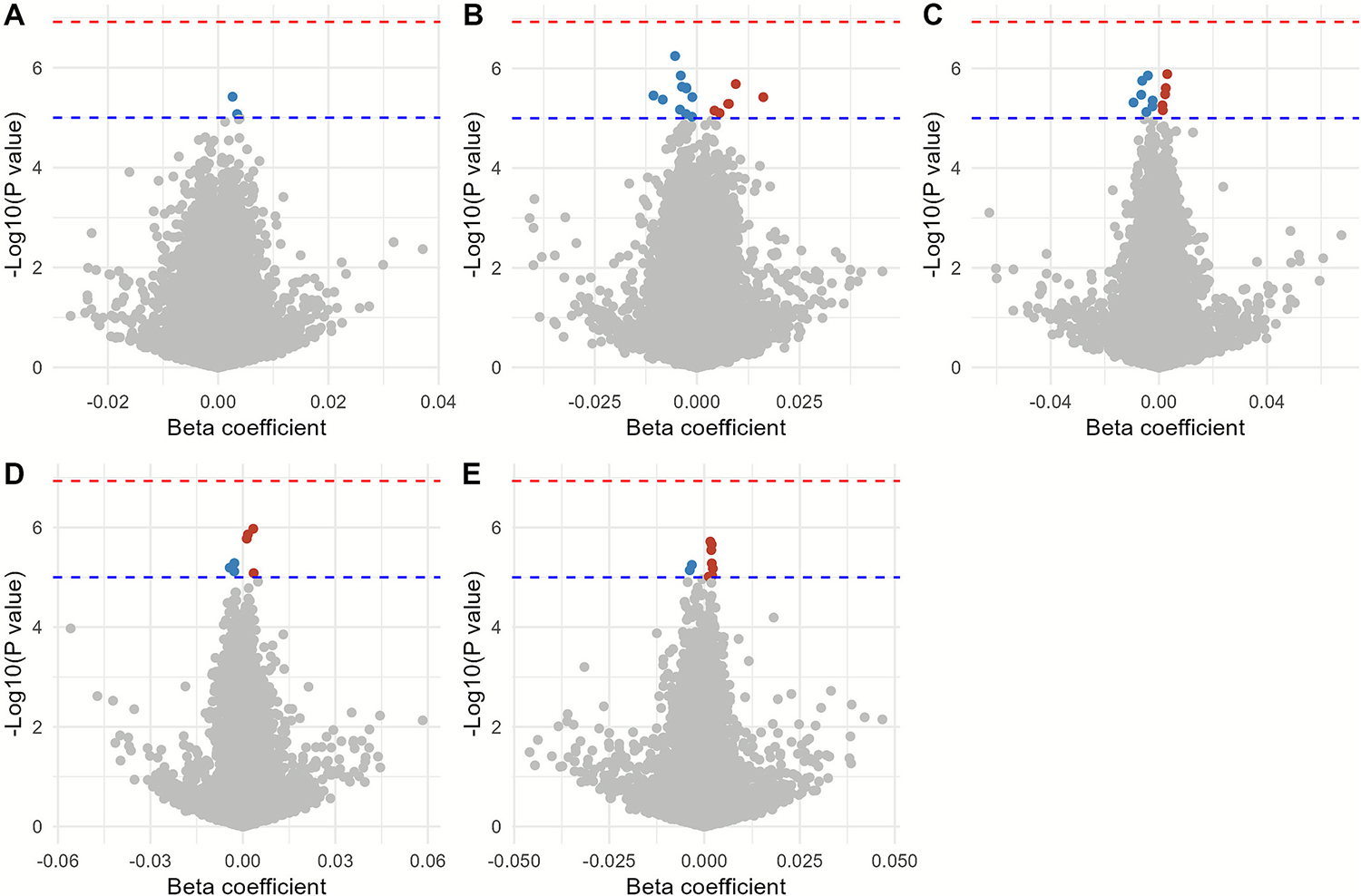 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNoise Effects and Management · Air Quality and Health Impacts · Biomarkers in Disease Mechanisms
Introduction
Road traffic noise is the second leading environmental stressor in Europe, according to the World Health Organization (WHO) (Basner and McGuire, 2018). A recent assessment of exposure to transportation noise showed that around 20 % of the European population is living in areas with harmful levels of noise (exceeding 55 dB in Day-Evening-Night Level (L_den_)) (Peris, 2020), and the population’s exposure to transportation noise is projected to increase due to urban expansion and growing demand for mobility (Peris, 2020). Numerous epidemiological analyses have demonstrated associations between noise exposure and adverse health effects, best documented for cardiovascular outcomes (Münzel et al., 2024), but also metabolic (Vienneau et al., 2024; Persson et al., 2024; Eze et al., 2017), respiratory (Liu et al., 2021; Eze et al., 2018), and reproductive outcomes (Sørensen et al., 2024) among adults, as well as hearing function (Selander et al., 2016), cognitive, behavioral, learning, and other neurodevelopmental outcomes among children and adolescents (Roche et al., 2024; Terzakis et al., 2022). Children may be more susceptible to the effects of noise due to the developing auditory system and because the impact of noise on hearing and quality of life at early stages of development can influence child health trajectories (Balk et al., 2023).
The proposed pathophysiological mechanisms underlying the adverse effect of noise exposure relate to the activation of stress response pathways (Münzel et al., 2017; Kuntic et al., 2023) and nighttime sleep disturbance (Kröller-Schön et al., 2018) (with dysregulation of the circadian clock), supported by animal studies (Münzel et al., 2021) and gene-environment studies (Eze et al., 2017). Noise can activate downstream stress response, such as the activation of the sympathetic nervous system (SNS) and the hypothalamic–pituitary–adrenal (HPA) axis, which further converges in oxidative stress and inflammation associated with endothelial Nitric Oxide Synthase (eNOS) uncoupling, endothelial dysfunction, high blood pressure and hyperglycemia, subsequently triggering adverse health outcomes (Sørensen et al., 2024).
DNA methylation (DNAm), a process during which methyl groups are added to the C5 position of the cytosine within a cytosine-guanine (CpG) dinucleotide, is the most widely studied epigenetic mark and may be a potential mechanism through which noise exposure may impact health outcomes (Leso et al., 2020). Indeed, noise exposure has been reported to be associated with differential DNAm at specific genes in rat brain tissue (Guo et al., 2017). Yet evidence of associations between road traffic noise exposure and DNAm in humans is scarce. The Swiss SAPALDIA cohort (Study on Air Pollution And Lung Disease In Adults) reported that long-term exposure to source-specific (road traffic, aircraft, railway) noise in adults was associated independently of exposure to traffic-related air pollution with differential blood DNAm of genes annotated to pathways related to inflammation, cellular development, and immune response (Eze et al., 2020). In a subset of the Isle of Wight birth cohort (Commodore et al., 2019), self-reported frequencies of heavy vehicles driving by the residential address was found to be associated with blood DNAm at 34 CpG sites at age 18 years. Another study, conducted among 610 female participants in Sweden (318 breast cancer cases and 292 controls), showed that road traffic noise exposure was associated with blood DNAm in two core circadian genes (Thacher et al., 2024). To the best of our knowledge, no epigenome-wide association study has been conducted in relation to noise exposure among children.
In the current study, we aimed to investigate the association between prenatal road traffic noise exposure and cord blood DNAm, as well as between infancy and recent road traffic noise exposure and child blood DNAm in a multi-cohort epigenome-wide association meta-analysis.
Methods
Study population
2.1.
A total of six European birth cohorts in the Pregnancy And Childhood Epigenetics Consortium (PACE) (Felix et al., 2018) were included in the discovery meta-analysis: BAMSE (Children, Allergy, Environment, Stockholm, Epidemiology), the Generation R Study, HELIX (Human Early Life Exposome), INMA (Environment and Childhood), LISA (Influence of Life-style factors on Development of the Immune System and Allergies in East and West Germany) (Heinrich et al., 2002) and PIAMA (Prevention and Incidence of Asthma and Mite Allergy). BAMSE comprises two different DNAm datasets from two separate projects, i.e., Mechanisms of the Development of Allergy that was assessed for genome-wide DNAm together with the PIAMA study (MeDALL) and EpiGene (Xu et al., 2018), while HELIX comprises one DNAm dataset including data from six independent European birth cohorts: BiB (Born in Bradford), EDEN (Study of determinants of pre- and postnatal development), INMA, KANC (Kaunas Cohort), MoBa (The Norwegian Mother and Child Cohort Study), Rhea (Mother-Child Cohort in Crete).
Detailed information about each cohort including recruitment and eligibility is provided in the Supplemental methods. Ethical approval for each cohort was granted by local institutional review boards and informed consent was obtained.
Noise exposure assessment
2.2.
Long-term exposure to road traffic noise was estimated at participants’ residential addresses. Road traffic noise exposure levels are expressed in L_den_ based on penalties of noise exposure during evenings (by 5 dB) and nighttime (by 10 dB). Noise levels were modeled using standardized approaches, incorporating traffic flow, road characteristics, land use, and building geometry. Detailed descriptions of noise exposure assessment at each cohort are provided in the Supplemental Methods. For the cord blood analyses, we calculated prenatal road traffic noise exposure as the average noise levels during pregnancy at the maternal residential addresses. For the child blood analyses, we calculated infancy road traffic noise exposure as the average noise levels from birth up to the child’s first birthday, and also the recent road traffic noise exposure (in early and late childhood) as the average noise levels for the 12 months before the biosampling date. Changing address during the exposure period was taken into account by using time-weighted averages across all residential histories.
DNA methylation profiling
2.3.
DNAm was measured in cord blood or child blood using the Illumina Infinium 450 K array in all included cohorts except LISA, where the EPIC array (version 1) was used. Sampling processing, quality control and normalization were handled by each cohort with details presented in the Supplemental Methods. Untransformed DNAm beta values were used as the outcome ranging from 0 to 1, with higher values indicating higher level of methylation at the CpG site.
Statistical analysis
2.4.
The overall design of the study is presented in Fig. 1.
Cohort-specific analysis
2.5.
In the discovery phase, six cohorts (BAMSE, Generation R, HELIX, INMA, LISA, PIAMA) conducted the EWAS analyses locally following a prespecified analysis plan and common statistical code. Associations between noise exposure and methylation levels across the epigenome were assessed using multiple robust linear regression analyses for each CpG site individually, implemented with the limma R package. Road traffic noise exposure was entered into the model as continuous variable in dB. Effect estimates are reported per 10 dB increase in noise levels. Analyses in cord blood were adjusted for an a priori selected panel of covariates: child sex, maternal education and cord blood cellular composition (Natural killer cells, B cells, CD4T and CD8T lymphocytes, Monocytes, Granulocytes, Nucleated red blood cells) (Gervin et al., 2019); while analyses in children were adjusted for child sex, maternal education, child age at biosampling, and blood cellular composition (Natural killer cells, B cells, CD4T and CD8T lymphocytes, Monocytes, Granulocytes) (Houseman et al., 2012). Information on cohort-specific data collection of the covariates as well as association between noise and cell types are presented in detail in Supplemental methods. Because several cohort-specific EWAS results showed potential inflation (lambda values > 1.2), we applied the Bacon (van Iterson et al., 2017) method to all cohort-specific EWAS. The lambda values for cohort-specific EWAS results before and after Bacon are presented in Supplemental Table S1. A sensitivity analysis was conducted by further adjusting maternal smoking during pregnancy in cord blood analysis, and for both maternal smoking during pregnancy and environmental tobacco exposure in child blood analyses. We also ran analyses further adjusting for ambient air pollution (particular matter ≤ 2.5 μm (PM_2.5_) and Black carbon) exposure modelled for the same exposure time-windows as noise exposure in five of the included cohorts (BAMSE Epigene,PIAMA, Generation R, LISA and HELIX). Details for PM_2.5_ and black carbon exposure assessment can be found elsewhere (Yu et al., 2024; Eeftens et al., 2012).
Meta-analysis for Differentially Methylated Positions (DMP)
2.6.
The results based on 450 K and EPIC arrays were meta-analyzed. The analysis was restricted to probes assessed in both 450 K and EPIC arrays, since only one cohort (LISA) measured DNAm using the EPIC array. We conducted five separate meta-analyses, i.e., for prenatal noise exposure and cord blood DNAm, infancy noise exposure and DNAm in early (4–6 years) and late (8–10 years) childhood, as well as recent noise exposure and DNAm in early and late childhood. Cohort-specific EWAS results after Bacon correction were meta-analyzed using fixed-effects inverse variance-weighting in METAL (Willer et al., 2010). All meta-analyses were independently repeated at Erasmus MC in Rotterdam and results were compared to minimize human error. We filtered out all cross-reactive probes defined by Chen et al (Chen et al., 2013), probes located on the sex chromosomes, and probes only available in one cohort (numbers of probes per model are presented in the Supplemental Methods). P-values were then False Discovery Rate (FDR) corrected for multiple comparisons using the Benjamini-Hochberg procedure (Benjamini and Hochberg, 1995). Genome-wide significance was defined as FDR P-value < 0.05 and suggestive significance as an absolute P-value < 1 × 10^–5^. We calculated the I^2^ statistic to explore heterogeneity across cohorts (Higgins and Thompson, 2002). Leave-one-study-out analyses were also conducted to explore if any of the individual studies were unduly influencing the findings.
Differentially Methylated Region (DMR) analysis
2.7.
Differentially methylated regions were identified using the Comb-p (Pedersen et al., 2012) and DMRcate (Peters et al., 2015) methods. Comb-p identifies DMRs by aggregating low p-values from neighboring CpG sites within a specific region, while DMRcate detects DMRs using a tunable kernel smoothing approach applied to association signals. Input for both DMR methods were the discovery meta-analyzed EWAS results including regression coefficients, standard errors, uncorrected P-values and chromosome positions (for Comb-p only). Detailed input parameters for DMR analysis are presented in Supplemental methods. We defined DMRs as those identified by both methods following multiple-testing correction (Sidak p-value < 0.05 for Comb-p and FDR-adjusted p-value < 0.05 for DMRcate), requiring at least two significant consecutive CpGs within the DMR. The DMRs were further annotated using the rGREAT method (Gu and Hübschmann, 2023).
Look-up in the ALSPAC study and SAPALDIA study
2.8.
We performed a look-up of the significant and suggestive findings in 589 newborns and 605 7-year-old children from the ALSPAC (Avon Longitudinal Study of Parents and Children) Study (see Supplemental methods for further information). This approach was chosen because the available data in ALSPAC did not allow road traffic noise exposure to be estimated as a continuous variable. Instead, it was estimated as a categorical exposure (L_den_ < 55 dB, 55–59.9 dB, ≥60 dB) and was entered into the model as an ordinal variable (Gonçalves Soares et al., 2024). Other than that, the EWAS analysis followed the same statistical code as in the discovery analysis. We looked up the suggestive DMPs from the discovery EWAS meta-analysis in the ALSPAC results. DMPs with p-values less than 0.05 divided by the number of tests as well as a consistent direction of association were considered significant. DMR analyses (Comb-p and DMRcate) were also conducted based on the ALSPAC EWAS results. We examined overlap between the DMRs from the meta-analysis and those in ALSPAC. We considered any DMRs that were FDR significant and that overlapped in terms of chromosome position as significant. Additionally, we report overlapping DMRs at the nominally significant level (absolute p-value < 0.05) with at least one nominally significant CpG (Broséus et al., 2024). We further performed a look-up of our top DMPs and DMRs using publicly available EWAS summary statistics from the SAPALDIA adult cohort, which examined transportation noise exposure in relation to DNAm using linear mixed-effect models (Eze et al., 2020).
Bioinformatics analyses
2.9.
Both suggestive DMPs as well as all the CpGs within the DMRs were used as input in the follow-up bioinformatics analyses. To test whether methylation levels of CpGs were associated with nearby gene expression levels, we looked them up in two publicly available resources: one dataset using 38 cord blood samples (Rojas et al., 2015; Barrett et al., 2013; Rager et al., 2014) and the HELIX Expression Quantitative Trait Methylation (eQTM) catalogue of children’s blood (Ruiz-Arenas et al., 2022). We also conducted enrichment analysis of the CpGs for Gene Ontology (GO) terms and pathways of the Kyoto Encyclopedia of Genes and Genomes (KEGG) and Reactome using the Enrichr website (Kuleshov et al., 2016). Moreover, we searched whether these CpGs have been previously associated with any exposure or health traits using the EWAS catalog (www.ewascatalog.org) and the EWAS Atlas (https://ngdc.cncb.ac.cn/ewas/atlas/index). We also looked up whether any CpGs have potential causal relationships with any disease using the DMRdb database, a disease-centric Mendelian randomization database (Zheng et al., 2024). Finally, eFORGE version 2.0 was used to test for enrichment of tissue-specific DNaseI hypersensitivity regions (Breeze et al., 2019).
Results
We meta-analyzed results from two cohorts with data on DNAm in cord blood (Generation R and INMA, N = 1477), and five cohorts with data on DNAm in child blood (N = 1129 for early childhood with data from BAMSE, Generation R, INMA, LISA, PIAMA; N = 2065 for late childhood with data from BAMSE, Generation R, HELIX, LISA, PIAMA). The description of demographic and lifestyle characteristics of the included participants as well as road traffic noise exposure levels are presented in Table 1 and in Supplemental Table S2. Road traffic noise exposure varied between cohorts: INMA had the highest level of road traffic noise (61.7 ± 6.1 dB in the prenatal exposure window) and cohorts from Netherlands (PIAMA, Generation R) had the lowest level (53.5 ± 4.6 and 54.5 ± 7.9, respectively, in the infancy exposure window). The quantile–quantile plots of meta-analyses did not reveal significant inflation in the distribution of observed p-values after BACON correction (lambda values ranged from 0.98 to 1.17, Supplemental Fig. S1).
We did not observe genome-wide significant DMPs (FDR < 0.05) in any of the considered exposure time windows. However, 46 DMPs reached suggestive significance (p-values < 1 × 10^–5^) across different exposure periods: two DMPs for prenatal road traffic noise exposure and DNAm in cord blood, 16 and 7 DMPs for infancy road traffic noise exposure and DNAm in early and late childhood, respectively, and 12 and 9 DMPs for recent road traffic noise exposure and DNAm in early and late childhood, respectively (Table 2, Fig. 2). The I^2^ values ranged from 0 to a maximum of 69.6, with 15 CpGs having a value of > 50 (Supplemental Fig. S2). Sensitivity analyses additionally adjusting for maternal smoking during pregnancy and second-hand smoke exposure in childhood did not materially change the coefficients for these DMPs (Supplemental Fig. S3). Leave-one-study-out analyses showed no undue influence from any single cohort (Supplemental Fig. S4), and additional adjustment for ambient PM_2·5_ or black carbon yielded similar estimates with Kolmogorov–Smirnov tests showing no significant differences in the distribution of beta coefficients and p-values (Supplemental Fig. S5). Look up for these DMPs in the EWASs of other exposure time windows suggested stronger consistency of association estimates within the same age groups (Supplemental Fig. S6).
We identified a total of 93 FDR-significant DMRs (88 unique) associated with road traffic noise exposure (overlapping between the Comb-p and DMRcate methods), with 9 for prenatal exposure, 26 and 18 for infancy noise exposure and DNAm in early and late childhood, as well as 12 and 28 for recent noise exposure and DNAm in early and late childhood, respectively). Among these DMRs, 5 were found in two different exposure time windows: chr13:47472050–47472429 annotated to HTR2A (infancy exposure with child blood age 4–6 and age 8–10), chr14:95826570–95826997 annotated to CLMN (infancy and recent exposure with child blood age 4–6), chr16:787799–788184 annotated to NARFL (prenatal exposure with cord blood and infancy exposure with child blood age 4–6), chr4:1041044–1041062 annotated to FGFRL1 (infancy and recent exposure with child blood age 4–6) and chr6:28583971–28584289 annotated to SCAND3 (infancy and recent exposure with child blood age 4–6) (Table 3 and Supplemental Table S3).
In the ALSPAC Study, one suggestive CpG in child blood age 8–10 (cg09400092 annotated to SSTR1) was significantly associated with recent road traffic noise exposure (same direction of association with p-value = 0.00165). None of the DMRs identified in the discovery meta-analysis were associated at the FDR level, but 14 DMRs with at least one overlapping CpG were found to be nominally significant (Table 3 and Supplemental Table S3). In the further look up analysis in the adult cohort (SAPALDIA), we did not replicate the suggestive CpGs. Among the 93 FDR significant DMRs that we identified in children, we found two childhood DMRs were also associated in the SALPADIA study (chr7:27142427–27143586 annotated to HOXA2 and chr13:47472050–47472429 annotated to HTR2A, nominal p-values = 0.013 and 0.024, respectively).
We did not find functional enrichment for the suggestive DMPs and CpGs within the DMRs for GO terms, KEGG or Reactome pathways after FDR correction (Supplemental Table S4–S8). We observed four significant eQTMs for the suggestive DMPs and 135 significant eQTMs for the CpGs within the identified DMRs (Supplemental Table S9). A total of 72 out of these 139 CpG-transcript associations were inverse and 67 were positive. The most statistically significant eQTM was cg26855724 with CRYZ expression (log2 fold-change = −0.32;(standard error = 0.25; P value = 3.96E- (Chen et al., 2013). According to the EWAS catalog and EWAS Atlas, the suggestive DMPs and CpGs within the DMR regions have been previously reported in relation to sex, child age, gestational age, pregnancy factors (maternal body mass index, plasma folate, pre-eclampsia), rheumatoid arthritis, mental disorders (attention-deficit/hyperactivity disorder, schizophrenia) and environmental exposures such as ambient air pollution and smoking (Supplemental Table S10). For the Mendelian Randomization analysis (look up in the DMRdb database), we found causal relationships between CpGs (cg00880741 and cg00955808) and multiple diseases such as asthma, metabolic syndrome and sleep disorders (Supplemental Table S11). Finally, the CpGs within the DMRs were found to be enriched in multiple tissues including blood, fetal muscle, lung, and pancreas (Supplemental Figs. S7–S8).
Discussion
In this epigenome-wide meta-analysis, we combined data from six population-based European cohorts to investigate the association between prenatal road traffic noise exposure and DNA methylation in cord blood, as well as infancy and recent road traffic noise exposure and DNA methylation in early and late childhood. Although no differentially methylated positions (DMPs) reached genome-wide significance at any time point, we identified 46 suggestively associated CpG sites, of which one (cg09400092, annotated to SSTR1) was also significant in an independent birth cohort (ALSPAC). In addition, we also identified 93 DMRs (88 unique) associated with road traffic noise exposure, of which 14 were also associated at nominal significance in the ALSPAC Study. We did not find significantly enriched biological pathways, but multiple CpGs were associated with gene expression and many were associated with various environmental and lifestyle exposures or health outcomes. Although the identified CpGs may not represent a biological mechanism linking noise exposure to health, they remain of interest as potential markers of road traffic noise exposure.
To our knowledge, this is the first epigenome-wide association study for road traffic noise exposure conducted among children. Differential methylation at one CpG, cg09400092, was associated with road traffic noise exposure in both the meta-analysis and the ALSPAC Study. SSTR1 (Somatostatin Receptor 1) plays a role in several important biological functions including cell growth regulation, hormone regulation and neurotransmission. This gene has been previously reported to be associated with obesity and non-alcoholic fatty liver disease in mice (Huang et al., 2024), a finding in line with the epidemiological associations observed between transportation noise exposure and overweight/obesity (Persson et al., 2024) and diabetes (Eze et al., 2017).
Despite the differences in study design, population, and statistical methods with the previous SAPALDIA study, we observed two overlapping DMRs, annotated to HOXA2 and HTR2A, between the SAPALDIA and the current discovery analysis. The HOXA2 (homeobox A2) gene encodes a transcription factor that is crucial for embryonic development and is essential for the proper development of facial structures and the middle ear. Mutations in this gene have been linked to microtia and hearing impairment (Brown et al., 2013). This finding may contribute to a better understanding of the mechanisms behind hearing impairment in children following exposure to noise as reported in earlier epidemiological studies (Selander et al., 2016; Balk et al., 2023). The HTR2A gene encodes the 5-HT2A receptor, which is a critical component of the serotonergic system in the human brain. One study conducted among 532 Chinese college students (mean age 24.3 years) showed that polymorphisms within the HTR2A gene were associated with individual differences in empathic and autistic-like traits (Gong et al., 2015). In a study in mice, stress exposure was associated with expression levels of the HTR2A gene (Maple et al., 2015), which aligns with the previous evidence that noise exposure may trigger cortical activation and release of stress hormones (Babisch, 2003), and over time chronic stress may increase risk of cardiovascular and metabolic diseases. Although findings from animal models may not directly translate to humans, they may provide mechanistic insights into how environmental stressors such as road traffic noise exposure could influence gene regulation and may contribute to long-term health effects.
Among the five DMRs that overlapped between exposure windows in the discovery meta-analysis, chr16:787799–788184 (annotated to NARFL) and chr13:47472050–47472429 (annotated to HTR2A) showed significant differential DNAm at different time points: prenatal exposure with cord blood and infancy exposure with child blood age 4–6 for NARFL, and infancy exposure with early and late child blood for HTR2A, respectively. This finding may indicate that early-life road traffic noise exposure may have persistent effects on these epigenetic patterns from cord blood to child blood, although the current study cannot employ a true longitudinal design due to the limited overlapping samples with DNAm data across ages from the same cohorts. The NARFL gene plays a crucial role in cellular defense mechanisms against oxidative stress. For instance, studies showed that deletion or knockdown of this gene in mice can lead to increased levels of reactive oxygen species (ROS), which is in line with the evidence of inflammatory and oxidative downstream effects of noise exposure (Münzel et al., 2017; Daiber et al., 2019).
The strengths of the current study are the relatively large sample size, the objective assessment of road traffic noise exposure, which was based on individual address levels using refined validated models, the availability of epigenome-wide DNAm data in cord blood and child blood, which allowed to investigate the associations of road traffic noise exposure at different exposure time windows, and a harmonized analysis plan.
Our study also has some limitations. Although a p-value threshold of 10^–5^ is commonly used as a suggestive cutoff in EWAS, it is more lenient than conventional FDR-based criteria and may therefore also capture some CpGs that are not truly associated with noise exposure. Therefore, these findings should be considered exploratory rather than genome-wide statistically significant and interpreted with appropriate caution. The included studies generally neither have information on some of the factors affecting noise exposure, such as house characteristics (residential floor, insulation and orientation of the rooms), habits of keeping windows open, nor estimated noise levels on the least exposed facade, which may affect the precision of modelled noise exposure levels and attenuate estimated associations. Noise is a complex exposure with multiple dimensions—such as intensity, duration, frequency spectrum, and individual perception — all of which may influence biological responses through distinct pathways. We used annual average noise exposure (L_den_) to facilitate comparison between pregnancy, 4–6 years, 8–10 years, and potentially adulthood. Although this noise metric is found to be associated with several health outcomes, including noise annoyance, in previous studies (Environmental noise guidelines for the European Region: executive summary, 2025), it does not capture all subjective responses to noise, nor the detailed acoustic characteristics that may drive downstream biological effects. Future studies are warranted to investigate additional aspects of noise exposure, e.g. different lag time, shorter/longer time window, frequency spectrum, noise types and sources, individual perception, exposure reducing behavior, etc. Due to the observational study design, we cannot conclude anything on potential causal relationships. Although we have adjusted for predefined covariates in the main analysis as well as tested the role of maternal smoking and environmental tobacco smoking in the sensitivity analysis, residual confounding e.g, by maternal factors (i.e. maternal stress, maternal nutrition status) cannot be ruled out. Future studies with more comprehensive data on these characteristics are needed to better disentangle the complex relationships. Although low to moderate correlations between road traffic and air pollution were observed (Supplemental Table S12) and additionally adjusted for traffic related air pollution in four study did not materially change the associations of noise with DNAm, further work needs to be done to fully disentangle effects of road traffic noise and traffic related air pollution. Other area-level exposures, such as socioeconomic status (SES) and greenness measured as Normalized Difference Vegetation Index (NDVI) may be also associated with road traffic noise exposure as well as DNAm, and may therefore act as confounder. However, in we examined these correlations in one of the included studies (BAMSE EpiGene) and found very weak correlations between noise exposure and area-level SES (r = 0.02) or NDVI (r = −0.01). Further adjusting for these variables yielded effect estimates that were highly correlated with those from the main model (r = 0.88 for SES-adjusted and r = 0.87 for NDVI-adjusted models), suggesting that these area-level factors are unlikely to have materially confounded our findings. The current study was restricted to the European population due to data availability, which limits the generalization of our findings to other populations. The between-cohort differences in geographical settings, study period, methods for noise exposure assessment, as well as quality control, normalization, and adjustment for technical variation in the DNAm data could to some extent contribute to diluting possible associations, although an earlier published EWAS meta-analysis including the same cohorts reported robust results in relation to different data processing methods used across the cohorts for normalization and corrections for technical variables (Joubert et al., 2016). In addition, DNAm signatures are known to be cell- and tissue specific. Like many other EWAS analyses we used cord and peripheral blood cells to investigate the association between environmental exposure and DNAm patterns, which may not be the most directly relevant tissue in the pathway to noise-related health effects. The results from the eFORGE analysis showed enrichment in specific cells and tissues, indicating that future research using other relevant cells and tissues may be useful (Broséus et al., 2024; Mortillo and Marsit, 2023).
In conclusion, we observed suggestive evidence of the association between road traffic noise exposure and DNA methylation in cord and child blood, that overlapped in part with associations observed in adults. Further studies, ideally with larger sample sizes and more specific measures of noise exposure, are needed to provide additional insights into the role of DNA methylation as a marker or underlying mechanism linking road traffic noise exposure to adverse health outcomes.
Supplementary Material
Supplementary methods
Supplementary tables
Supplementary figures
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Basner M, Mc Guire S, 2018. WHO Environmental Noise guidelines for the European Region: a Systematic Review on Environmental Noise and Effects on sleep. Int. J. Environ. Res. Public Health 15.10.3390/ijerph 15030519 PMC 587706429538344 · doi ↗ · pubmed ↗
- 2Peris E, 2020. Environmental noise in Europe: 2020. Eur Environ Agency 1, 104.
- 3Münzel T, Molitor M, Kuntic M, , 2024. Transportation Noise Pollution and Cardiovascular Health. Circ. Res 134, 1113–1135.38662856 10.1161/CIRCRESAHA.123.323584 · doi ↗ · pubmed ↗
- 4Vienneau D, Wicki B, Flückiger B, Schäffer B, Wunderli JM, Röösli M, 2024. Long-term exposure to transportation noise and diabetes mellitus mortality: a national cohort study and updated meta-analysis. Environ. Health 23, 46.38702725 10.1186/s 12940-024-01084-0PMC 11068573 · doi ↗ · pubmed ↗
- 5PerssonÅ, Pyko A, Stucki L, , 2024. Long-term exposure to transportation noise and obesity: a pooled analysis of eleven Nordic cohorts. Environ. Epidemiol 8, e 319.38983882 10.1097/EE 9.0000000000000319 PMC 11233097 · doi ↗ · pubmed ↗
- 6Eze IC, Foraster M, Schaffner E, , 2017. Long-term exposure to transportation noise and air pollution in relation to incident diabetes in the SAPALDIA study. Int. J. Epidemiol 46, 1115–1125.28338949 10.1093/ije/dyx 020PMC 5837207 · doi ↗ · pubmed ↗
- 7Liu S, Lim Y-H, Pedersen M, , 2021. Long-term air pollution and road traffic noise exposure and COPD: the danish Nurse Cohort. Eur. Respir. J 58, 2004594.33986028 10.1183/13993003.04594-2020 · doi ↗ · pubmed ↗
- 8Eze IC, Foraster M, Schaffner E, , 2018. Transportation noise exposure, noise annoyance and respiratory health in adults: a repeated-measures study. Environ. Int 121, 741–750.30321849 10.1016/j.envint.2018.10.006 · doi ↗ · pubmed ↗